
Abstract
Preterm infants have a high risk of neonatal white matter injury (WMI) caused by hypoxia-ischemia. Cell-based therapies are promising strategies for neonatal WMI by providing trophic substances and replacing lost cells. Using a rat model of neonatal WMI in which oligodendrocyte progenitors (OPCs) are predominantly damaged, we investigated whether insulin-like growth factor 2 (IGF2) has trophic effects on OPCs in vitro and whether OPC transplantation has potential as a cell replacement therapy. Enhanced expression of Igf2 mRNA was first confirmed in the brain of P5 model rats by real-time polymerase chain reaction. Immunostaining for IGF2 and its receptor IGF2 R revealed that both proteins were co-expressed in OLIG2-positive and GFAP-positive cells in the corpus callosum (CC), indicating autocrine and paracrine effects of IGF2. To investigate the in vitro effect of IGF2 on OPCs, IGF2 (100 ng/ml) was added to the differentiation medium containing ciliary neurotrophic factor (10 ng/ml) and triiodothyronine (20 ng/ml), and IGF2 promoted the differentiation of OPCs into mature oligodendrocytes. We next transplanted rat-derived OPCs that express green fluorescent protein into the CC of neonatal WMI model rats without immunosuppression and investigated the survival of grafted cells for 8 weeks. Although many OPCs survived for at least 8 weeks, the number of mature oligodendrocytes was unexpectedly small in the CC of the model compared with that in the sham-operated control. These findings suggest that the mechanism in the brain that inhibits differentiation should be solved in cell replacement therapy for neonatal WMI as same as trophic support from IGF2.
Keywords
Introduction
Hypoxia-ischemia (H-I) in the perinatal period is an important cause of cerebral palsy in children. In the term infant, acute encephalopathy, also known as H-I encephalopathy 1,2 , is induced by compromised oxygen and glucose supply to the brain leading to cellular energy failure 3,4 . In the preterm infant, there is a high risk of neonatal white matter injury (WMI) because oligodendrocyte progenitor cells (OPCs), which in human are abundant at gestational weeks 20–28 5 , are particularly susceptible to H-I 6 –8 . With advances in neonatal intensive care, 9 –11 neonatal WMI has been reduced to a milder form, characterized by a higher prevalence of nondestructive lesions 12 –15 . Ischemia-induced neuroinflammation and the prenatal inflammatory response are also related to neonatal WMI 16,17 .
We previously established a rat model of neonatal WMI 18 in postnatal (P) day 3 (P3) animals by right common carotid artery occlusion followed by exposure to 6% oxygen (hypoxia) for 1 h. This model is based on the notion that late-phase OPCs in the immature brain are especially vulnerable to H-I 6 –8 . In this model, actively proliferating OPCs are damaged, resulting in a decreased number of mature oligodendrocytes (OLs) and hypomyelination in the adult sensorimotor cortex 19 . The model rats exhibit moderate motor deficits, especially in the hind limbs, accompanied by disorganization of OL development in layers II–III of the sensorimotor cortex 20 .
Cell-based therapy with grafted cells is a promising treatment strategy for neonatal WMI by providing neurotrophic substances and replacing lost OPCs 21 –23 . Many studies have shown that grafted cells provide trophic support, 24 –27 and we have reported the upregulation of several genes encoding trophic factors, such as insulin-like growth factor 2 (IGF2) in the ipsilateral sensory motor cortex of neonatal WMI 19 . We hypothesized that OPC transplantation into our neonatal WMI model could effectively replace lost late-phase OPCs as same as trophic support.
This study investigated whether the upregulation of IGF2 has an effect on cultured OPCs and whether OPC transplantation into our neonatal WMI model has potential as a cell replacement therapy. We show that enhanced IGF2 levels induce differentiation of OPCs in vitro and that OPCs grafted into the corpus callosum (CC) of nonimmunosuppressed neonatal WMI model rats showed long-term survival for at least 8 weeks after grafting and that the number of mature OLs was unexpectedly small in the CC compared with that of sham-operated control. These findings suggest that the mechanism to inhibit OLs differentiation in the CC of the neonatal WMI model should be solved, as same as by enhancing trophic support from IGF2, for better outcome in cell replacement therapy.
Materials and Methods
A Rat Model of Neonatal WMI
Animals were cared for according to the guidelines of the Institute for Experimental Animal Science, Nagoya City University Medical School, Japan. All experimental procedures were approved by the Committee of Animal Experimentation of Nagoya City University Medical School, and every effort was made to minimize the pain and discomfort of the animals used.
Male Wistar P2 or P3 rats and their mothers were obtained from Japan SLC, Inc. Animals were housed under a constant 12-h dark/light cycle and had access to standard bedding. The model was established according to a previously described method 18 . Briefly, P3 pups were subjected to right common carotid artery occlusion (RCAO) under isoflurane (Pfizer, NY, USA) anesthesia (induction at 5% followed by maintenance at 1.3%). After a 2-h recovery period with their dam, pups were exposed to 6% (v/v) hypoxia for 60 min. The pups were then returned to their dam and maintained under normal conditions. Sham-operated rats without hypoxia were used as a control.
Real-Time Polymerase Chain Reaction (PCR) for IGF2
To confirm the expression of Igf2 in neonatal WMI and sham-operated control rats at P5, real-time PCR was performed as reported 28 . Briefly, 3 mm thick coronal brain sections containing the sensorimotor cortex (bregma: +1.4–1.6 mm, lateral: 1.0–3.2 mm) were obtained using a brain slicer (ASI instrument, Eugene, OR, USA), and total RNA was isolated using Trizol reagent (Invitrogen, Carlsbad, CA, USA). PCR was performed by incubation at 95°C for 10 min, followed by 40 cycles of 15 s at 95°C, 1 min at 60°C, 45 s at 72°C, and 15 s at 80°C (for SYBR Green detection) using an ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster, CA, USA). Gapdh mRNA was used as an internal control, and gene expression was normalized against a corresponding amount of Gapdh mRNA. The relative amounts of each product were calculated by the comparative CT(2-ΔΔCT) method. Amplification was performed with the following primers: IGF2: 5′-TGTCTACCTCTCAGGCCGTACTT-3′ and 5′-CAGGTGTCGAATTTGAAGAACTTG-3′; GAPDH: 5′-TGTGTCCGTCGTGGATCTGA-3′ and 5′-CCTGCTTCACCACCTTCTTGA-3′.
For comparison of Igf2 expression between ipsilateral sensorimotor cortex, contralateral sensorimotor cortex, and sham-operated control cortex, the cortical samples of H-I received rat and sham-operated control rat were used. Data for each sample are represented as multiples of the sham-operated control that received hypoxia only.
OPC Preparation
Mixed glial cultures were prepared as previously reported 29 with some modifications. Briefly, cerebral cortices from P1 Wistar ST rats or SD-Tg (CAG-EGFP) rats expressing the green fluorescent protein (GFP) under control of the CAG promoter were dissected and then digested with 0.25% trypsin (Sigma) and 100 µg/ml DNAase for 20 min at 37 °C. Dissociated cells were isolated using a cell strainer (Falcon 40 µm) and maintained in Dulbecco’s modified Eagle’s medium (DMEM; Thermo Fisher Scientific KK, Tokyo Japan) containing 10% fetal calf serum (FCS) for 7–12 days.
OPCs were obtained from mixed glial cultures as previously reported
18
. Briefly, flasks were shaken at 200 rpm at 37 °C for 20–22 h. Detached cells were collected, and the cells were placed for 1 h on Petri dishes followed by another placement on culture dishes to remove contaminating astrocytes and microglial cells. For cell culture experiments, nonadherent floating OPCs were collected and plated on poly-
OPC Differentiation In Vitro
Plated OPCs were cultured for 24 h in DMEM plus 10% FCS and then expanded in Neurobasal A medium + 2% B27 supplements (NBM/B27) + 1% GlutaMAX (Gibco) containing 10 ng/ml fibroblast growth factor-2 (FGF2) and 10 ng/ml platelet-derived growth factor-AA (PDGF; R&D systems) for 4 days. OLs were differentiated in NBM/B27 medium + 1% GlutaMAX containing 10 ng/ml ciliary neurotrophic factor (CNTF; PeproTech, Texas, USA) and 20 ng/ml triiodothyronin (T3, Sigma). IGF2 (PeproTech) was added to the differentiation medium with/without CNTF and T3.
OPC Transplantation into the Neonatal WMI Model
At P5, 2 days after preparing the neonatal WMI model, pups were anesthetized with isoflurane. An incision was made in the skin and a hole drilled in the soft skull (anterior: 0, lateral: 1.2 mm from bregma). To prepare a glass pipette for OPC grafting, a glass pipette (GC200-10: 2.00 mm O.D., 1.16 mm I.D.; Harvard Apparatus Ltd, Holliston, Cambridge, UK) was pulled with a glass puller (PP-83, Narisige, Tokyo, Japan) to a diameter of OD: 160–180 µm. The glass pipette was connected to a 2.5-µl Hamilton syringe and inserted at the midline into the CC (anterior: 0, lateral: 0, ventral: 2.08 mm from bregma) advancing 2.4 mm from the hole at an angle of 30° to the vertical in the coronal plane. A total of 200,000 OPCs in 2 µl NBM/B27 were grafted at a rate of 0.5 µl/min. The pipette was left in place for 4 min, then withdrawn 500 µm, and left in place for another 4 min. Finally, the pipette was slowly removed and the scalp sutured.
OPC graft was similarly performed at P19 pups. We made a hole in the skull (anterior: 0, lateral: 1.5 mm from bregma), and the glass pipette was inserted into the CC advancing 3.0 mm from the hole at an angle of 30°.
Immunocytochemistry
After differentiation, OPCs were fixed with 4% paraformaldehyde (Sigma-Aldrich, St Louis, USA) in 0.1 M phosphate buffer (PB). After rinsing in phosphate-buffered saline (PBS), OPCs were then incubated in PBS with 0.25% (v/v) Triton (Nacalai tesque, Kyoto Japan), and then blocked with 3% (v/v) goat serum for 30 min. OPCs were incubated with primary antibodies for 1 h and then with appropriate secondary antibodies for 1 h.
The primary antibodies used were rabbit antiplatelet-derived growth factor receptor alpha (PDGFRα, 1:600; Santa Cruz Biotechnology, CA, USA), rabbit anti-NG2 (1:600; Merck Millipore, Germany), mouse anti-2′,3′-cyclic-nucleotide 3′-phosphodiesterase (CNPase, 1:600; Merck Millipore), and mouse antiadenomatous polyposis coli (CC1 clone: 1:250; Merck Millipore). The secondary antibodies were goat anti-mouse Alexa 488 (1:2000; Molecular Probes, Eugene, OR, USA) and goat anti-rabbit Alexa 594 (1:2000; Molecular Probes).
Immunohistochemistry
Rats were deeply anesthetized with pentobarbital (>50 mg/kg, intraperitoneal [i.p.], Kyouritu Seiyaku, Tokyo, Japan) and perfused transcardially with 4% paraformaldehyde in 0.1 M PB. Brains were removed and postfixed overnight followed by cryoprotection in 30% sucrose in 0.1% PB. Coronal sections of 40 µm thickness were cut using a microtome. The sections were washed in PBS, blocked with 3% (v/v) goat serum plus 0.5% (v/v) Triton (Nacalai tesque) in PBS for 1 h, and then incubated with primary antibodies overnight at 4 °C.
The primary antibodies used were rabbit anti-IGF2 (1:250; Abcam, Cambridge, UK), rabbit monoclonal anti-IGF2 R (1:500, clone EPR6599; GeneTex, CA, USA), rabbit anti-PDGFRα (1:250), mouse anti-CC1 (1:250), mouse antiglial fibrillary acidic protein (GFAP, 1:250; Sigma Aldrich, Tokyo, Japan), rabbit anti-IBA1 (1:250; Wako, Osaka, Japan), and chicken anti-GFP (1:1000; Abcam). The secondary antibodies were goat anti-mouse Alexa 594 (1:600; Thermo Fisher Scientific, Tokyo Japan), goat anti-rabbit Alexa 594 (1:600; Thermo Fisher Scientific), and goat anti-chicken Alexa 488 (1:600; Thermo Fisher Scientific).
Statistics
All statistical analyses were conducted using JMP ver.10 (JMP statistical software; SAS Institute). The threshold for significance for all experiments was set at P ≤ 0.05 marked with a single asterisk or P ≤ 0.005 marked with three asterisks. Comparison of Igf2 mRNA expression, immunostaining of oligodendrocyte lineage cells in vitro, and immunostaining of grafted GFP-positive cells were performed using the Mann-Whitney U test. The number of animals used in each experiment can be found in the Results section. All data are plotted as the mean ± standard error of the mean.
Results
IGF2 Expression is Enhanced in the H-I Brain of Neonatal WMI Model Rats
To investigate the factors that can recover myelination failure in the model, 19,20 we focused on IGF2, which may be able to potently induce differentiation of OPCs, similarly to IGF1 30 .
We first investigated the enhanced expression of Igf2 mRNA in the neonatal WMI model at P5 using real-time PCR. Igf2 gene expression of the ipsilateral H-I side was 10-fold higher compared with that in the contralateral side or sham-operated control that received hypoxia only (Fig. 1A).

IGF2 expression is increased in the brain of neonatal WMI model rats. (A) To confirm enhanced expression of Igf2 in the ipsilateral sensorimotor cortex (upper panel, boxed area), mRNA from the ipsilateral sensorimotor cortex (ipsi), and the contralateral sensorimotor cortex (contra) of the model at P5, and from the cortex of sham-operated sensorimotor control P5 rats (control) was measured using real-time PCR. Igf2 expression in the ipsilateral cortex was more than 15 times that of the contralateral side or sham-operated control, indicating specific expression of Igf2 in the neonatal WMI model. (B). Double staining for IGF2 (red) and OLIG2 (green, left panel), or GFAP (green, right panel) was performed. IGF2 co-localized with OLIG2 and GFAP at P5 in the CC. However, few ED1-positive cells and few NeuN-positive cells were seen. Scale bar: 25 μm. (C). Double staining for IGF2 R (red) and OLIG2 (green, left panel), or GFAP (green, right panel) was performed. IGF2 R was detected in OLIG2-positive cells and in GFAP-positive cells. Arrows in the picture indicate typical double-stained cells with yellowish color. Scale bar: 25 μm. CC: corpus callosum; GFAP: glial fibrillary acidic protein; IGF2: insulin-like growth factor 2; PCR: polymerase chain reaction; WMI: white matter injury.
We next performed immunostaining for IGF2 and IGF2 R with double staining for NeuN, GFAP, Olig2, or ED1 to determine which cell types express IGF2 and IGF2 R in the brain of neonatal WMI model rats (Fig. 1B, C). In the CC of neonatal WMI rats, in which OL lineage cells are dominant, IGF2 staining colocalized with OLIG2 and GFAP at P5 but not for ED1 or NeuN (Fig. 1B). IGF2 R was also detected in OLIG2-positive cells and GFAP-positive cells (Fig. 1C), indicating an autocrine or paracrine effect of IGF2 in OLIG2-positive cells and GFAP-positive cells. In the cortex, in which neurons and proliferating astrocytes are dominant at P5, IGF2 was colocalized with GFAP and NeuN but not with ED1 or OLIG2 (data not shown). IGF2 R was detected in NeuN-positive cells and OLIG2-positive cells (data not shown), indicating a different cell response in OLIG2-positive cells to IGF2 in the cortex.
IGF2 Induces OPC Differentiation In Vitro
To investigate the effect of IGF2 on OPCs in vitro, OPCs were prepared from mixed glial cultures and expanded with PDGF-AA (10 ng/ml) and FGF2 (10 ng/ml) for 5 days. IGF2 (10 ng/ml) was then added in the presence of CNTF (10 ng/ml) and T3 (20 ng/ml) and incubation continued for 6 days in vitro (DIV) (Fig. 2A). Cells were immunostained for markers of early OL progenitors (NG2 and PDGFRα) and for markers of mature OLs (CC1 and CNPase; Fig. 2B).
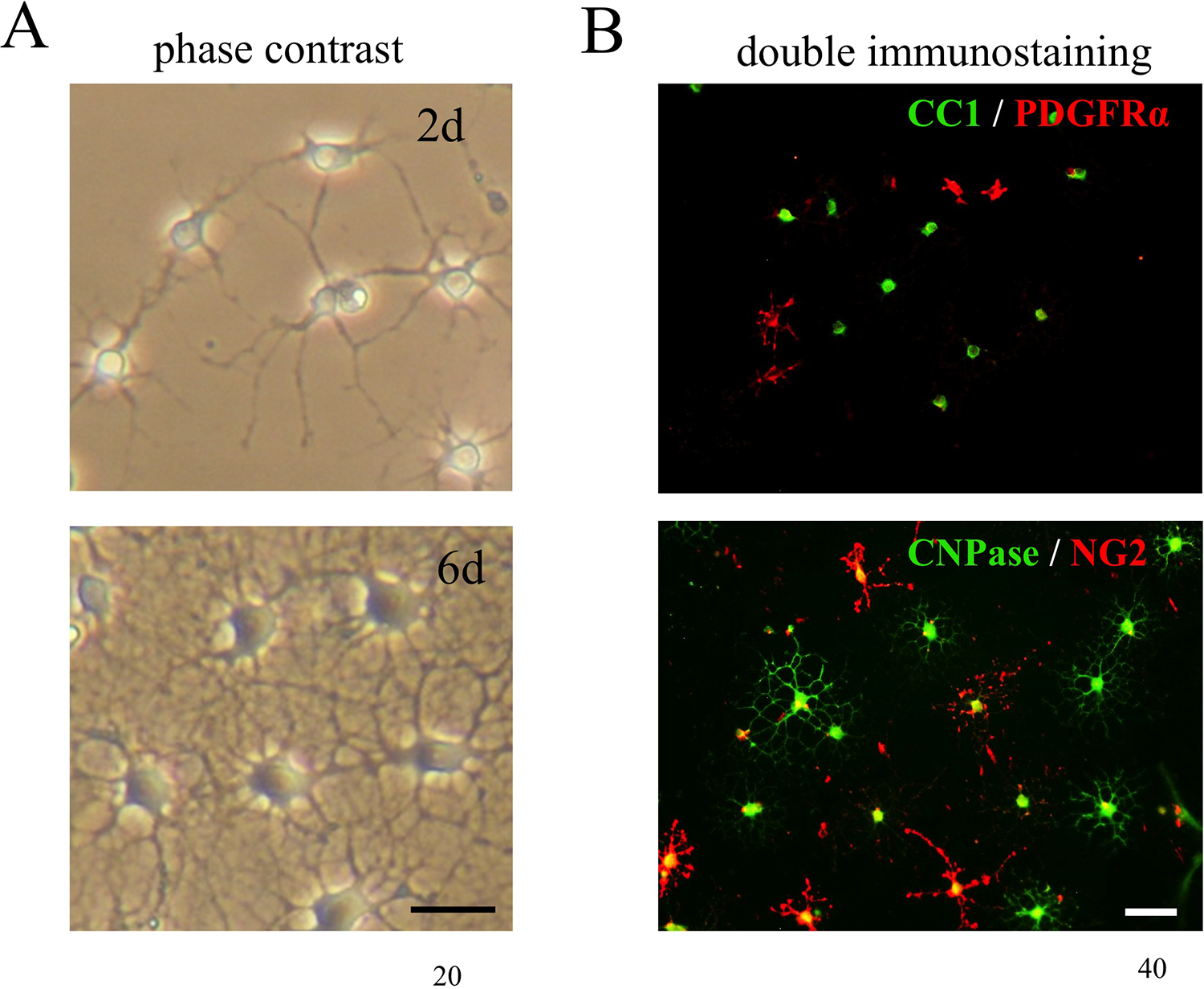
Cell culture and immunostaining of OL lineage cells. (A) OPCs were prepared from mixed glial cultures and expanded with PDGF-AA and FGF2 for 5 days. Cells were treated with IGF2 for 6 days in the presence of CNTF and T3. Under a phase-contrast microscope, small cell bodies with several neurites were observable after 2 DIV and typical mature OLs were seen after 6 DIV. Scale bar: 20 μm. (B) Double staining with a maker for early OL progenitors and a marker for mature OLs (upper panel): double staining of PDGFRα (red) and CC1 (green) (lower panel): double staining of NG2 (red) and CNPase (green). The numbers of NG2-positive cells, PDGFRα-positive cells, CNPase-positive cells, and CC1-positive cells were counted at 2 DIV (Fig. 3A) and at 6 DIV (Fig. 3B). Scale bar: 40 μm. CC, corpus callosum; CNPase, 2′,3′-cyclic-nucleotide 3′-phosphodiesterase; CNTF: ciliary neurotrophic factor; DIV: days in vitro; FGF: fibroblast growth factor-2; IGF2: insulin-like growth factor 2; OL: oligodendrocytes; OPC: oligodendrocyte progenitors; PDGFRα: platelet-derived growth factor receptor alpha.
After 2 DIV, the percentage of NG2-positive cells was significantly decreased by IGF2 treatment (control: 52.0 ± 3.9%; IGF2: 27.1 ± 2.9%, n = 8, P < 0.005; Fig. 3A, upper left graph) and that of PDGFRα-positive cells was also decreased (control: 49.1 ± 6.1%; IGF2: 30.8 ± 2.4%, n = 7, P < 0.05; Fig. 3A, upper right graph). However, the percentage of CNPase-positive cells was significantly increased (control: 46.4 ± 4.6%; IGF2: 65.7 ± 5.7%, n = 8, P < 0.05; Fig. 3A, lower left graph) and that of CC1-positive cells was significantly increased by IGF2 addition (control: 52.1 ± 4.2%; IGF2: 61.9 ± 3.5%, n = 8, P = 0.05; Fig. 3A, lower right graph).

IGF2 induces OPC differentiation in vitro. (A) Number of OL-marker-expressing cells at 2 DIV in the presence of CNTF and T3. The number of NG2-positive cells and PDGFRα-positive cells were significantly decreased by IGF2 treatment (n = 8 for NG2 experiment and n = 7 for PDGFRα experiment). However, the numbers of CNPase-positive cells and CC1-positive cells were significantly increased by IGF2 (n = 8 for CNPase and CC1 experiments). **P < 0.005 and *P < 0.05. (B) Number of OL-marker-expressing cells after 6 DIV in the presence of CNTF and T3. The number of NG2-positive cells was significantly decreased (n = 8), while the number of CNPase-positive cells and CC1-positive cells were significantly increased by IGF2 (n = 8 for CNPase experiment and n = 7 for CC1 experiment). A tendency of decreased cell number was shown by PDGFRα-positive cells after IGF2 treatment (P = 0.06). *P < 0.05. CNPase: 2′,3′-cyclic-nucleotide 3′-phosphodiesterase; CNTF: ciliary neurotrophic factor; DIV: days in vitro; FGF: fibroblast growth factor-2; IGF2: insulin-like growth factor 2; OL: oligodendrocytes; OPC: oligodendrocyte progenitors; PDGFRα: platelet-derived growth factor receptor alpha.
After 6 DIV, similar results were observed (Fig. 3B): the percentage of NG2-positive cells in total cells (control: 35.2 ± 2.1%; IGF2: 20.9 ± 2.3%, n = 8, P < 0.005; Fig. 3B, upper left graph) and PDGFRα-positive cells in total cells (control: 36.4 ± 4.0%; IGF2: 23.3 ± 3.2%, n = 7, P = 0.06; Fig. 3B, upper right graph) were decreased by IGF2 administration, while the percentage of CNPase-positive cells in total cells (control: 66.6 ± 2.9%; IGF2: 77.3 ± 2.4%, n = 8, P < 0.05; Fig. 3B, lower left graph) and CC1-positive cells (control: 62.6 ± 4.2%; IGF2: 74.6 ± 3.7%, n = 7, P < 0.05; Fig. 3B, lower left graph) were increased. Thus, IGF2 can promote the differentiation of OPCs into mature OLs.
To further investigate whether a single IGF2 treatment can induce OPC differentiation, expanded OPCs were allowed to differentiate with IGF2 (100 ng/ml) alone, that is, without CNTF and T3 (Fig. 4). The percentage of PDGFRα-positive cells was decreased by IGF2 treatment (control: 82.4 ± 2.1%; IGF2: 68.5 ± 1.2%, n = 4, P < 0.05), while the percentage of CC1-positive cells was increased (control: 6.0 ± 1.2%; IGF2: 18.0 ± 3.3%, n = 4, P < 0.05) after 2 DIV (Fig. 4A). However, the percentages of PDGFRα-positive cells and CC1-positive cells in total cells were similar after 6 DIV: PDGFRα-positive cells were 50.8 ± 6.7% in control and 48.8 ± 4.7% with IGF2 treatment (n = 4, P = 0.90) and CC1-positive cells were 44.1 ± 6.9% in control and 48.9 ± 4.9% with IGF2 treatment (n = 4, P = 0.69; Fig. 4B).

Single IGF2 treatment induces OPC differentiation. OPCs were induced to differentiate in vitro with 100 ng/ml IGF2 in the absence of CNTF and T3. (A) Number of PDGFRα-positive cells and CC1-positive cells after 2 DIV without CNTF and T3. The percentage of PDGFRα-positive cells (n = 4) was significantly decreased by IGF2 treatment, while the percentage of CC1-positive cells (n = 4) was significantly increased after 2 DIV. *P < 0.05. (B) Number of PDGFRα-positive cells and CC1-positive cells after 6 DIV without CNTF and T3. There was no significant difference in the number of PDGFRα-positive cells or CC1-positive cells between groups (n = 4 each group) after 6 DIV. CC: corpus callosum; CNPase: 2′,3′-cyclic-nucleotide 3′-phosphodiesterase; CNTF: ciliary neurotrophic factor; DIV: days in vitro; IGF2: insulin-like growth factor 2; OPC: oligodendrocyte progenitors; PDGFRα: platelet-derived growth factor receptor alpha.
Transplantation of OPCs into the Neonatal WMI Model Rat Brain
OPCs prepared from GFP-expressing rat-derived mixed glial cultures were grafted into the CC of neonatal WMI model P5 rats, which was 2 days after receiving H-I brain damage. To prevent the backflow of grafted cells, the pipette was accessed to the target point with an angle of 30° to the vertical in the coronal plane. To determine if the grafted OPCs can survive in the brain of the model, GFP-positive cells were counted at 2 and 8 weeks after transplantation. Due to the technical difficulty of our experiments, data of the pattern of cell-surviving place and of the number of grafted cells were big variation as follows.
At 2 weeks after grafting, GFP-positive cells were observed in most rats even without immunosuppression; transplanted cells were detected in 17 of 19 model rats and in 15 of 17 sham-operated rats. Although most cells were located around the injection site (CC and the cortex), some grafted cells were detected in remote areas, such as the external capsule (Fig. 5A).
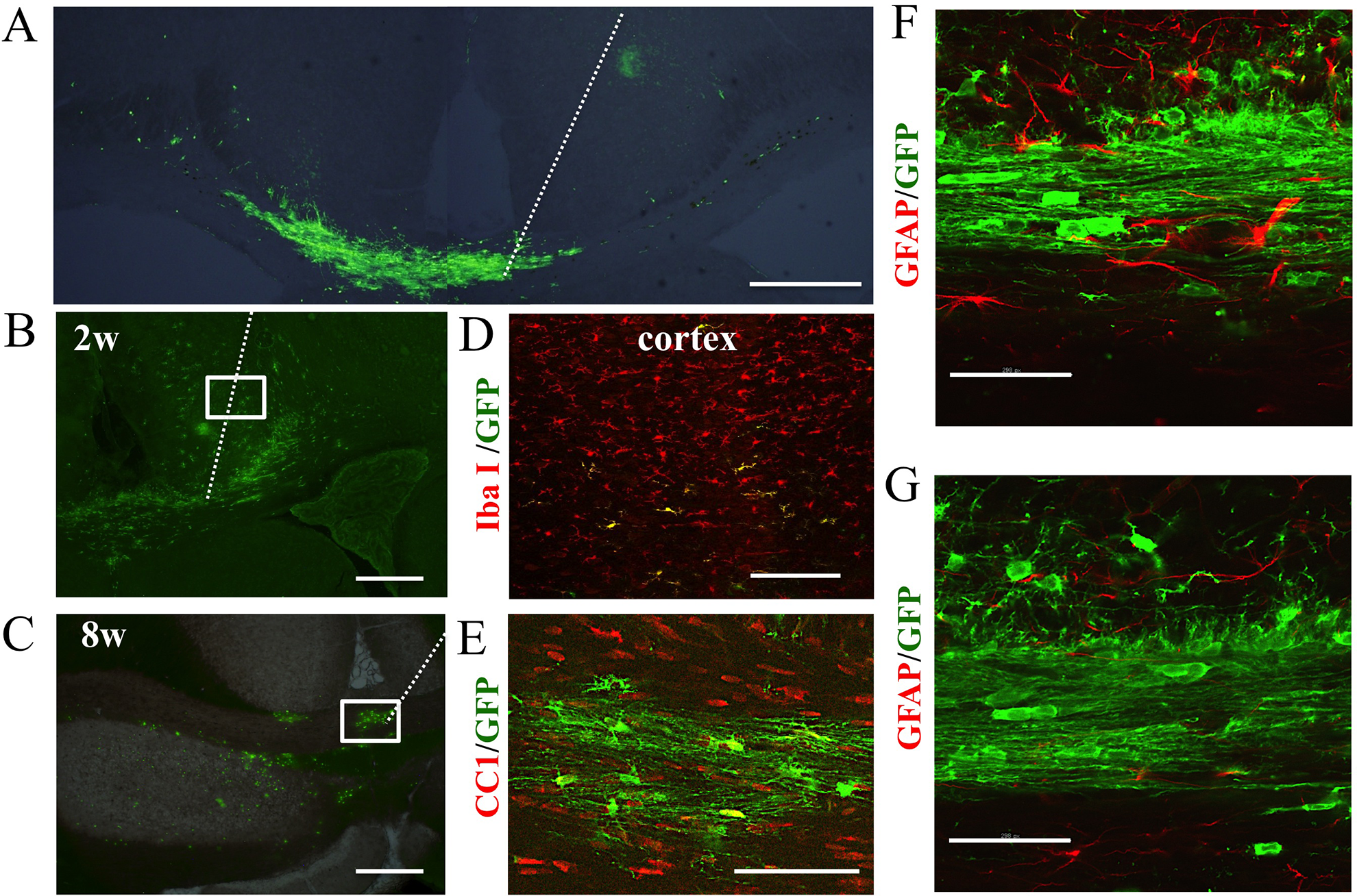
Transplanted GFP-positive OPCs expressing CC1 can survive in the CC for at least 8 weeks. (A) GFP-expressing rat-derived OPCs were grafted into the brain of P5 neonatal WMI model rats. At 2 weeks after grafting, GFP-positive cells were observed in most rats without immunosuppression: 17 of 19 model rats and 15 of 17 sham-operated rats. Most cells remained around the injection area of the CC and the cortex, while some cells were detected in remote areas. Low magnified picture (0.5×) of the coronal section of 0.0 mm from bregma. White dotted line: track of the pipette for transplantation. Scale bar: 0.5 mm. (B and D) GFP-positive cells were detected in the CC and cortex of most of the model rats at 2 weeks after grafting (B). Although most of the grafted cells in the CC expressed CC1, most of the grafted cells in the cortex co-expressed IBA1 (D). The number of surviving cells in the CC varied among animals (152–1819 cells, n = 8). However, the number of surviving cells in the model was similar on both sides of the CC (right-CC: 310.0 + 81.4 cells; left-CC: 403.0 + 168.0 cells). Picture of the coronal section of 0.0 mm (B) or −1.8 mm (D) from bregma. White dotted line: track of the pipette for transplantation. Scale bar in B: 0.5 mm, D: 100 μm. (C and E) At 8 weeks after transplantation, GFP-positive cells were detected in model rats despite the lack of immunosuppression (C). Co-expression of CC1 (red) and GFP (green) was observed in the CC, showing the myelin-like structure along the fibers (E). Scale bar in C: 0.5 mm, E: 100 μm. (F and G) Double staining for GFAP and GFP was performed in the CC of the model rats at 2 weeks after grafting, revealing that the percentage of GFAP-positive cells in GFP-positive grafted cells was low in the CC at P19. Scale bar in F and G: 100 μm. CC: corpus callosum; GFAP: glial fibrillary acidic protein; GFP: green fluorescent protein; OPC: oligodendrocyte progenitors.
In 11 rats of 17 WMI model rats, GFP-positive cells were detected in both the CC and the cortex (Fig. 5B), although in two rats, GFP-positive cells were only detected in the CC and in four rats GFP-positive cells were only detected in the cortex. In six sham-operated rats (n = 13), GFP-positive cells were found in both the CC and the cortex; however, GFP-positive cells were only observed in the CC in one rat and only in the cortex in six rats. The total number of surviving grafted cells in the CC was very varied among individual animals (152–1819 cells, n = 8), but the number was similar on both sides of the CC (right-CC: 310.0 + 81.4 cells; left-CC: 403.0 + 168.0 cells) in the neonatal WMI model.
At 8 weeks after transplantation, GFP-positive cells were detected in all investigated neonatal WMI (n = 3) and sham (n = 4) rats despite the lack of immunosuppression (Fig. 5C). The shape of the positive cells resembled that of mature myelinating OLs (Fig. 5E).
Differentiation of Transplanted OPCs in the Rat Brain
To determine whether grafted OPCs can differentiate into mature OLs in the brains of neonatal WMI model rats, GFP-positive transplanted cells were co-stained with CC1, a marker for mature OLs, or PDGFRα, a maker for immature OLs, at 2 weeks after grafting. The ratio of GFP to CC1 and GFP to PDGFRα positive cells was investigated in the brain slice containing the largest number of GFP-positive cells.
The percentage of CC1-positive cells in all GFP-positive cells (Fig. 6C) was 49.3 ± 3.3% in the CC of neonatal WMI rats (n = 11), which was significantly lower than that in sham-operated control rats (67.5% ± 5.8%, n = 7, P < 0.05; Fig. 6C). However, a small percentage of the grafted cells was PDGFRα-positive (Fig. 6B) in the CC of the model rats (9.9 ± 2.1% of GFP-positive cells, n = 11), which is comparable to that of sham-operated control rats (6.5± 2.4% of GFP-positive cells, n = 7; Fig. 6B). In addition, the percentage of GFAP-positive cells in GFP-positive grafted cells was low in the CC at P19 (Fig. 5F and G). Thus, the differentiation of grafted OPCs was unexpectedly inhibited in the CC of neonatal WMI model rats compared with sham-operated control rats.

Transplanted OPCs show a lack of maturation in neonatal WMI model rat brains. (A) To investigate whether grafted OPCs differentiate into mature OLs; GFP-positive transplanted cells were co-stained with PDGFRα or CC1 at 2 weeks after grafting. Grafted GFP-positive cells became CC1-positive cells (yellow nucleus in the right merged picture) with decreased expression of PDGFRα (green cell body in left merged picture). Scale bar: 100 μm. (B and C) The percentage of PDGFRα-positive cells (B) or CC1-positive cells (C) among GFP-positive cells was investigated at 2 weeks after transplantation. The percentage of CC1-positive cells was significantly lower in the neonatal WMI model (n = 11) compared with that in the sham-operated control (n = 7). (C). However, there was no significant difference in the number of PDGFRα-positive cells in sham-operated control and the neonatal WMI model rats. *P < 0.05. CC1: adenomatous polyposis coli; GFP: green fluorescent protein; OL: oligodendrocytes; OPC: oligodendrocyte progenitors; PDGFRα: platelet-derived growth factor receptor alpha.
The differentiation pattern of the grafted cells was completely different in the cortex; most GFP+ cells were co-stained with IBA1, and a few cells were co-stained with CC1 or PDGFRα (Fig. 5D).
Later Time Point of Grafting OPCs
To investigate if later time point of OPC grafting may solve the problem of inhibited differentiation, OPCs were grafted into the CC of neonatal WMI model P19 rats, which was 16 days after H-I in low inflammatory environment.
At 2 weeks after grafting (P33), GFP-positive cells were detected in four of 10 model rats and in four of seven sham-operated rats. The number of surviving grafted cells in the CC was similar between the WMI model rat (90.3 ± 5.7, n = 4) and sham-operated rat (90.0 ± 24.1, n = 4). The percentage of CC1-positive cells in all GFP-positive cells was 60.0 ± 2.3% in the CC of neonatal WMI rats (n = 4), which was significantly lower than that in sham-operated control rats (70.1.5% ± 1.6%, n = 4, P < 0.05).
Discussion
In this study, we investigated whether IGF2, whose gene is upregulated in neonatal WMI, 19 exhibits a trophic effect on OPCs, and we also evaluated OPC grafts as cell replacement therapy for neonatal WMI, in which OPCs are damaged and lost because of H-I in the developing brain. Specific enhancement of Igf2 mRNA expression in the brain of neonatal WMI rats was confirmed by real-time PCR. A trophic effect of IGF2 on OPCs was shown in vitro; a single dose of 100 ng/ml IGF2 is enough to induce OPCs to differentiate into mature OLs. OPCs derived from a GFP-expressing rat can survive for at least 8 weeks after transplantation into the CC without the immunosuppressive drug. The ratio of mature OLs in the grafted cells was significantly decreased in neonatal WMI model rats in comparison with sham-operated rats.
Hypothesis of OPC Maturation in a rat Model of Neonatal WMl
There are two major responses in our neonatal WMI model that disrupt OPC maturation and myelination in developing white matter. The first response is an acute phase characterized by the targeted death of OPCs due to H-I or inflammatory mediators. The second response is a chronic phase characterized by a significant increase in proliferation and arrested maturation of the remaining OPCs, resulting in impaired myelination 8,31 –33 .
Effects of reactive astrocytes and active microglia are probably important in the impaired OPC maturation and myelination in the chronic phase of this model. Accumulation of astrocyte-derived hyaluronic acid (HA) and its receptor, CD44, inhibits OPC differentiation and myelination in a WMI model 34 –37 . Active microglia in the brain are maintained in a proinflammatory state, 38 –40 which disrupts proliferation and differentiation of immature OLs through increased levels of proinflammatory cytokines, such as tumor necrosis factor alpha (TNFα), interleukin (IL)1β, IL2, and IL17 41 –45 .
However, we hypothesized that trophic factor(s)/substances(s) are newly expressed or enhanced in the brain of neonatal white matter where endogenously generated OPCs are damaged by H-I. To discover trophic factors that promote survival/differentiation of grafted OPCs, we previously performed a cDNA microarray study, 19 which showed that IGF2 expression is enhanced in the brain of neonatal WMI model rats. In this study, we confirmed the increase of IGF2 using real-time PCR and the effect of IGF2 on OPCs in vitro.
The Effect of IGF2 on OPCs In Vitro
IGF2 stimulates the differentiation of multipotent neural progenitor cells into OLs 46 ; therefore, it is likely that increased IGF2 expression in the brain of the model rats promotes OL survival/differentiation. We confirmed the effect of IGF2 on OPC differentiation in cultured OPCs; IGF2 directly induced OPC differentiation without the support of CNTF and T3. IGF2 and its receptor IGF2 R were expressed in OLIG2-positive cells of the CC and the cortex, suggesting autocrine and paracrine effects of IGF2. IGF2 can weakly bind to the IGF1 receptor as well as strongly bind to IGF2/mannose-6-phosphate receptor 47 ; therefore, IGF2 probably activates extracellular signal-regulated kinase and Akt/mammalian target of rapamycin pathways in OPCs, similarly to IGF1, which promotes OL differentiation and myelination 48 –50 . Our preliminary data showing that IGF2 has no additive effect with IGF1 on OPC growth 48 –50 support this idea.
It is known that nasal administration of IGF1 protects from injury and improves neurological dysfunction in a lipopolysaccharide-induced WMI injury model 51 . The question of why IGF2 rather than IGF1 is enhanced in the brain of neonatal white matter is interesting, but the data from this study do not answer this question. It is possible that the mechanism for IGF2 expression is activated by H-I in the postnatal developing brain where astrocytes and OLs are produced and functionally immature, as the expression of Igf2 mRNA in Fig. 1A seems to have a tendency to depend on the strength of H-I (ipsilateral side > contralateral side >> control).
Long-Term Survival of Grafted OPCs in the White Matter without Immunosuppression
Cell therapy is widely discussed as a promising treatment for brain injury 52 . The use of stem cells, OPCs, and other precursor cells improves neurobehavioral disorders because these cells have been shown to secrete neuroprotective factors despite definite cell survival 24,27,53 . Similarly, grafted OPCs showed a tendency to increase endogenous OPCs (GFP-negative and CC1-positive cells) in the CC in our preliminary experiment. We investigated whether OPC transplantation into our neonatal WMI model is effective at replacing lost OPCs, relating to a possibility for improvement of disturbed motor function.
We showed that grafted cells could survive and differentiate into mature OLs only in the CC, but not in the cortex, for at least 8 weeks without immunosuppression. This phenomenon was observed in both neonatal WMI model and sham-operated control rats, indicating that the environment of the cortex is different from that of the white matter and is not suited for the survival of OPCs and that immunotolerance in the developing brain is related to the long-term survival of grafted OPCs. It was noted in our preliminary data that OPC transplantation with cyclosporine induces functional recovery of motor function in the model. Thus, OPC transplantation into neonatal WMI model may be effective from the aspect of cell replacing lost OPCs.
Differentiation of Grafted OPCs into Mature OLs is Reduced in Neonatal WMI Model Rats
As IGF2 can induce OPCs to mature into OLs, we predicted that greater numbers of grafted OPCs would differentiate into CC1-positive cells in the brain of neonatal WMI model rats compared with that in sham-operated control rats. However, fewer grafted CC1-positive cells were present in the CC of neonatal WMI model rats than in sham-operated control rats. It is probable that grafted OPCs cannot proliferate/survive/differentiate in the CC (where IGF2 is produced and acting) because of reactive astrocytes and active microglia. Astrocyte-derived HA and CD44 inhibit OPC differentiation 36 –39 and proinflammatory cytokines, such as TNFα, IL1β, IL2, and IL17 from active microglia, disrupt the proliferation of immature OPCs 41 –45 .
It is noteworthy that an inhibitory mechanism as same as trophic substance(s) is important for replacing damaged or lost OPCs in the brain of neonatal WMI rats. Our findings also suggest that a strategy to support damaged OPCs by trophic factor(s)/substances(s) during the acute phase may be less effective. Rather, the focus should be directed toward a strategy to promote the differentiation and myelination of pre-OLs during the chronic phase of the neonatal WMI model. This notion is also supported by later time point of OPC grafting revealing that the number of mature OLs was unexpectedly small in the OPC-grafted CC as compared with that in PBS-injected control.
Footnotes
Acknowledgments
Ethical Approval
This study was approved by the Committee of Animal Experimentation of Nagoya City University Medical School, Nagoya, Japan.
Statement of Human and Animal Rights
Animal care and all experimental procedures were conducted according to the guidelines of the Institute for Experimental Animal Science, Nagoya City University Medical School, Japan, and every effort was made to minimize the pain and discomfort of the animals used.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: this study was supported by Grants-in-Aid for Scientific Research (# 26430020 and # 18K07882 to HH, #16K10100 to SM) and Grants-in-Aid for Early-Carrier Scientist (#19K17337 to SO) from the Japan Society for the Promotion of Science (JSPS). This study was partly supported by a Grant-in-Aid for Scientific Research on Innovative Areas (Adaptive Circuit Shift) to HH.