
Abstract
Osteoporotic fracture is the main complication of osteoporosis (OP) and accounts for millions of injuries annually. Local intervention by intra-marrow injection has been a good option for preventing osteoporotic bone loss when the osteoporotic femoral fracture has been treated. In this study, tail vein transplantations were examined to evaluate the cell-based therapeutic approach for treating OP with adipose-derived stem cells (ADSCs) and platelet-rich fibrin releasates (PRFr) in an ovariectomized (OVX) mice model. Thirty-six 12-wk-old female ICR mice were randomly divided into six groups: untreated control; sham-operated; OVX-control; OVX-ADSCs; OVX-PRFr; and OVX-ADSCs+PRFr. Starting 8 wk after ovariectomy, the OVX mice received tail vein injections once each week for four consecutive weeks, then were evaluated radiographically and histopathologically 8 wk after the first injection. We also assessed changes to bone trabeculae in the proximal tibial growth plate. In OVX mice treated with ADSCs or PRFr alone, or with a combination of ADSCs and PRFr, the trabecular bone mineral density (BMD), bone volume ratios (BV/TV), and numbers (Tb.N) in the proximal tibia areas were significantly higher than that in the OVX-control group. Significant differences between OVX-treated mice and OVX controls were found for trabecular separation, but not for trabecular thickness. These results indicate that ADSCs or PRFr treatment enhances bone microarchitecture in OP. The treatment of bone loss of OVX mice with ADSCs+PRFr induced greater bone consolidation with bone tissue production (P < 0.01) when compared to the others. Thus, we conclude that the transplantation of ADSCs combined with PRFr might provide an alternative strategy for the treatment of various bone disorders in OP with an unlimited source of cells and releasates.
Keywords
Introduction
Osteoporosis (OP) is a systemic skeletal disease characterized by low bone mineral density (BMD) and microarchitecture deterioration of bone tissue, leading to bone fragility and an increased risk of fracture 1 . Loss of estrogen seems to be the most important mechanism in the development of OP. The use of ovariectomized (OVX) rat or mice models is a well-established and reproducible method of simulating the postmenopausal condition and has been found effective in mimicking postmenopausal cancellous bone loss over a relatively short period of time 2 . During the initial period after ovariectomy, bone resorption exceeds bone formation causing a net loss of bone 3 . Statistically significant bone loss is detectable in the proximal tibial metaphysis after 14 d 3,4 , in the lumbar vertebral body after 60 d 5 .
Currently, bisphosphonates 6 , hormone therapy with estrogen 7 , selective estrogen receptor modulators/estrogen agonist-antagonists 8 , denosumab 9 , salmon calcitonin 10 , and parathyroid hormone analogs 11 are the drugs of choice for OP 12,13 based on the goal of blunting the resorption component of bone homeostasis, a tightly coupled process of bone formation and bone resorption 14 . While professional organizations have compiled recommendations for the management of OP in various populations, a consensus has yet to develop as to which is the gold standard. These medications have yet to fully reduce the number of fracture cases and are associated with several side effects 15 . As the population continues to age and some types of medicines can cause bone loss, OP is likely to become an even more prevalent and serious health condition with economic burden for the society 16 . Therefore, a safer and more effective therapeutic strategy is urgently needed.
Bone remodeling is a complex process involving the close interactions between different progenitor cell lineages, mainly osteo- and adipo-progenitor cells 17 . It has been thoroughly proven that osteoblasts and adipocytes are both derived from mesenchymal stem cells within the bone marrow. Signaling pathways involved in these two processes could be modified to favor osteogenesis, resulting in the amelioration of OP 18 . Previous study has indicated that the increase in the number of adipocytes subsequently induced further apoptosis in osteoblasts and promoted the proliferation and differentiation of osteoclasts, resulting in an increase in bone resorption and overall bone loss 19 . Regenerative medicine approaches could be a better option in treating OP and/or osteoporotic damages compared with regular surgical techniques 20 . Therefore, bone marrow transplantation 21 and bone marrow cell intratibial injections 22 have been considered to enhance osteoporotic prevention or osteoporotic bone repair in animal models 23 . Thereby stem cell therapy for OP could potentially reduce the susceptibility of fractures and augment lost mineral density by either increasing the numbers or restoring the function of resident stem cells that can proliferate and differentiate into bone-forming cells 24 . Such OP therapies can be carried out by the exogenous introduction of mesenchymal stem cells (MSCs), typically procured from bone marrow, adipose, and umbilical cord blood tissues or through treatments with growth factors, drugs, or small molecules that recruit endogenous stem cells to osteoporotic sites 24 .
Platelet concentrates for topical and infiltrative use—commonly termed platetet-rich plasma (PRP) or platelet-rich fibrin (PRF)—are commonly used or tested as surgical adjuvants or regenerative medicine preparations in most medical fields, particularly in orthopedic surgery, cardiovascular surgery, plastic/reconstructive surgery, sports medicine, dentistry, and oral implantology 25 . The concept is that within our blood there are growth factors, healing proteins, cell signals, and immune cells, which are part of the normal healing process that can be concentrated and applied or injected to a wound or surgical site to potentially enhance healing. Besides, a PRF clot that is squeezed to extract the releasate PRF (the serum exudate expressed from the clot) is called a PRF releasates (PRFr), and is rich in growth factors, leukocytes, lipids, and also proteins such as vitronectin and fibronectin 26 . This exudate may be used to hydrate graft materials, rinse the surgical site, and store autologous graft. In addition, it has been shown that the concentrations of growth factors such as platelet-derived growth factor (PDGF)-AB, transforming growth factor (TGF)-β1, epidermal growth factor (EGF), basic fibroblast growth factor (b-FGF), vascular endothelial growth factor (VEGF), and P-selectin (a marker of platelet activation) were significantly higher (P < 0.05; < 0.05; < 0.001; < 0.05; < 0.001; < 0.01, respectively) in PRFr than in PRP 27 . In vitro study has demonstrated that autologous growth factors released by PRF gradually expressed stronger and more durable effect on proliferation and differentiation of rat osteoblasts than PRP 28 .
As in previous reports, the observation that increased bone marrow fat content is closely associated with the development and/or consequence of OP 16 demonstrated that PRP treatment mainly exerted its action via promoting bone regeneration but also appeared to suppress adipogenesis within the marrow. Additional pertinent observation has demonstrated that PRP could stimulate the differentiation of embryonic fibroblasts into osteoblast-like cells and transplantation of these PRP-treated cells could significantly improve the bone architecture in OVX-SAMP8 osteoporotic mice 29 .
Our previous study has provided an alternative method for functional reconstruction of rabbit osteochondral defects with an unlimited source of cells and blood-derived fibrin releasates 30 . We hypothesize that PRFr serve as a mixture of growth factors and act as a unique source/carrier of stem cells 27,31 , which could induce osteogeneic differentiation and enhance osteogenesis, thereby favoring bone formation and ameliorate OP. The goal of this study was to develop a method to stimulate bone formation in order to restore proper bone remodeling process and explore the potential application of PRFr for promoting osteogenesis of progenitor cells within the adipose-derived stem cells (ADSCs) using an osteoporotic mouse model.
Materials and Methods
Culture and Identification of Mouse ADSCs
A total of 5.0 g of subcutaneous adipose tissue was extracted from the ventral sides of 8-wk-old female ICR mice (also an albino strain, was named after the Institute of Cancer Research in the USA; BioLASCO Co., Ltd., Yilan, Taiwan) following CO2 euthanasia. It was then immediately washed with phosphate-buffered saline (PBS) containing 1% penicillin/streptavidin (15140-122, Gibco, Santa Clara, CA, USA). Adipose tissue biopsies were chopped finely and adipocytes were isolated by collagenase digest (PBS solution, containing 0.2% collagenase type I, C0130, Sigma-Aldrich, St Louis, MO, USA) in a shaking water bath at 37°C for 30 min. After collagenase digest, the cell suspension was passed through a 100-µm nylon mesh and centrifuged at 500 × g for 10 min. The supernatant was discarded and the cell pellet was resuspended in Minimum Essential Medium Eagle Alpha Modification (M0894, Sigma-Aldrich) containing 10% fetal bovine serum (FBS) and added into 100-mm dishes for culture in a humidified 5% CO2 incubator at 37°C. Culture medium was replaced 2 d later and once every 2 d thereafter. When the cells were 80% to 90% confluent, they were digested with trypsin-ethylenediaminetetraacetic acid (EDTA) solution (0.05% trypsin, 0.5 mM EDTA, T4174, Sigma-Aldrich) and plated at a density of 3.3 × 105 cells per dish for further culture.
For molecular characterization, mouse ADSCs at passage 2 were harvested and resuspended in a culture medium at a density of 1 × 107 cells/ml. These cells were analyzed for expression of surface markers CD34, CD45, CD73, and CD90 by flow cytometry. Mouse ADSCs were stained with the rat anti-mouse CD34 (553731, BD Biosciences, San Jose, CA, USA) and CD73 (550738, BD); rabbit anti-mouse CD45 (ab10558, Abcam, Cambridge, MA, USA); and mouse anti-mouse CD90 (554895, BD). Cells (1 × 106) were incubated with the appropriate antibody in 0.1 ml PBS for 60 min at 4°C. Then, cells were washed and stained with phycoerythrin-conjugated goat anti-mouse immunoglobulin (Ig) (550589, BD), Alexa Fluor 647-conjugated goat anti-rat IgG (ab150159, Abcam), and DyLight 488-conjugated donkey anti-rabbit IgG (SA5-10038, Thermo, Waltham, MA, USA) secondary antibodies at 4°C for 30 min. Subsequently, cells were washed twice with PBS, centrifuged, then the final cell pellet was resuspended in fluorescence-activated cell sorting (FACS) sorting buffer (PBS, 1 mM EDTA, 25 mM HEPES, 1% FBS, pH 7.0), and filtered using a Falcon 35 µm cell strainer (352235, BD) before sorting. ADSCs were done on a BD FACS Aria cell sorter and the data were analyzed using BD FACSDiva software 32,33 .
Xenogenic PRFr Preparation
A laboratory mouse has a circulating blood volume of about 1.5 to 2.5 ml (6% to 8% of the body weight) 34 , making it difficult to get enough volume of mouse blood. Since rabbits are of the order lagomorphs and similar to rodents, large lot sizes available from pooled rabbit donors may be an alternative blood source35. Blood samples were collected from the experimental rabbits under general anesthesia. After adequate skin cleaning, disinfection, and sterilization, 6 ml of blood was drawn from the right jugular vein of each animal and collected in vacutainer tubes (367988, BD) with clot activator and gel. The vacutainer was rotated gently for 30 s and centrifuged at 3000 × g for 10 min. The resultant jelly-like PRF needed to be separated from the gel clot without the red blood cells sinking to the bottom of the tube. Using forceps, the PRF clots were transferred into 15-ml sterile centrifuge tubes and the jelly-like PRF allowed to stand for at least 5 h 30 . Following centrifugation of the PRF at 5000 × g for 10 min, the supernatant PRFr was collected in sterile tubes, frozen, and stored at –20°C until use. All preparation and delivery steps were carried out under standard disinfection procedures. One milliliter of whole blood yielded about 0.2 ml of PRFr in general.
Experimental Animals, Surgical Procedure, and ADSCs and/or PRFr Injection
All experiments were performed with the approval of the Institutional Animal Care and Use Committees of National Taiwan University (102-EL-82) and were carried out in accordance with the International Council for Laboratory Animal Science guidelines for the care and use of laboratory animals 36 . All efforts were made to minimize animal suffering and the number of animals used. After assessment of the anesthetic plane, back hair elimination, and disinfection, an approximately 0.5 cm linear incision (parallel to the spine) was made at 0.5 cm from the spine and 1 cm below the ribs. After the removal of bilateral ovaries, the abdominal muscles were then closed followed by the skin using an absorbable suture (Vicryl 3.0, ETHICON Inc., Somerville, NJ, USA). The same procedure was performed for the animals designed to the sham group but without ovary removal. Animals were then observed twice a day until healing of the incision had occurred.
OVX female mice (12 wk of age) were used for the experiments. Mice were then divided into six groups with six mice in each group: Control (Ctrl), Sham, OVX-Ctrl, OVX-ADSCs, OVX-PRFr, and OVX-ADSCs+PRFr. Serving as controls were unoperated animals and sham-operated animals who underwent the same manipulations as the OVX mice but without ovary removal. Injections consisting of ADSCs and/or PRFr were prepared in vitro under general sterile conditions and collected in a 1-ml syringe. Eight weeks after ovariectomy 37,38 , the experimental mice received tail vein injections through a 26-gauge needle once each week for four consecutive weeks (quadruple-injection). The OVX-ADSCs group received 3 × 105 ADSCs suspended in 0.6 ml of PBS and the OVX-PRFr group received 0.6 ml of PRFr, while the OVX-ADSCs+PRFr group received 3 × 105 ADSCs suspended in 0.6 ml of PRFr. Intravenous administration can be technically difficult, and the use of a restraining device is often required. It is common practice to stimulate dilation of the tail veins in mice by immersing them in warm (40°C) water for approximately 15 s 39 . Eight weeks after treatments, the animals were sacrificed and were subjected for micro-computed tomography (CT) analysis followed the protocol as described below.
Micro-CT Analysis
The methods used were in accordance with guidelines for the use of micro-CT in rodents 40 . The mice’s left tibia were harvested 8 wk after the first injection, placed in a custom-fabricated jig with water, and scanned with a SkyScan 1176 MicroCT scanner (SkyScan, Kontich, Belgium) operating at 40 kV, 600 μA, 0.3 µm of rotation step, 0.5 mm Al filter, and 9 mm/pixel of scan resolution. The image slices were reconstructed using the NRecon (v.1.4.4, SkyScan) software system. The three-dimensional parameters of bone microarchitecture were calculated using CTAn (v.1.7.0.0, SkyScan) software. For trabecular bone, the proximal tibia was selected for analysis within a conforming volume of interest commencing at the growth plate and extending a further longitudinal distance of 0.5 to 1.5 mm in the proximal direction to assess site-specific responses to OVX and treatment. Microstructural measurements included total volume (TV, volume of region of interest, mm3), bone volume (BV, volume of region segmented as bone, mm3), bone volume fraction (BV/TV, %), BMD (g/cm3), trabecular thickness (Tb.Th, mean trabecular thickness, mm), trabecular number (Tb.N, average number of trabeculae per unit length calculated as 1, mm–1), and trabecular separation (Tb.Sp, distance between trabeculae, mm).
Histological Examination
To determine osteogenesis in the treated mice, tibia specimens of the mice were collected after 8 wk of treatment. Bone samples were fixed for 2 wk with 10% buffered formaldehyde, decalcified in a solution containing 20% sodium citrate and 50% formic acid in 1:1 dilution distilled water for 2 to 3 wk, dehydrated by an alcohol gradient and xylene soak, and placed on Baptist wax and embedded. Sections (5 μm) were cut sagittally along the tibial shaft axis and collected on glass slides before being deparaffinized and subjected to hematoxylin and eosin (H&E) staining using standard protocols. After mounting with coverslips, specimens were viewed and analyzed under a light microscope (Canon Inc., Tokyo, Japan). Relatively well-developed trabecular and cortical bone was observed in the tibia of untreated and sham control mice, and osteoporotic histological profiles were observed in OVX control mice as significant decreases in trabecular and cortical bone masses and increases in connective tissues.
Serum Biochemistry
At the end of the experimental period, the mice were euthanized by carbon dioxide inhalation and blood samples were collected by maximal terminal collection by cardiocentesis with a 1-ml tuberculin syringe with a 5/8-inch 25-gauge needle. Whole blood was clotted at room temperature for 2 h, and serum was separated by centrifugation at 700 × g for 10 min and stored at –80°C until analysis. Total calcium (Ca) and phosphorus (P) levels were quantified using commercial assays in the integrated biochemical analyzer Advia 1800 (Siemens Healthcare Diagnostics, Eschborn, Germany).
Statistical Analysis
All results were presented as mean ± standard derivation. The statistical significance of the tibia and biochemical effects was determined by one-way analysis of variance (ANOVA) using a Statistical Analysis System (2014, SAS/STAT User’s Guide, ver. 13.2. SAS Institute, Inc., Cary, NC, USA), followed by ANOVA with Duncan’s test. Differences with P < 0.05 and <0.01 between the test groups and the control group were considered to be statistically significant 41 .
Results
Analysis of BMD in Tibial Bone Tissue
BMD analysis showed significantly different changes in proximal tibial trabecular BMD in the OVX groups (P < 0.05) (Fig. 1). Eight weeks after the first injection, the OVX-ADSCs (mean BMD, g/cm3, 0.066 ± 0.008), OVX-PRFr (0.074 ± 0.009), and OVX-ADSCs+PRFr (0.077 ± 0.010) groups had a significant increase in tibial shaft BMD compared with the OVX-Ctrl (0.047 ± 0.006) group. In particular, the BMD in OVX-ADSCs+PRFr was higher than in OVX-ADSCs and OVX-PRFr.
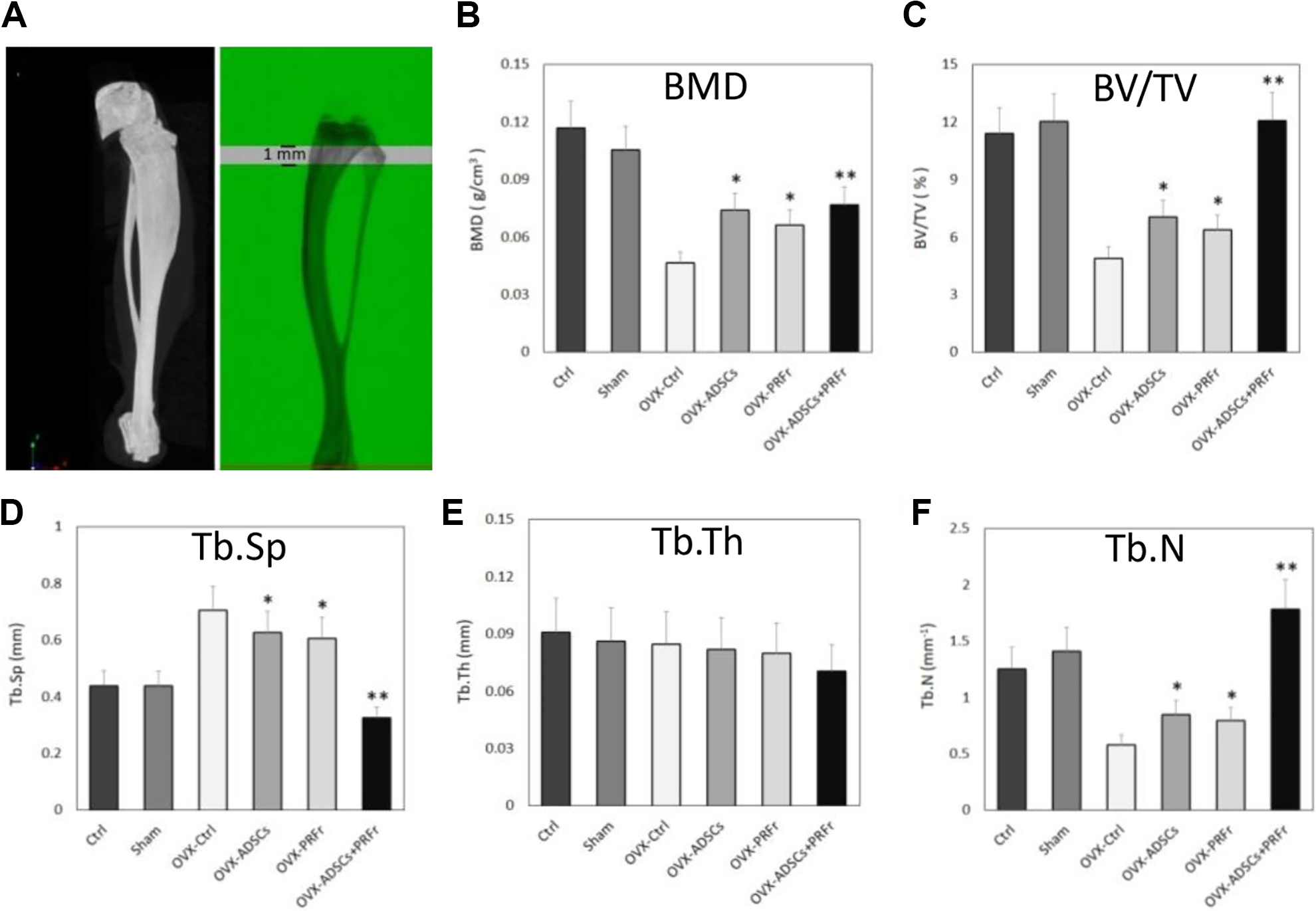
A. Schematic representation of the trabecular bone area using micro-CT analysis. The proximal tibia was selected for analysis within a conforming volume of interest commencing at the growth plate and extending a further longitudinal distance of 0.5 to 1.5 mm in the proximal direction to assess site-specific responses to OVX and treatment. B–F. Micro-CT analysis of Ctrl, Sham, OVX-untreated, and OVX-treated tibia from ICR mice (six groups, n = 6) 8 wk post-transplantation. Illustration of BMD, BV/TV, Tb.Sp, Tb.th, and Tb.N from the entire group. The asterisks indicate significant differences as compared with the OVX-Ctrl group (*P < 0.05; **P < 0.01).
Microstructure Analysis of Tibial Bone
Significant differences in bone mineral quantitative data were confirmed with the micro-CT analysis as shown in Fig. 1. Quantitative analysis of the tibia (Fig. 1) confirmed that the OVX-Ctrl group had the lowest values in BV/TV (4.89 ± 0.58) and morphological parameters such as Tb.N (0.57 ± 0.09). The sham-operated treatment showed minimal effects on the tibial bone mineral as well as in the Ctrl group. When the relative trabecular BV/TV was measured, it was clear that either ADSCs+PRFr-treated mice (P < 0.01) or ADSCs-/PRFr-treated alone (P < 0.05) demonstrated a significantly higher trabecular bone ratio. Otherwise, increased Tb.N and decreased Tb.Sp in OVX-injection groups were observed. Though the BV (P < 0.05) and Tb.N (P < 0.05) in tibial shaft showed a similar trabecular bone formation between OVX-ADSC and OVX-PRFr groups compared with the OVX-Ctrl group, it did show a progressive increase of trabecular bone mass in the ADSC or PRFr injections alone. However, injection with ADSCs+PRFr markedly increased the BV (P < 0.01) and bone parameters (P < 0.01) in the OVX groups except for Tb.Sp, which was significantly decreased (P < 0.01). Additionally, no significant changes in Tb.Th were observed in any of the treatment groups tested. Overall, the ADSCs+PRFr treatment did restore the BV and bone parameters to the normal levels as well as the Ctrl and Sham groups.
Histological Analysis of Tibial Bone
Figure 2 shows the H&E-stained diminished trabecular bone of the OVX mice. Decreased Tb.N and increased Tb.Sp in OVX-ADSCs/OXV-PRFr versus OVX-ADSCs+PRFr were observed. It can be obviously seen that more trabecular bone formation was found in the tibia while trabecular spacing was decreased in the OVX-ADSCs+PRFr group. Similarly, trabecular conjunction points were obviously increased in the OVX groups treated with ADSCs+PRFr when compared to the groups treated with ADSCs or PRFr alone.

Histological sections of osteoporotic mice proximal tibia 8 wk post-transplantation, of either ADSCs, PRFr, or in combination therapy (ADSCs+PRFr), stained using H&E staining methods. The black arrows indicate bony trabeculae. Scale bar—200 μm.
Analysis of Mineral Elements in Serum
The quantitative analysis of Ca and P in serum at 8 wk post-transplantation is shown in Fig. 3. These data showed that the Ca had similar levels in the Ctrl and sham groups, while there were significantly higher concentrations of Ca in the OVX-ADSCs, OVX-PRFr, and OVX-ADSCs+PRFr groups than in the OVX-Ctrl group. Figure 3 also showed similar P contents in all groups regardless of the unoperation/injection for the OVX models, except that the P content in the OVX-ADSCs+PRFr group was significantly higher than in the OVX-Ctrl group (P < 0.05), implying that the ADSCs+PRFr injection might be associated with positive bone parameters.

Total calcium and phosphorous in serum (six groups, n = 6) 8 wk post-transplantation. The asterisks indicate significant differences as compared with the OVX-Ctrl group (*P < 0.05; **P < 0.01).
Discussion
OP is a skeletal disease characterized by decreased bone mass and microarchitectural deterioration of bone tissue, resulting in in an increased risk of fragility fractures. The pathophysiological involution OP of estrogen deficiency has been identified in both the early accelerated and late slow phases of bone loss in women following menopause. The accelerated phase in women is most apparent during the first decade after menopause and involves a disproportionate loss of cancellous bone. It is mediated mainly by the loss of the direct restraining effects of estrogen on bone cell function 42 . OVX mice are classically used as an animal model to simulate postmenopausal bone loss and patterns of bone loss after ovariectomy. In the present study, we intended to explore the feasibility of using ADSCs and PRFr as a cell-based therapeutic approach for treating OP in an OVX mice model by vein injection.
One of the key cellular players in bone physiology is the osteoblast, a bone-forming cell derived from bone marrow-resident mesenchymal stromal cells. Bone marrow-derived mesenchymal stem cells (BMSCs) are self-renewing in vivo. The shift in their differentiation from osteogenic to adipogenic lineage with increase in age can potentially affect osteoblast formation and bone remodeling, and thus be a factor in OP development and pathogenesis 43 . ADSCs have been shown to be pluripotent and explored for their usage in tissue engineering. We focused on ADSCs because of the easy isolation procedure and abundant supply and that fat-derived ADSCs are a more preferred source of autologous mesenchymal cells compared to BMSCs. Moreover, ADSCs proliferate at a higher rate than BMSCs 44 , and have been found to secrete a variety of cytokines and growth factors with trophic paracrine effects that may mediate and stimulate tissue repair and regeneration 45 . In this study, flow cytometry was used to characterize ADSCs isolated from young female ICR mice. ADSCs were distinguished and they showed the highly positive expression of surface markers such as CD73 and CD90 and negative expression of CD34 and CD45.
Previous studies have shown that autologous growth factors released by PRF gradually expressed stronger and more durable effect on proliferation and differentiation of rat osteoblasts than those released by PRP 28 . A variety of tests suggested that PRF was superior to PRP in growth factor release, from the aspects of expression of alkaline phosphatase and induction of mineralization; therefore, the use of PRFr seems to be a more promising method to stimulate cellular activities related to healing responses in a way that is controllable and has a relatively long-term effect 28,46 . Although not yet conclusive, it appears that most research supports a positive role for platelet concentrates in bone regeneration in vivo 47,48 . A study further reported that PRP-treated osteo-progenitor cells underwent osteogenesis and they ameliorated OP in OVX-SAMP8 group model 14 . The stimulation of VEGF and MSCs combined with VEGF has been shown to enhance bone formation and healing. Another study has proven that the cross-talk between VEGF and bone morphogenetic protein (BMP) signaling pathways in MSCs promotes osteoblastic differentiation of MSCs, which aids in bone formation and fracture repair 49 . In assessing the efficacy of PRP and MSCs in the treatment of bone failure of OVX animals, a study using OVX-rat model showed good repair of osteoporotic bone defects (a 1.5-mm diameter tibial defect was made) treated with PRP and BMSCs 50 , and also showed that the expression levels of osteogenesis-specific marker genes including RUNX2 (runt-related transcriptionfactor 2), OSX (osterix), and OPN (osteopontin) were significantly upregulated in the OVX-PRP/BMSCs rats 50 . However, PPARγ2 expression was suppressed particularly in the OVX-PRP/BMSCs group 50 . Similarly, a study demonstrated that xenogenic PRP (xenogeneic profiles of cytokines and growth factors) did not induce elevation of bone resorption 35 ; and the treatment of bone failures in rabbits with secondary OP with allogeneic MSCs and xenogenic PRP could induce greater bone consolidation with mature and immature bone tissue production 35 .
In the present study, by using a well-established osteoporotic mice model, we demonstrated that treatment with allogeneic ADSCs combined with xenogenic PRFr can be used to enhance bone formation. To our knowledge, this is the first experimental study of this kind to examine the efficacy of a combination of ADSCs and PRFr in OP treatment. Our results show a significant loss of BMD in the proximal tibia (40%) in mice 16 wk post bilateral OVX, as measured by micro-CT of the proximal tibia. Ultimately, ADSCs+PRFr transplants were shown to be effective in restoring tibial bone density, 8 wk post-transplantation. OVX mice that received ADSCs, PRFr, and ADSCs+PRFr transplants showed significantly higher bone regeneration (an average of 58%, 42%, and 64% of improved BMD, respectively, P < 0.05) over the OVX-Ctrl group. Quantification analysis also demonstrated that Tb.N clearly show significant losses (68%) in mice 16 wk after ovariectomy. The OVX group treated with ADSCs (27%; P < 0.05%), PRFr (22%; P < 0.05%), and ADSCs+PRFr (120%; P < 0.01%) observed higher bone restoration when compared to OVX-Ctrl at 8 wk post-transplantation. During the late phase of bone healing, the newly formed bone in the proximal tibia was remodeled in the sham and the OVX-ADSCs+PRFr-treated mice, whereas remodeling was incomplete in the OVX-ADSCs, OVX-PRFr, and the OVX groups’ mice, as observed in the micro-CT images. Notably, the significantly increased levels of BV and Tb.N of proximal tibia growth plates were observed in the OVX-ADSCs+PRFr (P < 0.01), OVX-ADSCs (P < 0.05), and OVX-PRFr (P < 0.05) groups’ mice as compared with that in the OVX mice, indicating that the positive effect of ADSCs on bone formation might be enhanced in the presence of PRFr. As concentrated growth factors remarkably promoted the growth and osteogenic differentiation capacities of ADSCs in vitro 51 , it is thus possible that PRFr may accelerate bone formation during skeletal repair by upregulating the proliferation and osteogenesis of ADSCs, which is consistent with that of an earlier report 50 .
PRFr is rich in b-FGF, BMPs, EGF, PDGF, TGF, VEGF, insulin-like growth factor (IGF), nerve growth factor (NGF), and so on 27,52 . During the past few years, progress has been made toward the understanding of local regulation of bone remodeling especially in relation to OP. Cytokines have shown to be powerful regulators of bone resorption and formation, though under superior control from estrogen/testosterone, parathyroidhormone, and 1alpha,25(OH)2D3 53 . Collectively, BMPs are useful for improving fracture healing by both increasing osteogenesis and creating a favorable healing environment by altering cytokine release by endogenous cells 54,55 . To this end, the production of b-FGF as a result of BMP-2 exposure during the early phases of healing promotes the induction of BMSCs to become osteoblasts while improving angiogenesis and mitogenesis simultaneously 55 . EGF protracts extracellular signal-regulated kinase and Akt phosphorylation and acts as an important mediator for osteogenic factor 56 . IGF-1 plays an important role in cell proliferation, differentiation, and apoptosis and is the major mediator of the effect of growth hormone on both bone growth and mineralization 57 , via decreasing osteoblast apoptosis and promoting osteoblastogenesis through the phosphoinositide 3-kinase 58 . NGF promotes cartilage differentiation and increases osteoclast formation and healing of mice tibial fractures by increasing the levels of COL2A1 and SOX9 mRNA expression, and may be used for the prevention and treatment of OP 59 . Some of the cytokines primarily enhance osteoclastic bone resorption, e.g., interleukin-1 (IL-1), interleukin-6 (IL-6), and tumor necrosis factor, while others, e.g., IGF, PDGF, and TGF-β, primarily stimulate bone formation 53 . Moreover, VEGF plays the role of a key upregulator of angiogenesis that is essential for bone development and regeneration, especially for large bone defects 60 . Therefore, PRFr may regulate and mediate the balance between osteogenesis and adipogenesis in vivo, which may also contribute to the positive effects of PRFr on bone formation in osteoporotic mice.
Our results suggest that ADSCs and PRFr may be useful in enhancing BV and treating pathological conditions of bone loss. Although mice are usually utilized for the purpose of initial evaluation and biological compatibility tests, these tests only provide limited information. Larger subjects and longer periods may provide more definitive and meaningful support for using this therapeutic approach. In conclusion, the present results indicated that ADSCs and PRFr promote new bone formation. Better results were obtained by combining the two into a single treatment for osteoporotic bone defects. Therefore, the combination therapy of ADSCs and PRFr may be a promising novel therapeutic strategy in osteoporotic bone regeneration.
Footnotes
Author Notes
Shinn-Zong Lin and Tzong-Fu Kuo contributed equally to this work.
Ethical Approval
Ethical approval to report this study was obtained from the Institutional Animal Care and Use Committees (IACUC) of National Taiwan University (NTU-102-EL-82).
Statement of Animal Rights
All experiments were carried out in accordance with the International Council for Laboratory Animal Science guidelines for the care and use of laboratory animals. This research was carried out in strict accordance with an approved IACUC protocol at National Taiwan University (NTU-102-EL-82).
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partly financed by a grant from the Gwo Xi Stem Cell Applied Technology Co., Ltd, Hsinchu, Taiwan.