
Abstract
Bronchopulmonary dysplasia (BPD), also known as neonatal chronic lung disease, is an important cause of respiratory illness in preterm newborns that results in significant morbidity and mortality. Long noncoding RNAs (lncRNAs) have been discovered with many biological functions. However, the role of lncRNAs in the pathogenesis of BPD remains poorly understood. Here, we established a mouse lung injury model that mimicked human BPD. Subsequently, we found the lncRNA H19 expression level was significantly increased in BPD compared with normal lung tissues using quantitative real-time polymerase chain reaction. Next, we observed that overexpression of lncRNA H19 enhanced mitogen-activated protein kinase (MAPK) signaling pathway. In addition, we also found that dysfunction of lncRNA H19 altered the expression of inflammatory factors. Thus, our study validates that lncRNA H19 contributes to the progression of BPD by regulating MAPK signaling pathway, which could be used as a potential target for treating BPD.
Keywords
Introduction
Bronchopulmonary dysplasia (BPD), a chronic lung disorder of preterm birth, is triggered by the disturbances in physiologic lung development 1 . BPD is predominantly characterized by enlarged alveoli, arrested lung growth, abnormal pulmonary function, and so on 2 . It is an important cause of respiratory illness in preterm newborns that results in significant morbidity and mortality. Little is known about the molecular mechanisms underlying BPD; however, among the emerging candidate regulators of BPD are lncRNAs.
The vast majority of the genome is actively transcribed into noncoding RNAs (ncRNAs) under different conditions. Long noncoding RNAs (lncRNAs) are a type of ncRNAs with >200 nucleotides in length and do not encode for proteins 3,4 . LncRNAs have been implicated in many human diseases and regarded as important regulators of many biological activities and disease progression 5,6 . For instance, the lncRNA UCA1 has been verified to promote proliferation, migration, and immune escape and inhibit apoptosis in gastric cancer by sponging antitumor miRNAs 7 . LncRNA HOTAIR has been shown to enhance estrogen receptor (ER) signaling and confers tamoxifen resistance in breast cancer 8 . LncRNA UCA1 exerted oncogenic functions in non-small-cell lung cancer by targeting miR-193a-3p 9 . However, the functional roles of lncRNAs involved in the BPD progression still remain to be clarified. Based on our preliminary lncRNA Array data, lncRNA H19 is one of the upregulated lncRNAs in the lung tissues of neonatal mice under hyperoxia exposure than in neonatal mice under room air (RA) exposure.
Mitogen-activated protein kinase (MAPK) pathway is known to activate a wide range of transcription factors involved in innate immunity, cell growth, stress response, apoptosis, and differentiation 10 . It has been reported that inhibition of the p38-MAPK signaling pathway suppresses the apoptosis and expression of proinflammatory cytokines in human osteoarthritis chondrocytes 11 . It has also been shown that lipoxin A4 reduces inflammation through formyl peptide receptor 2/p38-MAPK signaling pathway 12 . Recently, accumulated studies show that lncRNAs regulate biological functions through altering MAPK signaling pathway in various types of diseases. For instance, lncRNA BANCR promotes endometrial cancer cell proliferation and invasion by regulating MMP2 and MMP1 via extracellular signal-regulated kinase (ERK)/MAPK signaling pathway 13 . However, the effect and possible mechanism of lncRNAs and MAPK in BPD remained unclear.
In this paper, we aimed to detect the expression of lncRNA H19 in BPD, as well as to recover the underlying molecular mechanisms. We first detected the expression of lncRNA H19 in the lung tissue of mice with BPD. After that, lncRNA H19 was overexpressed and knocked down in A549 cells. MAPK signaling response and inflammatory factors were also determined.
Materials and Methods
Reagent
Epidermal growth factor (EGF) and phorbol 12-myristate 13-acetate (PMA) were purchased from Sigma-Aldrich (Shanghai, China).
Exposure of Neonatal Mouse Pups to Hyperoxia
All animal procedures were performed according to the protocol approved by the Ethics Committee of the Foshan Chancheng Central Hospital, Guangdong, China. Kunming mice were purchased from the Chinese Academy of Sciences (Beijing, China), and were bred in-house based on previously described model 14 . Briefly, the neonatal mice, delivered on the same day, were mixed and divided into two groups (n = 15 each group). One group of pups was placed in a sealed Plexiglas chamber (22°C –27°C, humidity between 50% and 70%) and exposed to 95% O2 from postnatal day 1 to 4, then allowed to recover in RA for the following 10 d. The other group was kept in RA. The hyperoxia treatment was maintained by continuous oxygen (4 L/min) and monitored with an oxygen sensor (VacuMed, Ventura, CA, USA).
Tissue Collection
The neonatal pups (n = 15 each group) were euthanized on postnatal day 14, trachea cannulated, and had their lungs fixed with 4% paraformaldehyde by instilling endotracheally at a 30 cm H2O pressure 15 . For RNA and protein samples, the lungs were excised and frozen in liquid nitrogen immediately after mincing (cutting into small pieces) on ice and were then stored at −80°C until subsequent analysis.
Morphological Assessment
The morphology of mice lungs was determined by the mean linear intercept (MLI), which was performed as a measure of interalveolar wall. The MLI was obtained by dividing the total length of a line drawn across the lung section by the total number of intercepts encountered in 50 lines per mouse lung.
Cell Line
A549 cells were purchased from the China Center for Type Culture Collection (CCTCC, Wuhan, Hubei, China). The cells were cultured in 1640 (Invitrogen, Shanghai, China), with 10% fetal bovine serum (Invitrogen), 100 U/ml penicillin, and 100 mg/ml streptomycin (Sigma-Aldrich) at 37°C in a humidified atmosphere with 5% CO2.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
The total RNA from tissues and cells was extracted using the mirVanaTM miRNA isolation kit (Ambion, Austin, TX, USA) according to the manufacturer’s protocol. After RNA isolation, RNA samples were reverse transcribed to cDNA using the high-capacity RNA-to-cDNA kit (Thermo Fisher Scientific, Shanghai, China). For H19, tumor necrosis factor α (TNF-α), interleukin 1β (IL-1β), IL-6 expression analysis, the quantification of cDNA was analyzed using SYBR Green Master Mix (Applied Biosystems, Beijing, China) with glyceraldehyde 3-phosphate dehydrogenase as a housekeeping gene for normalization. The thermal cycling conditions are as follows: 95°C for 4 min, followed by 35 cycles consisting of 95°C for 20 s, 61°C for 15 s, 72°C for 30 s, and the final cycle expanded 72°C for 5 min. All the primers are listed in Table 1.
qRT-PCR Primers for mRNA.

GAPDH: glyceraldehyde 3-phosphate dehydrogenase; IL-1β: interleukin 1β; qRT-PCR: quantitative real-time polymerase chain reaction; TNF-α: tumor necrosis factor α.
Overexpression and Silencing
PcDNA3.1-H19, pcDNA3.1 vector control (H19-Con), H19 targeting siRNA (si-H19), and siRNA negative control vector (si-NC) were commercially available at Genepharma Inc. (Shanghai, China). Cell transfection was performed by Lipofectamine RNAiMAX (Thermo Fisher Scientific) according to the manufacturer’s protocol.
Luciferase Reporter Assay
For the pathway activity, A549 cells were transfected with PcDNA3.1-H19 or H19 targeting siRNA (si-H19) or their respective negative control vector (H19-con or si-NC) with ERK-MAPK or c-Jun N-terminal kinase (JNK)-MAPK pathway-luciferase-reporter construct (OBiO Technology, Shanghai, China). EGF (10 ng/well) or PMA (1 ng/well) was added after 24-h transfection. After 18-h stimulation, the cells were harvested. ERK-MAPK or JNK-MAPK transcriptional activities were measured by Dual Luciferase Reporter Assay System (Promega, Madison, WI, USA). Data were shown as a ratio of firefly luciferase activity to Renilla luciferase activity.
Statistical Analysis
All statistical analyses were performed by using the SPSS 22.0 software (IBM, Armonk, NY, USA). Student’s unpaired t-test (comparison of two groups) or analysis of variance followed by Student–Newman–Keuls (comparison of more than two groups) was used for the difference analysis. All data were expressed as mean ± standard error (SE). P <0.05 indicated statistical significance.
Results
Successful Establishment of BPD Mouse Model
BPD mouse model was established via exposing neonatal mice to hyperoxia. We first validated the successful establishment of BPD mouse model. In Fig. 1A, we showed that the body weight of neonatal mice began to significantly decrease in the third day after hyperoxia exposure compared with those exposed to RA. We also confirmed the BPD model by hematoxylin-eosin staining. Histological analysis showed airway enlargement in BPD mice group, which was reflected by a significant increase in the MLIs (Fig. 1B, C). The lung tissues of hyperoxia-treated neonatal mice showed disordered alveolar structure, as is evident by the higher MLI values in neonatal mice compared with those after hyperoxia exposure.
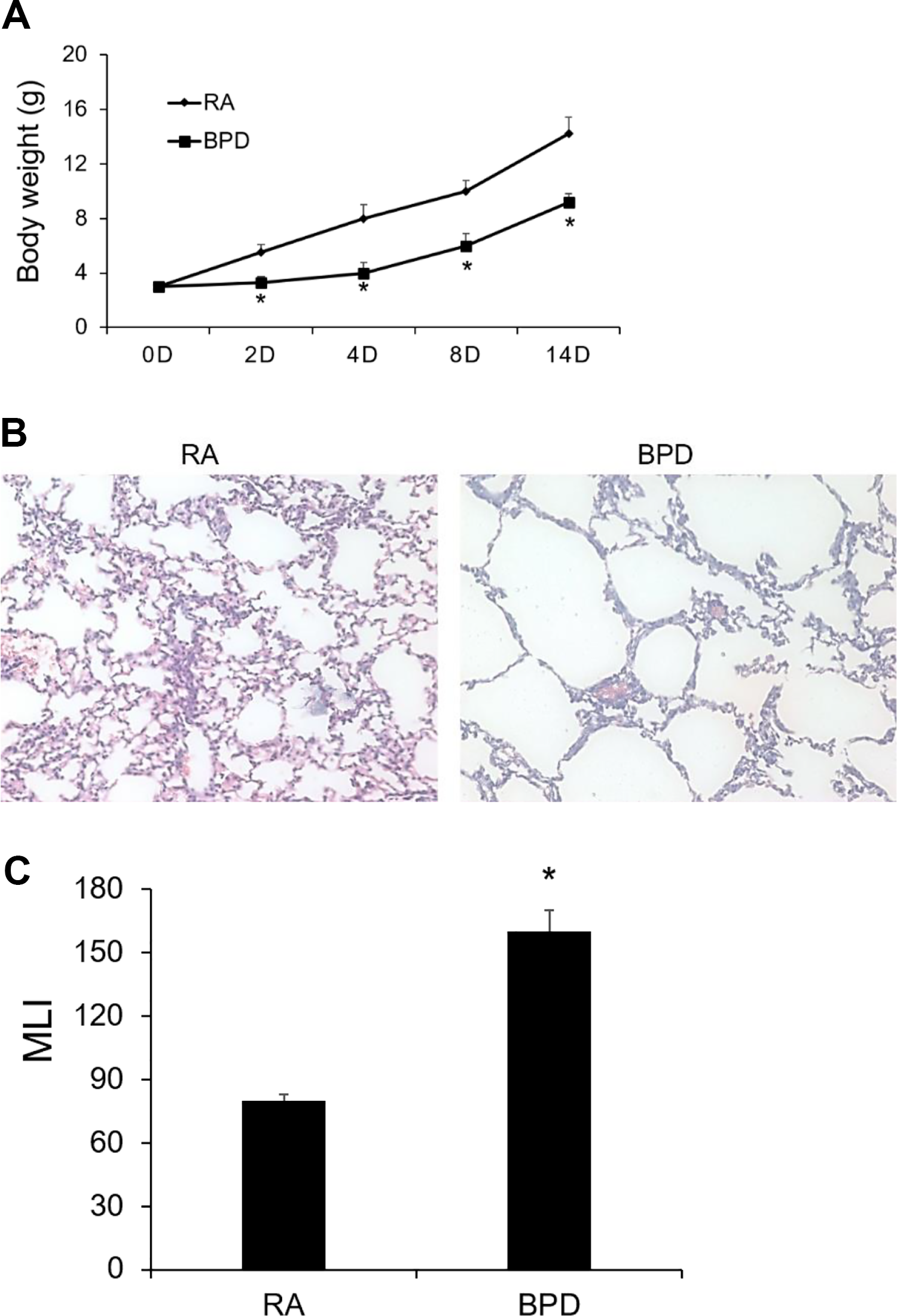
Hyperoxia-induced BPD mouse models are successfully established. (A) Body weight of neonatal mice under exposure of RA or hyperoxia-induced BPD; (B) Hematoxylin-eosin staining of lung tissues of neonatal mice under exposure of RA or hyperoxia-induced BPD (20×); (C) MLI. Results shown are means ± standard error. *P < 0.05 vs. RA.BPD: bronchopulmonary dysplasia; D: days; RA: room air; MLI: mean linear intercepts.
LncRNA H19 Was Upregulated in Hyperoxia-Induced BPD
To explore for important lncRNAs involved in the progression of BPD, the lncRNAs expression of BPD mice and mice explored to RA were detected using the lncRNA array. Based on our preliminary lncRNA array data, lncRNA H19 is one of the upregulated lncRNAs in the lung tissues of neonatal mice under hyperoxia exposure than in neonatal mice under RA exposure. To validate the expression of lncRNA H19 in BPD, qRT-PCR was performed to determine lncRNA H19 expression in the lung tissues of neonatal mice. The expression of lncRNA H19 was significantly increased in lung tissues of neonatal mice under hyperoxia exposure than in neonatal mice under RA exposure (Fig. 2).

LncRNA H19 was highly expressed in the lung tissues with BPD. LncRNA H19 was detected in the lung tissues with BPD using quantitative real-time polymerase chain reaction, compared with those from RA. Results shown are means ± standard error. *P < 0.05 vs. RA.BPD: bronchopulmonary dysplasia; RA: room air.
Overexpression of LncRNA H19 Enhanced MAPK Signaling Pathway
To decipher the underlying molecular mechanism and the effects of lncRNA H19, we subsequently overexpressed and knocked down of lncRNA H19 in A549 cells. The overexpression and silencing of lncRNA H19 was verified by qRT-PCR (Fig. 3A, B). LncRNA H19 overexpression is known to activate MAPK signaling pathway in a variety of cancers 16 . Therefore, we investigated whether alternation of lncRNA H19 can regulate MAPK signaling pathway. As depicted in Fig. 3C, lncRNA H19 overexpression significantly upregulated the activity of both ERK and JNK luciferase reporters after stimulation compared to the control vector, H19-Con. However, lncRNA H19 silencing remarkably downregulated the activity of both ERK and JNK luciferase reporters after stimulation compared to the si-NC (Fig. 3D). These results suggest that lncRNA H19 contributed to the progression of BPD via upregulating both ERK and JNK signaling pathways.

Effects of lncRNA H19 on ERK and JNK signaling pathways. (A, B) A549 cells were transfected with H19-OE, H19-Con, si-H19 or si-NC. The lncRNA H19 expression was detected using quantitative real-time polymerase chain reaction. Results shown are means ± SE. *P < 0.05 vs. control. (C) A549 cells were cotransfected with ERK or JNK luciferase-reporter construct and H19-OE or H19-Con. The cells were stimulated with EGF or PMA. The firefly luciferase activity was normalized to Renilla luciferase activity. (D) A549 cells were cotransfected with ERK or JNK luciferase-reporter construct and si-H19 or si-NC. The cells were stimulated with EGF or PMA. The firefly luciferase activity was normalized to Renilla luciferase activity. Results are expressed as fold change over control. *P < 0.05 vs. control, ^P < 0.05 vs. stimulation. Error bars represent SE. EGF: epidermal growth factor; ERK: extracellular signal-regulated kinase; H19-Con: pcDNA3.1 vector control; H19-OE: PcDNA3.1-H19; JNK: c-Jun N-terminal kinase; PMA: phorbol 12-myristate 13-acetate; SE: standard error; si-H19: H19 targeting siRNA; si-NC: siRNA negative control vector.
LncRNA H19 Promoted Lung Inflammation
MAPK signaling pathways are important signal transducers mediating inflammation. We next determined the levels of the inflammatory cytokines in the lung tissues of neonatal mice under hyperoxia exposure than in those under RA exposure. The mRNA levels of TNF-α, IL-1β, and IL-6 were significantly higher in the lung under hyperoxia than RA (Fig. 4A). To elucidate the effects of lncRNA H19 expression alternation on BPD progression, we subsequently determined the mRNA levels of TNF-α, IL-1β, and IL-6 in A549 cells after lncRNA H19 overexpression or knockdown. As we expected, the mRNA levels of TNF-α, IL-1β, and IL-6 were strongly increased after lncRNA H19 overexpression (Fig. 4B). In contrast, the mRNA levels of TNF-α, IL-1β, and IL-6 were strongly decreased in A549 cells with lncRNA H19 silencing (Fig. 4C). These data support that lncRNA H19 contributes to the progression of BPD during the inflammatory response.

Effect of lncRNA H19 on lung inflammation. (A) mRNA levels of TNF-α, IL-1β, and IL-6 in the (A) lung tissues with BPD using qRT-PCR, compared with those from RA. Results shown are means ± SE. *P < 0.05 vs. RA (B) in the A549 cells transfected with H19-OE or H19-Con using qRT-PCR. Results shown are means ± SE. *P < 0.05 vs. H19-Con (C) in the A549 cells transfected with si-H19 or si-NC using qRT-PCR. Results shown are means ± SE. *P < 0.05 vs. si-NC. BPD: bronchopulmonary dysplasia; H19-Con: pcDNA3.1 vector control; H19-OE: PcDNA3.1-H19; IL-1β: interleukin 1β; qRT-PCR: quantitative real-time polymerase chain reaction; RA: room air; SE: standard error; si-H19: H19 targeting siRNA; si-NC: siRNA negative control vector; TNF: tumor necrosis factor.
Discussion
Among the complications associated with mechanical ventilation in preterm infants, BPD is a serious disorder considering the lack of effective treatment options 17 . With the rapid development of perinatal medicine, the survival rate of premature infants of very low-birth-weight infants and extremely low-birth-weight infants has remarkably improved 18 . There is compelling evidence highlighting the regulatory role of lncRNAs in the pathogenesis of BPD by targeting signaling pathways and specific genes. Therefore, understanding the molecular mechanisms in BPD might offer novel strategies for preventing or arresting progression of BPD.
We initially established the BPD mice model and found that lncRNA H19 expression was significantly increased in lung tissues of neonatal mice with hyperoxia-induced BPD. LncRNA H19 has been found to promote the migration and invasion of colon cancer cells via MAPK signaling pathway 19 . LncRNA H19 has also been shown to accelerate tenogenic differentiation and promotes tendon healing through targeting miR-29b-3p and activating TGF-β1 signaling 20 . Despite its emerging and complicated biological functions, lncRNA H19 exerts its functions mainly through producing miRNAs as its primary precursor or interacting with diversified miRNAs 21 . From the results specified earlier, it is indicated that lncRNA H19 might be involved in the progression of BPD.
MAPK pathway is known to play an essential role in the assembly and disassembly of multiprotein complexes and determines cell fate. The signaling mediators of MAPK include ERKs, JNKs, and p38 MAPK 22 . Notably the activation of the MAPK signaling is important in the response of inflammation 23 . LncRNAs have been reported to regulate biological functions via altering MAPK signaling pathway 20 . LncRNA AOC4P has been reported to affect biological behavior of gastric cancer cells through MAPK signaling pathway 24 . LncRNA NEAT1 has also been identified as a novel inflammatory regulator acting through MAPK pathway in human lupus 25 . In spite of all these studies, there are no studies on lncRNA H19 as it relates to BPD progression. In this study, we identified lncRNA H19 as a novel inflammatory regulator acting through MAPK signaling pathway in BPD.
In summary, lncRNA H19 was shown to be upregulated in lung tissues of BPD mice model. In addition, lncRNA H19 has also been identified to regulate inflammatory responses via MAPK signaling pathway. However, it is worthy to consider the impeded effect change when cotransfected with ERK or JNK luciferase-reporter construct and H19-OE or H19-Con. It is also needed to consider that hyperoxia may induce epigenetic changes in newborn mice lungs. To the best of our knowledge, our study is the first to reveal lncRNA H19 and its regulatory function with MAPK signaling pathway in BPD, which provides a foundation for further understanding of the molecular mechanism of BPD development.
Footnotes
Authors’ Contributions
JZ conceived the study. WM and JZ designed the experiments and wrote the manuscript. WM and YL collected the samples and completed the experimental part of the study. YL, WC, and BM analyzed the data and revised the manuscript.
Acknowledgments
The authors would like to thank the AceSci Biomed Inc. (Wuhan, Hubei, China) for helping in English proofreading.
Ethical Approval
Ethical approval (IRB-ATT-001-24) was obtained for all experimental procedures by the Ethics Committee of the Foshan Chancheng Central Hospital, Guangdong, China.
Statement of Human and Animal Rights
All procedures in this study were performed according to the protocol approved by the Ethics Committee of the Foshan Chancheng Central Hospital, Guangdong, China.
Statement of Informed Consent
Verbal informed consent was obtained from the patients involved for their anonymized information to be published in this article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The research was supported by grants from the project of the National Science Fund of Distinguished Young Scholars in Foshan, Guangdong.