
Abstract
This study aimed to investigate the protective effects of erythrocyte-mediated endoplasmic reticulum (ER) stress in macrophages in hemorrhagic shock. An hemorrhagic shock model was established in male BALB/c mice. Animals were randomly divided into three groups (n = 8): control group (A), erythrocyte reinfusion group (B), and TLR9 inhibition group (C). Eight healthy BALB/c mice were also included as group N (n = 8). Mice in group A were not treated, while mice in groups B and C were transfused with red blood cells separated from the blood of mice in group N. Flow cytometry was used to detect the expression of erythrocyte surface protein TLR9 in each group. Immunofluorescence assay was used to analyze the distribution and relative expression of protein STING in macrophages. Flow cytometry was used to analyze the expression of STING, ATF6, and IRE1 in macrophages. Enzyme-linked immunosorbent assay was used to analyze the levels of inflammatory signal molecules, including IFN-α, IFN-β, IL-6, CCL4, CCL5, and IL-6. FITC-Annexin V was used to analyze the apoptosis of immune cells (macrophages) in mouse blood samples and to detect the concentration of calcium ions in erythrocyte cytoplasm. The results showed that the expression of erythrocyte surface protein TLR9; the distribution of STING-positive cells in macrophages; the expressions of STING, ATF6, and IRE1 in macrophages; the levels of inflammatory signal molecules; the apoptosis rate of macrophages; and the intracellular calcium concentration in erythrocytes in group B were higher than those in group A, followed by group C. These results suggest that TLR9 regulates ER stress in macrophages of mice with hemorrhagic shock through the TLR9-cGAS-STING-IFN signaling pathway. Increased expression of TLR9 enhanced macrophage activity, reduced apoptosis, enhanced inflammatory response and immune response, and restored electrolyte level, which might be a therapeutic option for the treatment of hemorrhagic shock.
Keywords
Introduction
Hemorrhagic shock (HS) is a common critical clinical condition that can increase the susceptibility to infection, leading to sepsis and multiple organ failure due to the inhibition of cellular immunity 1 . TLR9 is a surface protein receptor for mitochondrial DNA (mtDNA). cGAS is a sentry response protein in cells. It detects foreign and altered DNA and then initiates a cascade of signals triggering the internal defense function 2,3 . When erythrocytes carrying mtDNA reach macrophages via blood circulation, the endoplasmic reticulum (ER) of macrophages can be activated 4 . Protein interferon gene stimulator protein (STING), located in ER, is a downstream signal molecule of cGAS. Combining with TBK1, STING induces the production of I-IFNs (IFN-α and IFN-β). I-IFNs regulate inflammatory factors through the PI3K-AKT signaling pathway to protect immune cells and organs from apoptosis and damage 5,6 . The aim of this study was to investigate the protective effects of erythrocyte-mediated ER stress in macrophages in HS.
Materials and Methods
Laboratory Animals and Grouping
Forty healthy and clean BALB/c male mice (weighing 20–28 g) were purchased from the Laboratory Animal Research Institute of Chinese Academy of Medical Sciences (Beijing, China). Mice were acclimated to the environment and fasted for 12 h before experiments, during which they had free access to drinking water. All experimental procedures were performed according to the ethical standards of experimental animals. Eight healthy BALB/c mice were randomly selected as group N (n = 8). The rest animals were subjected to the establishment of HS model by cardiac puncture as previously described 7 . Twenty-seven mice were successfully modeled. Twenty-four of them were randomly selected and divided into three groups (n = 8): control group (A), erythrocyte reinfusion group (B), and TLR9 inhibition group (C).
Treatment
Fresh blood samples were obtained from the healthy mice in group N. The red blood cells were separated and transfused to mice in groups B and C. Mice in group C mice were concomitantly given TLR9 antibody via intravenous injection and their venous blood samples were collected after 3 days of treatment. Animals in group A were not treated.
Measurements
A volume of 5 ml fresh blood was obtained from mice in groups A, B, and C and isolated into single cell suspension. After centrifugation for 15 min at 1,500 r/min, the supernatant was discarded. Cells were resuspended with 0.35 ml BDTM stationary solution to obtain single cell suspension and stored at 4°C for further analysis. Flow cytometry was used to detect the expression of erythrocyte surface protein TLR9 in each group of mice. Cells were washed with phosphate buffered saline (PBS) and centrifuged at 1,500 r/min for 5 min. After two repetitions, samples were blocked with 2% rat serum and reacted with rabbit anti-TLR9 (primary antibody) 50 ml (1:50) for 1.5 h at 4°C. After two washes with PBS, the second antibody was added 50 ml (1:50) and incubated at 4°C for 45 min. After two washes with PBS, 30 µl PBS was added for detection. The fluorescence was detected by the Listmodl software system. The Multicycle software was used for data processing and the positive cell rate was recorded. Immunofluorescence analysis was performed to determine STING distribution and relative expression in macrophages. Macrophages in blood samples were obtained and cultured in RPMI-1640 containing 10% fetal bovine serum (FBS, Gibco, Invitrogen, Paisley, UK) in a 5% CO2 incubator at 37°C. After two days, adherent cells were collected. Cells were fixed at −20°C for 5 min with 4% formaldehyde, blocked with 10% rat serum at room temperature for 1 h, and then incubated with rabbit anti-STING-1 (1:100) overnight at 4°C. After being washed with PBS, samples were incubated with the second antibody (1:100) at room temperature for 45 min, and fluorescence microscopy was used to observe 60% buffered glycerol seals. Single cell suspension was analyzed by flow sorting. Cells (1 × 107) were suspended with 300 µl buffer (5% FBS + RPMI-1640) and incubated with 3 µl FC blocking antibody at room temperature for 5 min. Then cell suspension was divided into two tubes. Tube 1 was the negative control. Tube 2 was labeled with 3 µl CD11b-APC antibody and 3 µl CD11c-PE antibody for 20 min at 4°C. Then samples were washed twice with flow buffer and then resuspended with 2.5 ml flow buffer for flow cytometry. The protein expressions of STING, ATF6, and IRE1 in macrophages were analyzed by flow cytometry. A volume of 1.0 ml single cell suspension was taken and the rabbit anti-STING, ATF6, and IRE1 antibodies were used as the first antibody. The levels of inflammatory signaling molecules, including IFN-α, IFN-β, IL-6, CCL4, CCL5, and CXCL10, were analyzed by enzyme-linked immunosorbent assay (ELISA). The venous blood was extracted from mice and collected in an anticoagulant tube of ethylenediaminetetraacetic acid. The blood samples were centrifuged at room temperature for 0.5–1 h and stored at −20°C for ELISA analysis. The detection was performed according to the manufacturer’s instructions. The apoptosis of immune cells in mouse blood was analyzed by Flow FITC-Annexin V analysis. After stimulating for apoptosis, cells were collected in flow tube, centrifuged at 4°C for 10 min at 1,000 r/min, washed with PBS twice, and then resuspended. Suspended cells (5.0 × 104) were centrifuged for 5 min at 1,000 r/min to remove the supernatant. After the addition of 195 µl FITC-Annexin V conjugate, cells were suspended and added with 5 µl FITC-Annexin V. Then 10 µl propylene iodide staining solution was added. Flow cytometry was performed in triplicate. The intracytoplasmic calcium concentration in erythrocytes was detected using fluo-3 fluorescence labeling and flow cytometry. Venous blood samples (2 ml) were collected and centrifuged for 10 min at room temperature of 1,000 r/min. Then membrane and plasma were carefully separated. Samples were washed with normal saline for three times. The supernatant was discarded after centrifugation for 10 min at 1,000 r/min. Fluo-3 fluorescent indicator (10 μl) was added to the prepared erythrocyte suspension. The supernatant was incubated at 37°C for 30 min and centrifuged at 800 r/min for 10 min. After adding 2 ml pyrolysis solution, the concentration of Ca2+ on the side of fluorescence spectrophotometer was measured.
Statistical Method
SPSS19.0 software was used to analyze the data. Numerical data are expressed as mean ± standard deviation. T-test was used for the comparison between groups. Categorical data are expressed in percentage (%). Chi-square test was used for the comparison between groups. P < 0.05 was considered statistical significance.
Results
The Expression of Erythrocyte Surface Protein TLR9 in Each Group of Mice
The percentage of positive cells in groups A, B, and C was 10.5%, 14.4%, and 5.3%, respectively. Group B showed higher percentage of positive cells compared to group A. Group C has the lowest percentage of positive cells among the three groups and the difference was statistically significant (P < 0.05, Fig. 1).

Expression of erythrocyte surface protein TLR9 in three groups of mice.
The Distribution and Relative Expression of Protein STING in Macrophages
Immunofluorescence analysis showed that the protein expression of STING in group B was higher than that in group A. The STING expression in group C was lower than that in both group A and group B. The difference was statistically significant (P < 0.05, Fig. 2).
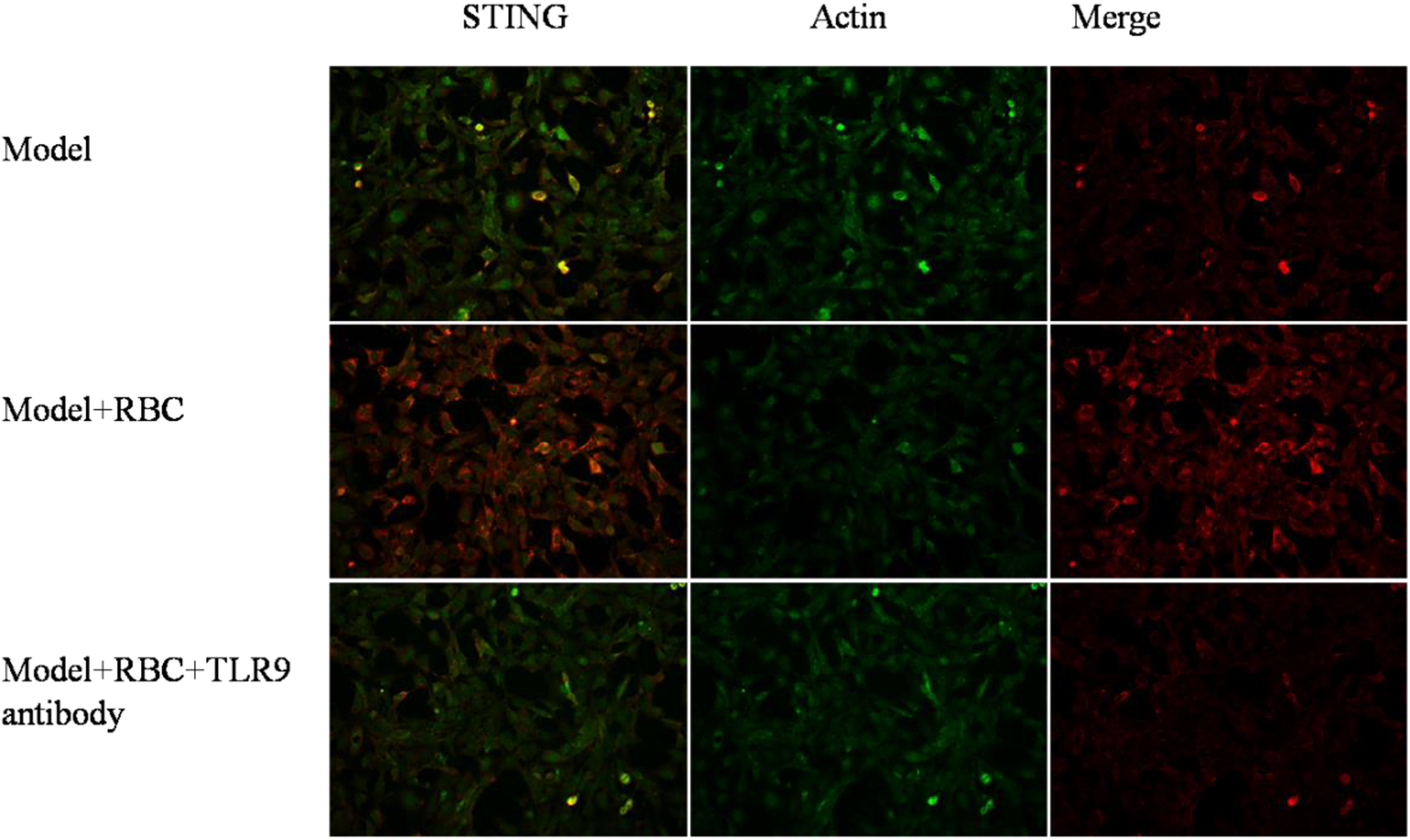
Distribution of protein STING in macrophages.
Flow Sorting of Macrophages in Each Group of Mice
The distribution of M1 and M2 in macrophages by flow cytometry showed that the ratio of M1 to M2 was 33.8% and 36.7%, respectively (Fig. 3).

Distribution of M1 and M2 in macrophages of mice.
The Protein Expressions of STING, ATF6, and IRE1 in Macrophages
The protein expressions of STING, ATF6, and IRE1 in macrophages in group B were higher than those in group A. The expressions of the above-mentioned proteins in group C were lower than those in group A and group B. The difference was statistically significant (P < 0.05, Fig. 4).
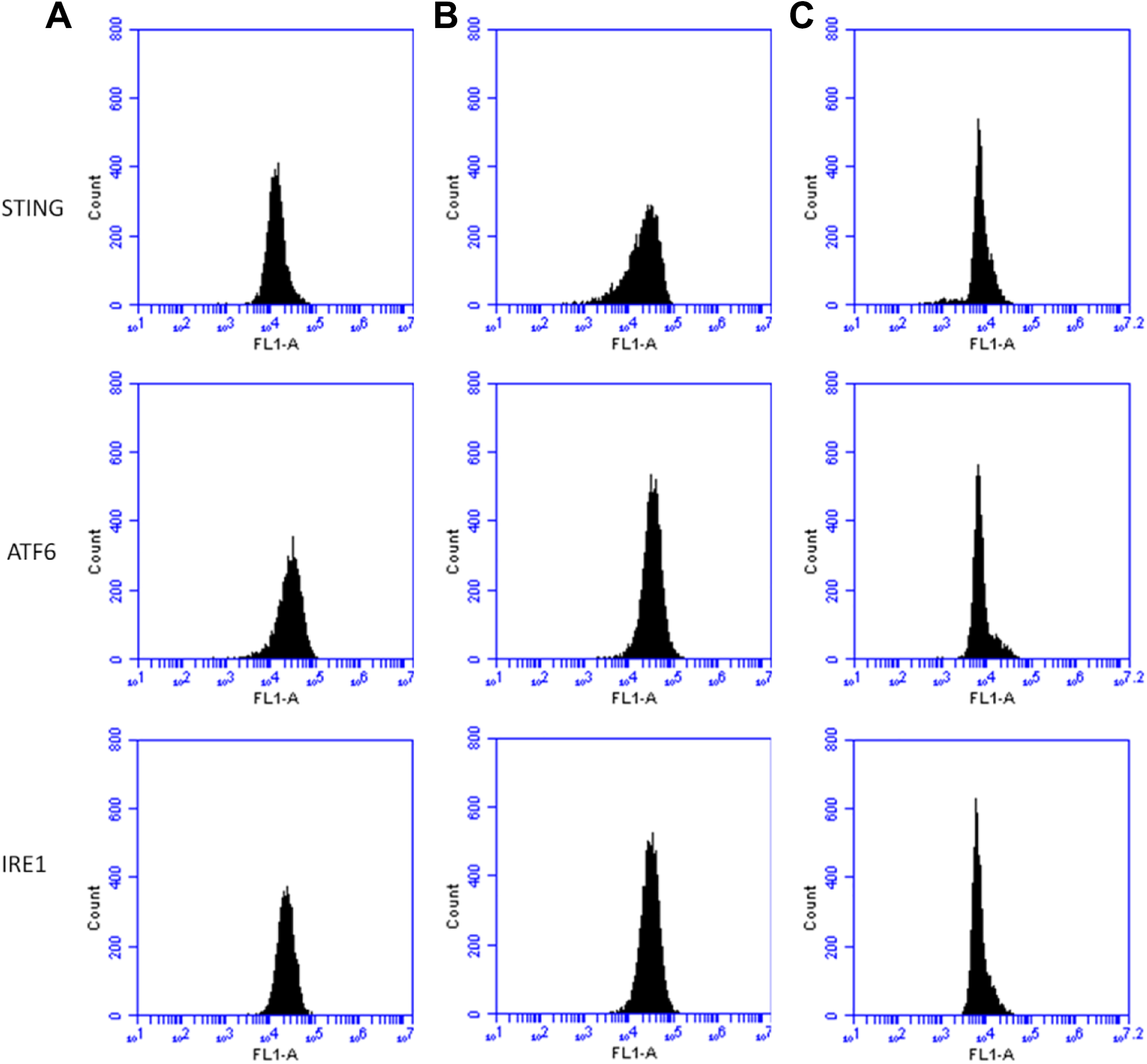
Expression of STING, ATF6, and IRE1 in macrophages.
The Secretion of Inflammatory Signaling Molecules
The levels of inflammatory signal molecules (IFN-α, IFN-β, IL-6, CCL4, CCL5, and CXCL10) shown in Fig. 5 were higher in group B than in group A. Group C showed the lowest levels of these molecules compared to groups A and group B with statistical significance (P < 0.05, Table 1).

Expression of inflammatory signaling molecules IFN-α, IFN-β, IL-6, CCL4, CCL5, and CXCL10.
Expression of Inflammatory Signaling Molecules IFN-α, IFN-β, IL-6, CCL4, CCL5, and CXCL10.

The Apoptosis of Immune Cells in Mouse Blood
The early apoptotic rates of immune cells in groups A, B, and C were 5.20%, 22.20%, and 2.10%, respectively. Group B showed a higher rate than group A, and group C had the lowest level of apoptotic cell death, which was statistically significant (P < 0.05, Fig. 6).

Apoptosis of immune cells in the blood of mice.
The Concentration of Intracytoplasmic Calcium in Erythrocytes
The intracytoplasmic calcium concentration in erythrocytes in groups A, B, and C was 170.33, 337.16, and 105.29 nM, respectively. The calcium concentration in group B was higher than group A, and group C showed the lowest concentration of intracytoplasmic calcium, which was statistically significant (P < 0.05, Fig. 7).

Calcium concentration in erythrocyte cytoplasm.
Discussion
HS can lead to the increase of macrophage apoptosis, the decrease of macrophage phagocytosis of apoptotic cells, the production of inflammatory reaction, and ultimately immune dysfunction 8 . TLR9 is a highly conserved transmembrane protein that belongs to the Toll-like receptor family. It shows inhibitory activity against cell growth and apoptosis. TLR9 is an important receptor for identifying invasive microorganisms, mediating innate immune cells, and recognizing mtDNA 9 . MtDNA is an extranuclear genetic material. When hypoxia and ischemia occur, mtDNA can cause cell energy metabolism disorder by regulating mitochondrial oxidative phosphatization 10 . STING is primarily expressed in mammals. As an important connective protein, STING can sense the invasion of microorganisms to host cells. It is mainly localized on the ER in resting state and plays an important role in immune response 11 . In this study, the expression of erythrocyte surface protein TLR9 was higher in group B than that in group A. The expression of TLR9 inhibitor in group C was lower than that in groups A and B. These findings suggested that the reinfusion of red blood cells increased the expression of TLR9. The results of the distribution and relative expression of STING in macrophages were consistent with those of TLR9, indicating that TLR9 might affect the expression of STING. In this study, transfused red blood cells carried mtDNA to macrophages, which were recognized by macrophage TLR9 and activated the STING signaling pathway.
Macrophages play an important role in cellular immunity. Antigen presenting and the release of immune active factors are the main function of macrophages in immune regulation. They can activate T cells and secrete various lymphokines, such as IL-1 and IL-6 12 . ATF6 and IRE1 are type II transmembrane proteins on the ER, which play an important role in ER stress response. ATF6 is normally situated on the ER surface and is inactive. ATF6 transports to the Golgi apparatus and finally to the nucleus during ER stress. It regulates the activation of transcription factors and various enzymes, and promotes cells to resume normal function 13 . This study showed that erythrocyte transfusion induced the expression of STING, ATF6, and IRE1 in macrophages, indicating that the activity of ER was increased. However, when TLR9 was inhibited, the expression of these proteins was downregulated compared to the control group and the erythrocyte transfusion group. The expression of STING, ATF6, and IRE1 in phagocytes was consistent with that in erythrocytes, suggesting that TLR9 may play a role in regulating the activity of ER in macrophages.
Apoptosis is a form of cell death. One of the important features of apoptosis is that the cell membrane remains intact during this process and is engulfed by macrophages before lysis 14 . Neutrophils are important inflammatory cells. The apoptosis of neutrophils during shock is a compensatory response of the body. If macrophages fail to clear neutrophils in time, they will release a large amount of toxic substances in cells and enhance inflammatory response 15 . In this study, we showed that the secretion of inflammatory signaling molecules IFN-α, IFN-β, IL-6, CCL4, CCL5, CXCL10, and erythrocyte reinfusion group were induced in group B, and the TLR9 inhibition group exhibited the lowest levels of these molecules. The apoptotic rate in all groups suggested that the transfusion of red blood cells reduced the apoptosis of macrophages. Macrophages are one of the main sources of IFN-α, IFN-gamma, and IL-6 in early immune response. The secretion of IFN-gamma will promote the secretion of INF-gamma by NK cells and Th cells and enhance the immune response of the body. IFN-gamma acts as feedback on macrophages and activates some macrophages into immune cells capable of phagocytizing pathogenic microorganisms 16,17 . CCL4, CCL5, and CXCL10 are chemokines that play an important chemotactic role in infectious immune diseases. They can induce the chemotaxis of a variety of inflammatory cells, such as monocytes, macrophages, and lymphocytes, until they reach the inflammation site 18 .
Calcium ion is an important substance to maintain the integrity of cell function and structure. It plays a coupling and second messenger role in cell activity, secretion, and metabolism 19 . Usually, intracellular and extracellular calcium ions keep a dynamic balance. In HS patients, the dynamic balance between intracellular and extracellular calcium ions was disrupted. Calcium ion is also closely related to cell apoptosis. Elevated concentration of calcium ion can inhibit cell apoptosis 20 . In this study, the concentration of calcium ion in erythrocyte cytoplasm increased after erythrocyte transfusion, indicating improved electrolyte balance in mice. The calcium ion concentration was decreased in TLR9 inhibition group, indicating aggravated electrolyte imbalance. These data implied that the imbalanced electrolyte level may be caused by the decrease of immune response and the aggravation of HS.
There are some limitations in this study. First, the sample size was relatively small. Further studies with a larger sample size are needed to validate the results obtained from the current study. Second, other factors related to ER stress (e.g., reactive oxygen species) and inflammatory responses (e.g., IL-4 and IL-10) may also be included in future studies. Third, a TLR9 overexpression model may be used to further demonstrate the effect of TLR9 upregulation in ER stress in HS.
In conclusion, TLR9 regulates ER stress in macrophages of HS mice through the TLR9-cGAS-STING-IFN signaling pathway. The upregulation of TLR9 increased macrophage activity, reduced apoptosis, enhance inflammatory response and immune response, restored electrolyte levels, suggesting that TLR9 upregulation might be a therapeutic option for the treatment of HS.
Footnotes
Acknowledgments
We would like to acknowledge the reviewers for their helpful comments on this article.
Author Contributions
Each author has made an important scientific contribution to the study and has assisted with the drafting or revising of the manuscript. All of the authors have consented to publish this research.
Ethical Approval
All animal experiments were approved by the Animal Research Ethics Committee of The Shanghai Gongli Hospital, the Second Military Medical University.
Statement of Human and Animal Rights
All of the experimental procedures involving animals were conducted in accordance with the Institutional Animal Care guidelines of Shanghai Gongli Hospital, the Second Military Medical University, Shanghai, China and approved by the Administration Committee of Experimental Animals, Shanghai, China.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work is supported by Key Disciplines Group Construction Project of Pudong Health Bureau of Shanghai (No. PWZxq2017-10) and National Natural Science Foundation of China (No. 81870147).