
Abstract
There is still a lack of sufficient research on the mechanism behind neurogenic bladder (NB) treatment. The aim of this study was to explore the effect of overexpressed stromal cell-derived factor-1 (SDF-1) secreted by engineered immortalized mesenchymal stem cells (imMSCs) on the NB. In this study, primary bone marrow mesenchymal stem cells (BM-MSCs) were transfected into immortalized upregulated SDF-1-engineered BM-MSCs (imMSCs/eSDF-1
Introduction
Neurogenic bladder (NB) is a vexing problem for both the patients and the urologists 1 . As is well known, NB is not a homogeneous entity, but a widely used term to denote lower urinary tract dysfunction, which is a common complication of neurological disease following surgical treatment of pelvic neoplasms 2 . The symptoms involve frequent urination, urinary incontinence, voiding by abdominal staining, and urinary retention 3 . Currently, the primary goals of NB management include protection of the upper urinary tract function and restoration of bladder dysfunction 4 . However, the mechanism underlying NB treatment is still not fully understood.
Mesenchymal stem cells (MSCs) represent a novel and potential therapeutic strategy with convincing and promising evidence in many medical fields 5 –7 . Currently, stem cell therapy has evolved from animal experiments to clinical settings 8 , which indicates their safety and effectiveness. In our previous study 9 , we found that MSC therapy accelerated the recovery of injured nerve and tissue. Therefore, we believe that MSC therapy is effective in NB treatment. Immortalized stem cells were selected for experimental use for increased accuracy 10,11 , because immortalized mesenchymal stem cells (imMSCs) exhibit higher proliferation and anti-senescence ability, compared with general MSCs.
In a recent study, researchers found that stromal cell-derived factor-1 (SDF-1) induced stem cells to recruit endothelial cells, which contributed to vascular pericytes 12 . In our previous experiment, we found that SDF-1 was active during the whole period of tissue recovery and angiogenesis 9 . Therefore, we wondered whether NB can be ameliorated effectively under a high SDF-1 microenvironment. We utilized genetic engineering technology to overexpress SDF-1 to treat the NB in a rat model.
In this study, we developed a rat model of NB via bilateral pelvic nerve (PN) transection, and analyzed the effect of high SDF-1 on NB via upregulation of SDF-1-engineered imMSCs. We hypothesized that upregulated SDF-1 in imMSCs might improve NB function and restore bladder wall efficiently by promoting angiogenesis, tissue regeneration, and nerve recovery.
Materials and Methods
Cell Culture and Preparation
Primary bone marrow mesenchymal stem cells (BM-MSCs) were obtained from Catholic Institute of Cell Therapy (CIC, Seoul, Korea), were cultured in low glucose-containing Dulbecco’s modified Eagle’s medium (Gibco, Waltham, MA, USA) supplemented with 20% fetal bovine serum (Gibco, Waltham, MA, Gibco) and 5 ng/ml basic fibroblast growth factor (bFGF, Cell Signaling Technology, Danvers, MA, USA) at 37°C and 5% CO2. To generate upregulated SDF-1-engineered imMSCs, c-myc, hTERT, tetracycline transactivator, and SDF-1 genes were synthesized and transfected with pBD lentiviral vector (SL-BIGEN, Seongnam, Korea). In brief, the reference sequence of SDF-1 was NM_000609
6
. We designed an optimized DNA sequence to introduce into a vector (Genscript, Nanjing, China). DNA sequencing was performed to confirm the accuracy of the right reading frame of the introduced sequence (Cosmo Genetech, Seoul, Korea). After transfection, cells were seeded on 12-well plates, at a density of 5 × 105 cells per well. After 48 h, the supernatant was harvested. The SDF-1 protein level was measured. The SDF-1 expression level was about 10 ng/ml. Immortalized, upregulated SDF-1-engineered BM-MSCs (imMSCs/eSDF-1
ELISA
BM-MSCs, imMSCs/eSDF-1
Experimental Animal and Study Design
All animal experiments in this study were approved by the Institutional Animal Care and Use Committee of the Catholic University of Korea (CUMC-2016-0218-01). The 12-week-old male Sprague-Dawley rats, each weighing about 370 to 400 g, were purchased from Orient Bio Co (Orient Bio Co., Seongnam, Korea), and separated into three experimental groups (NB, NB+imMSCs/eSDF-1
MSC Treatment
One week after the NB model was established, rats in the NB+imMSCs/eSDF-1
Voiding Pattern Analysis
For voiding pattern analysis, rats were individually housed in metabolic cages with a grid at the bottom, and a filter paper below the grid to collect the urine for 3 h. Filter papers were photographed under ultraviolet light and analyzed using the Image J software (NIH, Bethesda, MD, USA) to identify the surface area of the individual voiding spots. The number of spots was defined as the voiding frequency, and 50 µl urine was considered equal to 2.8 cm2 on the filter paper. Voiding pattern analysis was conducted before and after treatment.
Cystometry
Cystometry experiments were performed in all rats at week 4 after the treatment. In brief, rats were anesthetized using a subcutaneous injection of 1.2 mg/kg of urethane. A suprapubic midline laparotomy was performed to expose the bladder, and a 25-gauge needle connected to a polyethylene tubing was inserted into the bladder through the bladder dome. The tubing was connected to a pressure transducer and a Harvard syringe pump via a three-way stopcock to record intravesical pressure and to infuse saline into the bladder. After emptying the bladder, cystometry was performed using a saline infusion at a rate of 0.04 ml/min. The contraction interval and the contraction pressure (maximum bladder pressure during voiding) were recorded using a polygraph (Grass 7D; Grass Institute Co., Quincy, MA, USA). Nonvoiding contractions (NVCs) were determined during the 2 to 4 min before each voiding contraction. NVCs were defined as contractions >4 cm H2O from the baseline pressure during bladder filling. After cystometry, PN and bladder were collected for immunofluorescence staining and western blot analysis.
Histology and Immunofluorescence Staining
The collected PN and bladder were fixed in 4% paraformaldehyde for 24 h at 4°C before creating a paraffin block, as described previously 9 . The primary antibodies were used as follows: vascular endothelial growth factor (VEGF; diluted 1:200; Santa Cruz Biotechnologies, Santa Cruz, CA, USA), βIII-tubulin (diluted 1:200; Abcam, Cambridge, UK), bFGF (diluted 1:500; Cell Signaling Technology, Danvers, MA, USA), SDF-1 (diluted 1:200; Abcam, Cambridge, UK), poly-ADP-ribose polymerase (PARP, diluted 1:500; Abcam, Cambridge, UK), and 6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Inc., Burlingame, CA, USA) were used to stain the nuclei. Digital images were obtained using a Zeiss LSM 510 Meta confocal microscope (Zeiss, Oberkochen, Germany), and bladder images were analyzed using ZEN 2012 (Zeiss, Oberkochen, Germany). The other resulting images were analyzed using Image J to determine the positive rate for each figure. In brief, we used Image J to split each color from merged figure, calculate the intensity of each color, and then added them. The positive was the ratio of a special color fluorescence intensity to all colors intensity.
Masson Staining
To evaluate the proportion of collagen and smooth muscle, the bladder walls of rats in each group were stained with Masson trichrome. Bladder wall sections were deparaffinized, rehydrated with graded alcohols, immersed in warm Bouin’s solution (Sigma, St. Louis, MO, USA) (55 to 60°C) for 2 h and washed under running tap water for 2 min followed by distilled water for 30 to 60 s. They were stained with Weigert Hematoxylin (Merck, Darmstadt, Germany) for 10 min and washed out. Subsequently, the smooth muscle was stained red with Biebrich Scarlet-Acid-Fuchsin (Sigma, St. Louis, MO, USA) for 10 min, rinsed and immersed in phosphomolybdic phosphotungstic acid (Sigma, St. Louis, MO, USA) for 15 min. The collagen was stained blue with Aniline Blue (Sigma, St. Louis, MO, USA) for 10 min and rinsed in distilled water followed by immersion in 1% acetic acid for 5 min. Finally, the tissues were rehydrated with 100% ethanol, left to air dry and mounted. All sections of bladder tissue were stained at the same time.
Analysis of Apoptosis In Vivo
We assessed the apoptosis in PN using an immunostaining kit to determine the PARP levels as described previously 9 . The antibodies to PARP along with other primary antibodies are listed in the Histology and Immunofluorescence Staining section.
Western Blot
Western blot was performed as previously described 9 . In brief, the collected tissue was homogenized using ice-cold RIPA buffer (Cell Signaling Technology, Danvers, MA, USA) containing EDTA-free protease inhibitor cocktail. The phosphatase inhibitor cocktail (Roche Diagnostics GmbH, Basel, Switzerland) and particulate mass were removed by centrifugation (15,000g) for 15 min at 4°C. The supernatants were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The primary antibodies included neuronal nitric oxide synthase (nNOS, diluted 1:200; Santa Cruz Biotechnologies), AKT (diluted 1:200; Cell Signaling Technology, Danvers, MA, USA), p-AKT (diluted 1:200; Cell Signaling Technology, Danvers, MA, USA), p-ERK (diluted 1:200; Cell Signaling Technology, Danvers, MA, USA), SDF-1(diluted 1:500; Cell Signaling Technology, Danvers, MA, USA), and β-actin (diluted 1:1,000; Abcam, Cambridge, UK).
Statistical Analysis
All data are presented as mean ± standard error and analyzed using SPSS version 22.0 software (SPSS Inc., Chicago, IL, USA). Student’s t-test and one-way analysis of variance (ANOVA) or two-way ANOVA (followed by Bonferroni post hoc tests) as appropriate were used to evaluate the significant differences among groups. P < 0.05 was considered statistically significant.
Results
imMSCs/eSDF-1
+
Express Higher SDF-1 In Vitro and Vivo
As shown in Figure 1(A, a) imMSCs/eSDF-1
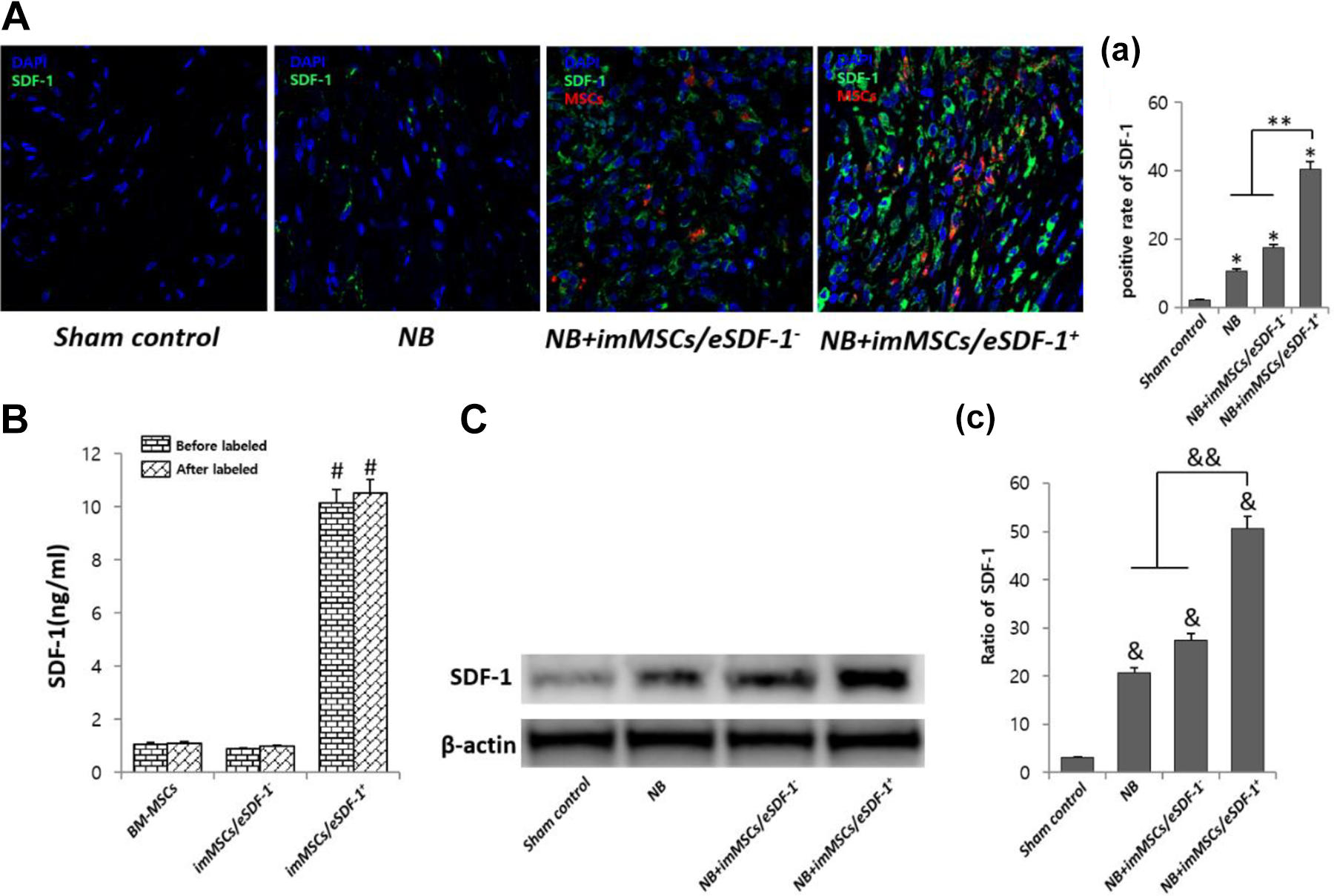
ImMSCs/eSDF-1
imMSCs/eSDF-1
+
Improves NB Significantly
The result of cystometry (Figure 2A) in NB group was apparently better than other groups. As shown in Figure 2(B and C), the mean pressure of the voiding contractions and intermicturition interval in each group varied. The bladders of NB+imMSCs/eSDF-1
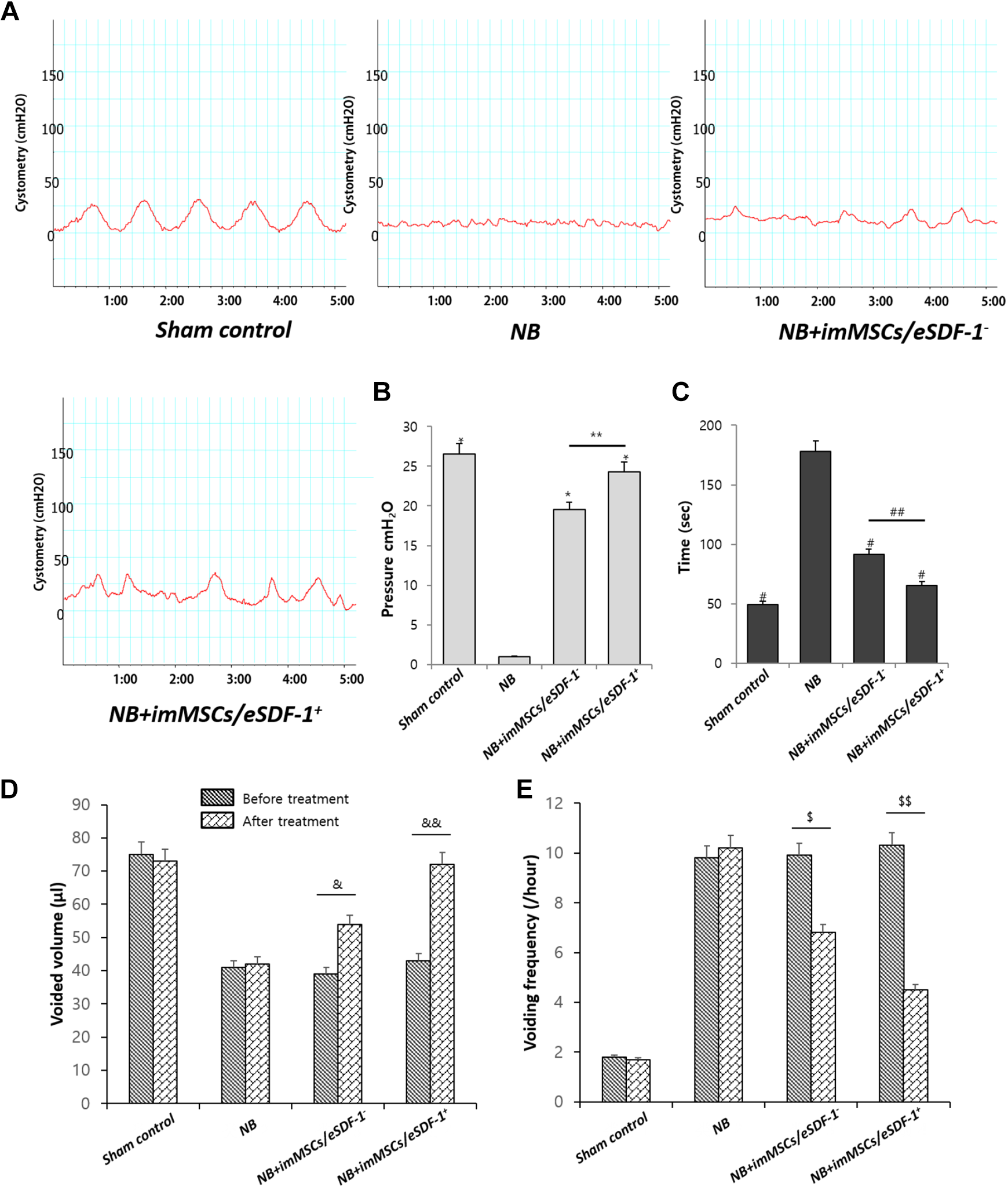
(A) Representative images of cystometry in each group. Sham control is the normal control group. NB means neurogenic bladder. ImMSCs/eSDF-1
MSCs Treatment Stimulates the Bladder Recovery Especially in a High SDF-1 Microenvironment
During the 4-week treatment, the bladder wall was assessed by Masson staining. As shown in Figure 3(A and B), the smooth muscle of bladder wall was thicker after MSC treatment (P < 0.01). The smooth muscle in the NB+imMSCs/eSDF-1

(A) Representative images of Masson Trichrome stain for each group. Red is smooth muscle, and blue is collagen. Original magnification: ×200. (B) Percentage area of smooth muscle for each group. Each bar shows the mean values (standard deviation). *P < 0.01 compared with NB group. **P < 0.05 compared with NB+imMSCs/eSDF-1
SDF-1 Induces MSC Homing and Subsequently Improves the Regeneration of Injured PN
As shown in Figure 4(A), nerve recovery was accelerated by imMSC injection. As suggested in Figure 4(B), the nerve volume in NB group was less than in others (P < 0.01). The results showed that the nerve in NB+imMSCs/eSDF-1

MSC treatment accelerates the recovery of injured PN. (A) Representative images of PN staining in each group. Green is βIII-tubulin, blue is DAPI nucleus, and red is MSCs. Original magnification: ×200. (B) Positive rate of nerve in each group. Each bar shows the mean values (standard deviation). *P < 0.01 compared with NB group. **P < 0.05 compared with NB+imMSCs/eSDF-1
High SDF-1 Expression Improves the Quantity of VEGF and bFGF
Figure 5(A) shows that imMSCs increase VEGF expression. As depicted in Figure 5(B), imMSCs improve bFGF expression. Figure 5(a and b) shows that the levels of VEGF and bFGF in the NB+imMSCs/eSDF-1

SDF-1 improves VEGF and bFGF expression in vivo. (A) Representative images of VEGF stain for each group. Original magnification: ×200. (a) Positive rate of VEGF for each group. Each bar shows the mean values (standard deviation). #
P < 0.01 compared with NB group. ##
P < 0.01 compared with NB+imMSCs/eSDF-1
Apoptosis Decreases with MSC Injection Especially in a High SDF-1 Microenvironment
As shown in Figure 6(A), without MSCs the apoptosis in the damaged PN was high. However, after imMSC injection the apoptosis was improved. Figure 6(B) displays a positive rate of PARP in each group. We found a higher degree of apoptosis in the NB group (P < 0.01). Furthermore, the apoptosis in the NB+imMSCs/eSDF-1
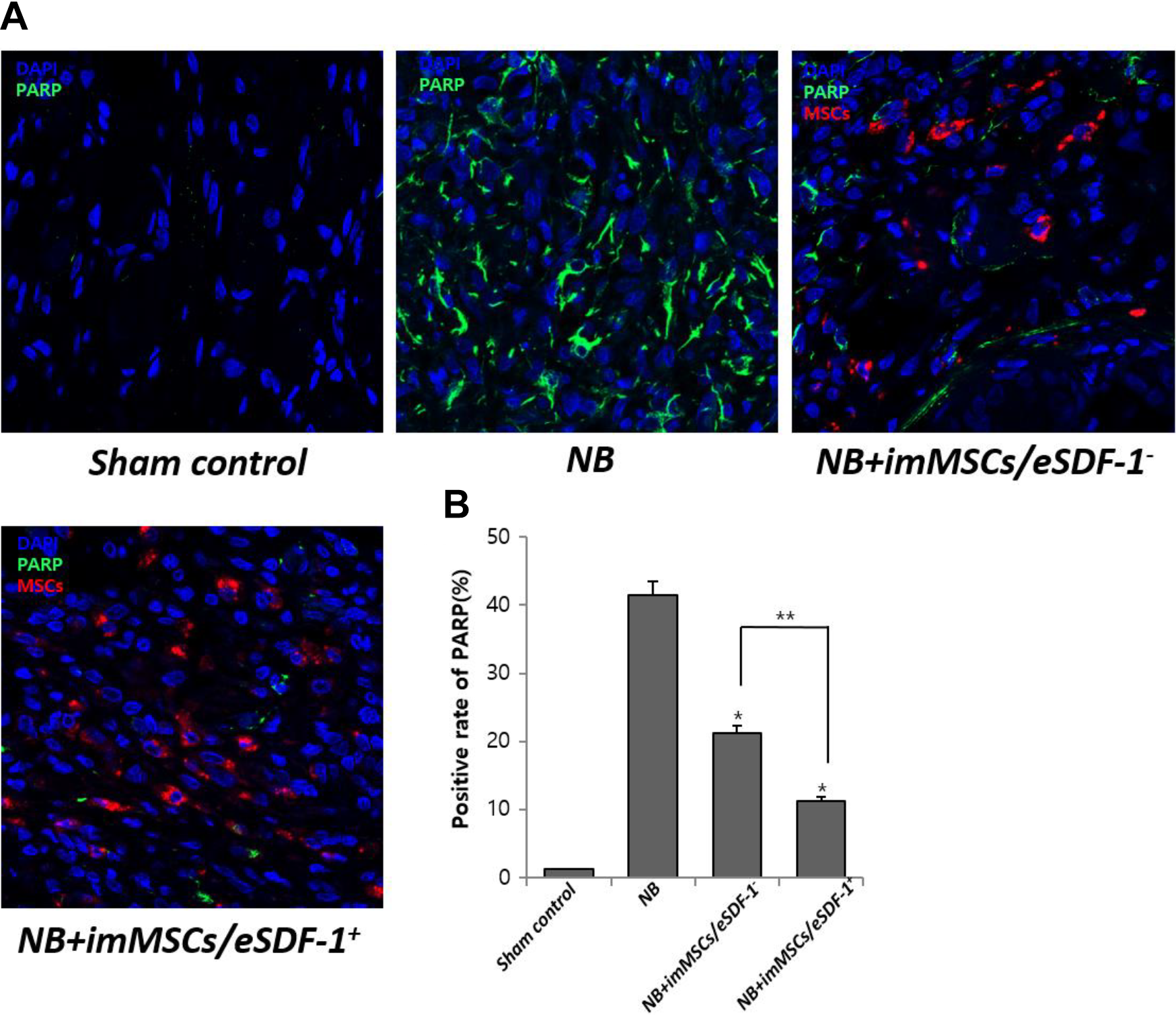
MSCs degrade apoptosis by increasing the expression of SDF-1. (A) Representative images of apoptosis for each group. Original magnification: ×200. (B) Positive rate of PARP for each group. Each bar shows the mean values (standard deviation). *P < 0.01 compared with NB group. *P < 0.05 compared with NB+imMSCs/eSDF-1
MSCs activate PI3K/AKT/mTOR and MAPK Signaling Pathway via enhanced SDF-1 Expression
In the first study investigating nerve recovery in a high SDF-1 microenvironment, we identified the underlying mechanism. As shown in Figure 7, p-AKT/AKT and p-ERK/β-actin levels in the NB+imMSCs/eSDF-1

(A) All groups were compared for nNOS, p-AKT, AKT, and p-ERK in PN by western blot. (B) Quantity analysis of western blot, including nNOS/β-actin, p-AKT/AKT, and p-ERK/β-actin. *P < 0.05 compared with values from NB group. **P < 0.05 compared with NB+imMSCs/eSDF-1
Discussion
Currently, the quality of life (QOL) during the postoperative period is of increasing concern 15 . QOL is an important aspect in the overall management of NB patients, for example, to evaluate treatment-related changes in a patient 16 . Pelvic surgeries including radical prostatectomy, rectum resection, and hysterectomy are associated with a risk of PN injury 17 –19 , which may trigger NB and subsequently lower the QOL postoperatively. Our results showed that NB decreased bladder function, reduced voided volume, and decelerate voiding frequency, which were ameliorated by imMSCs/eSDF-1. Obviously, QOL would be improved when NB was treated by imMSCs/eSDF-1. Li H et al. 20 illuminated that the regeneration of peripheral nerves after pelvic injury was a complex process involving neurons, Schwann cells, basal lamina, and end-organ responsiveness. Therefore, effective and potential treatments to recover the injured nerve and successfully cure NB are imperative. In this study, we established a rat model of NB and performed a series of investigations with the imMSCs engineered. We found that imMSCs injection led to a significant elevation in the mean intravesical pressure of NB rats and an obvious thickening of the smooth muscle of bladder wall. Meanwhile, we found that in NB rats higher levels of SDF-1 in PN facilitated the recovery. We found that exposure to imMSCs ameliorated experimental NB dysfunction.
In a recent report 21 , investigators established a rat model of bilateral cavernous nerve crush injury. After stem cell injection, the expression of neural markers in the penile dorsal nerves and the erectile dysfunction improved significantly, suggesting that stem cells facilitate the repair of injured nerve. In a previous study 14 , we also found some similar outcomes in a rat model of diabetes mellitus erectile dysfunction. However, in both the studies, neurotrophic cytokines were merely detected. In our study, the imMSCs around the injured nerve were directly observed. Under increased SDF-1 levels produced by imMSCs, the restoration of nerve was steadily enhanced. Ouyang X et al. 22 reported that MSCs enhanced the recovery of the injured smooth muscle. They injected MSC-derived exosomes into the corpus cavernosum and after 4 weeks the recovery of the smooth muscles in corpus cavernosum was improved. However, few studies focused on bladder restoration and nerve recovery with MSC treatment. In our study, we found that the injection of imMSCs resulted in a thickening of bladder wall in NB rats markedly. We found that under a high SDF-1 microenvironment, the VEGF expression was also increased. VEGF is the key to tissue restoration. VEGF activated the PI3K-Akt signaling pathway, which increased the expression of nNOS during the recovery of injured nerve 23,24 . However, VEGF stimulated the MAPK signaling pathway to induce further p-ERK synthesis and activated the gene proliferation 25 .
Naderi-Meshkin et al. 26 demonstrated that SDF-1 played an important role in MSC homing. Fandel et al. 27 also reported stem cell homing to the PN after injection accompanied by high SDF-1 expression in the PN. In this study, we observed after imMSCs injection, several imMSCs migrated to the damaged nerve as the SDF-1 expression was upregulated. Our findings suggested that the increased SDF-1 expression led to further aggregation of imMSCs around the injured nerve. We supposed that MSCs also expressed SDF-1, which suggested a positive feedback, induced by SDF-1 expression in the injured nerve. Injured nerves expressed SDF-1 inducing migration of MSCs, which further expressed SDF-1, and upregulated the concentration of SDF-1 rapidly, resulting in enhanced migration of MSCs to the target tissue. To verify our hypothesis, we used a gene transfer technique, which facilitated imMSCs expression of higher levels of SDF-1 compared with normal imMSCs. Based on our results, we found that the quality of stem cells and the effect of MSC treatment yielded better outcomes in the upregulated SDF-1 group compared with the normal SDF-1 group, which validated our hypothesis of a positive feedback mediated via SDF-1 between injured nerves and MSCs. In this positive feedback, nerve injury triggered higher expression of MSCs and their directional migration. Zhou F et al. 28 believed that MSCs spread rapidly, upon injection, and only a few MSCs reached the target area. A few studies 29 considered that reactive oxygen species were toxic to injected stem cells due to the induction of apoptosis, which reduced the levels of MSCs rapidly. Both studies reported that the injected MSCs failed to remain at the injection site. However, the homing mechanisms of MSCs driven by SDF-1 explain the congregation of MSCs around the injured nerve. Meanwhile, we used the immortalized MSCs to prevent apoptosis of stem cells, to generate a stable microenvironment 30 . In brief, we eliminated other effects induced by MSCs and an unstable microenvironment. Under such conditions, we demonstrated that imMSCs expressing a high level of SDF-1 had a positive effect on the restoration of injured nerves. The experiment involving MSC treatment was performed during an early stage of prostatitis.
There are still some defects in this article. First, we did not assess the viability of MSCs before injection, which decreased the accuracy of at least one part of the experiment. Comparing the viability before and after transfection, selection, and label would reduce errors by flow cytometry. Secondly, in the part of injured nerve regeneration, we just used the nerve staining and detection of nNOS expression. Adding a detection of some nerve biomarkers by a real-time polymerase chain reaction could make the results more convincing. At last, the mechanism research was not deep enough. So in the next study, we should focus on the pathway and factors to investigate the accurate mechanism of MSCs for NB.
Conclusions
The study demonstrated that high levels of SDF-1 expression induced rapid MSC homing to the target tissue and imMSCs/eSDF-1
Footnotes
Authors’ Contributions
Guan Qun Zhu, first author, was responsible for conception, study design, data analysis and interpretation, and manuscript writing. Seung Hwan Jeon was responsible for data collection and assembly, and data analysis. Kyu Won Lee and Hyuk Jin Cho were responsible for data collection and assembly. U Syn Ha, Sung Hoo Hong, and Eun Bi Kwon were responsible for providing the study material and animal models. Ji Youl Lee was responsible for conception and design, and data analysis and interpretation. Hyo-Jin Kim, Soon Min Lee, and Hey-Yon Kim were responsible for the design of cellular experiment. Woong Jin Bae and Sae Woong Kim, corresponding authors, were responsible for conception and design, financial support, drafting and revision of the manuscript, and final approval of the manuscript. All authors read and approved the final manuscript.
Availability of Data and Materials
The datasets supporting the conclusions of this article are included in this article.
Ethical Approval
This study was approved by the Institutional Animal Care and Use Committee of the Catholic University of Korea (CUMC-2016-0218-01).
Statement of Human and Animals Rights
This article does not contain studies with human subjects. Animal experiments were reviewed and approved by the Institutional Animal Care and Use Committee of the Catholic University of Korea (CUMC-2016-0218-01).
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Ministry of Science & ICT (NRF-2018M3A9E8020861)