
Abstract
Hyperbaric oxygen (HBO) therapy may promote neurological recovery from hypoxic-ischemic encephalopathy (HIE). However, the therapeutic effects of HBO and its associated mechanisms remain unknown. The canonical Wnt/β-catenin signaling pathways and bone morphogenetic protein (BMP) play important roles in mammalian nervous system development. The present study examined whether HBO stimulates the differentiation of neural stem cells (NSCs) and its effect on Wnt3/β-catenin and BMP2 signaling pathways. We showed HBO treatment (2 ATA, 60 min) promoted differentiation of NSCs into neurons and oligodendrocytes in vitro. In addition, rat hypoxic-ischemic brain damage (HIBD) tissue extracts also promoted the differentiation of NSCs into neurons and oligodendrocytes, with the advantage of reducing the number of astrocytes. These effects were most pronounced when these two were combined together. In addition, the expression of Wnt3a, BMP2, and β-catenin nuclear proteins were increased after HBO treatment. However, blockade of Wnt/β-catenin or BMP signaling inhibited NSC differentiation and reduced the expression of Wnt3a, BMP2, and β-catenin nuclear proteins. In conclusion, HBO promotes differentiation of NSCs into neurons and oligodendrocytes and reduced the number of astrocytes in vitro possibly through regulation of Wnt3/β-catenin and BMP2 signaling pathways. HBO may serve as a potential therapeutic strategy for treating HIE.
Keywords
Introduction
Hypoxic-ischemic encephalopathy (HIE) is the primary cause of neonatal cerebral injury 1 , with an incidence of 1.5 cases per 1000 full-term live births 2 , and may result in longer-term neurological deficits, such as cerebral palsy, epilepsy, and mental retardation, or death 3 –7 . Thus, it is essential to identify an effective treatment for HIE. In the central nervous system, the self-reproduction and multiple differentiation capacity of neural stem cells (NSCs) can be harnessed to repair the injured brain 8 . Hypoxic ischemia can, by itself, activate endogenous NSCs and promote their proliferation and differentiation; however, this type of ability is limited 9 . Therefore, other effective methods are required to enhance neurogenesis after brain injury.
Hyperbaric oxygen (HBO) therapy is a safe, noninvasive treatment that can elevate oxygen tension in tissues 10 . In China, HBO therapy [<3.0 atmospheric absolute (ATA)] has been widely used for treating HIE for many years without causing retinopathy of prematurity 11 . Experiments with neonatal rats have previously demonstrated that HBO therapy may treat the neurological damage which was caused by hypoxic-ischemic brain damage (HIBD) through inhibiting neural apoptosis and promoting NSC proliferation 12 –15 . However, the therapeutic effects of HBO and the mechanisms remain unknown.
The canonical Wnt/β-catenin and bone morphogenetic protein (BMP) signaling pathways play essential roles in NSC fate 16 –20 . Wnt/β-catenin signaling controls the multiplication of NSCs and the alteration of cell types 21 –23 . Canonical Wnt signaling is activated by Wnt ligands, leading to increase of β-catenin in the cytoplasm, followed by an elevated β-catenin in the nucleus which regulates the expression of several genes 24,25 . BMPs (secreted extracellular glycoproteins) influence many biological functions 26 . To date, approximately 20 BMPs have been identified, all of which belong to the transform growth factor β superfamily 27 . BMPs can inhibit the proliferation of NSCs and influence cell fate 28,29 . The role of BMPs, even in the same location or the same type of cell, is controversial 14,30 . Previous studies have revealed that HBO therapy promoted NSC proliferation by increasing the expression of Wnt3 and β-catenin 15,31 , resulting in neurological recovery of neonatal HIBD rats possibly through increasing the expression of BMP4 protein 14 . However, the exact mechanism by which Wnt/β-catenin and BMP are involved in the differentiation of NSCs cultured with rat HIBD tissue extracts after HBO treatment in vitro remains poorly understood. The aim of this present study was to investigate the effect of HBO on the differentiation of NSCs, as well as on Wnt3/β-catenin and BMP2 signaling pathways.
Materials and Methods
Rat NSC Cultures
Cerebral cortical tissue was excised from 3-day-old Sprague–Dawley rats, cut into slices, and then disaggregated in 0.25% trypsin. The tissues were agitated, filtered, and centrifuged, then suspended in DMEM: F12 medium (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA). NSCs were cultured in serum-free DMEM: F12 medium containing 20 µl/ml B27 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 10 ng/ml bFGF (Pepro Tech, Rocky Hill, NJ, USA) and 20 ng/ml EGF (PeproTech, Inc.). Cells were maintained at 37°C in an incubator with 5% CO2.
All animals in the present study were purchased from the Animal department, XiangYa School of Medicine, and the experimental protocols were evaluated and approved by the Animal ethics committee of XiangYa School of Medicine, Central South University (Changsha, China).
Animal Model of HIBD
The HIBD rat model was established using 7-day-old Sprague–Dawley rats, weighing 12.5–15.4 g in accordance with the Rice procedure 32 , in which neonatal rats were anesthetized with ether (place cotton balls containing ether into the beaker, place rats in, then sealed. The rats were excited and then suppressed, and fell down. When the cornea was slow and muscle tension was reduced, the animal was removed. This process took about 4–6 min). The skin was disinfected using 75% alcohol and a midline neck midline incision made of about 1–1.5 cm. The skin, subcutaneous fat, muscle, and the left carotid artery was separated in turn. The left common carotid artery was ligated with sterilized silk thread, and the blood vessel was cut in the middle of ligation, and the wound closed. Finally, the skin was disinfected again. The whole process was completed within 5–10 min. One hour later, rats were placed in a plastic hypoxia container (8% oxygen balanced with nitrogen) at a temperature of 36 ± 1°C for 2 h. Once these procedures were completed, rats were returned to their mothers.
Rat Brain Tissue Extracts
Next, hypoxic-ischemic brain tissue extracts were removed from rats using the procedure described by Wang et al 15 . Briefly, the left ischemic hemispheres were isolated from rats that had suffered from HIBD for 24 h. The left hemispheres of normal rats were used as the control group. Tissues were weighed and mixed with DMEM/F12 medium containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.). Finally, the tissue was placed on ice, centrifuged for 15 min at 10,000 rpm/min at 4°C followed by collection of the supernatant, which was stored at –80°C.
NSC Passage and Culturing with Brain Tissue Extract
NSC passage and purification method is as follows: neurospheres which grow to about 200 µm will be passaged. First, the medium and cell group were transferred together through a straw to a 15 ml centrifugal tube, centrifuged at 200 g for 5 min, and the supernatant discarded. Fresh DMEM/F12 medium (containing 20 µl/ml B27, 10 ng/ml bFGF, 20 ng/ml EGF) was added. The neurosphere was blown repeatedly with a syringe into a single cell group or small cells group, and then subcultured at 1:3. Two to three passages of NSCs were collected, centrifuged at 10,000 rpm/min for 5 min, washed twice with PBS, and then cultured in DMEM/F12 medium containing 10% FBS and brain tissue extraction (either HIBD or control normal brain). The volume of the brain tissue extract was equal to half of the medium applied.
In Vitro Inhibitors of Wnt and BMP Signaling Pathways
Recombinant secreted Frizzled-Related Protein 2 and 3 (sFRP2/3) (R&D Systems, Minneapolis, MN, USA) was added at a concentration of 500 ng ml-1; recombinant noggin protein (R&D Systems) was added at a concentration of 100 ng ml-1.
HBO Therapy
HBO therapy was performed as described by Zhang et al 31 . The NSCs were cultured in DMEM/F12 medium containing 10% FBS, or a mixture of DMEM/F12 medium containing 10% FBS and HIBD brain tissue extract for 1 h. HBO treatment was then conducted in an infant HBO chamber (YLC0.5/1A; 701 Institute of China State Shipbuilding Corporation, Wuhan, China). The infant HBO chamber was perfused with constant 100% oxygen and its pressure reached 1–3 ATA; at the same time, the oxygen concentration in the cabin was maintained at 85% or higher. After HBO treatment, cells were cultured in an incubator at 37°C with 5% CO2.
Cytoplasmic and Nuclear Protein Extracts
All procedures were carried out at 4°C. NSCs were collected, washed twice with PBS, and centrifuged at 6000 rpm/min for 8 min. A nuclear and Cytoplasmic Protein Extraction Kit (Beyotime Institute of Biotechnology, Haimen, China) was used to separate cytoplasmic and nuclear extracts according to the manufacturer’s instructions.
EdU Proliferation Experiment
Five groups of NSC suspensions were inoculated in 96-well plates. EdU solution was diluted with DMEM/F12 medium containing 10% FBS and incubated for 24 h. Then, cells were incubated with 50 µl fixative solution (i.e., PBS containing 4% paraformaldehyde) for 30 min at room temperature, followed by 50 µl 2mg/ml glycine, decolorized in a shaker for 5 min, washed with PBS for 5 min, and incubated with 100 µl permeant for 10 min. Then 100 ul Apollo staining reaction solution was added, and the sample was incubated in the dark at room temperature in a decolorization shaker for 30 min; the dyeing reaction solution was then discarded. After adding 100 µl PBS-Triton, the sample was placed in a decolorizing shaker 2–3 times, 10 min each time, and the penetrant discarded. Finally, Hoechst 33342 reaction solution was added. The images were captured using an Olympus confocal imager. The count was performed under a microscope. Six non-overlapping fields were randomly taken from each film, the percentage of EdU-positive cells was calculated, and the mean value was obtained (6 percent plus 6). Proliferation rate = EdU-positive cells/Hoechst-33342 positive cells×100%.
Western Blotting
Cells were dissolved in a buffer containing 200 mM LDTT, 20% glycerol, 4% SDS, and 100 mM Tris-HCl of pH 6.8. The cell lysates were boiled in sample buffer for 5 min, separated by 10% SDS-PAGE, and then transferred to a PDVF membrane (EMD Millipore, Billerica, MA, USA). At room temperature, the membrane was blocked with blocking solution (2% BSA, Wuhan Boster Biological Technology, Ltd., Wuhan, China) for 4 h, followed by incubation with primary antibodies at 4°C for 12 h. The membrane was washed three times using PBS containing 0.1% Tween-20, followed by a 1 h incubation with secondary antibodies at room temperature. The following primary antibodies were used: Rabbit mAb active Wnt3 (1:100; SC-28824, Santa Cruz, Biotechnology, Inc., Dallas, TX, USA), rabbit mAb active β-catenin (1:100; BA0426, Wuhan Boster Biological Technology, Ltd.), rabbit mAb active BMP2 (1:100; PB0727, Wuhan Boster Biological Technology, Ltd.), mouse mAb β-actin (1:200, BM0626, Wuhan Boster Biological Technology, Ltd.) and mouse mAb active PCNA (1:1000; 610664, BD Biosciences, San Jose, CA, USA). The secondary antibodies were HRP-conjugated Anti-Mouse IgG (1:500; BM2002, Wuhan Boster Biological Technology, Ltd.) and HRP-conjugated Anti-Rabbit IgG (1:500; BA1070, Wuhan Boster Biological Technology, Ltd.). Images were captured using an Odyssey scanner and band intensities were analyzed by Image Studio Lite 5.2 (LI-COR Biosciences, USA)
Immunocytochemistry
At room temperature the cells were fixed for 40 min using 4% buffered paraformaldehyde. Slides were permeabilized for 10 min using 0.5% Triton X-100, then blocked with 2% BSA (Wuhan Boster Biological Technology, Ltd.) for 1 h. Primary antibodies rabbit anti-rat neuron-specific enolase (NSE; 1:50; PA1061, Wuhan Boster Biological Technology, Ltd.), rabbit anti-rat microtubule-associated proteins 2 (MAP2, Abcam, ab32454, Shanghai, China), rabbit anti-rat glial fibrillary acidic protein (GFAP; 1:50; A00213, Wuhan Boster Biological Technology, Ltd.), mouse anti-rat Oligodendrocyte Marker O4 (O4; 1: 250; MAB345, Chemicon, Temecula, CA, USA), and rabbit anti-rat Myelin Basic Protein (MBP, Abcam, ab7349) were added to the tissues and incubated at 4°C for 12 h followed by incubation with the following secondary antibodies: FITC-conjugated anti-rabbit IgG (1:100; BA1105, Wuhan Boster Biological Technology, Ltd.), CY3-coupled anti-mouse IgG (1:100; BA1031, Wuhan Boster Biological Technology, Ltd.), and CY3-coupled anti-rabbit IgG (1:100; BA1032, Wuhan Boster Biological Technology, Ltd.). The nuclei were stained using Hoechst 33258. The images were captured using an Olympus confocal microscope.
Differential Cell Count
Using a fluorescence microscope, six non-overlapping fields were randomly selected in each slide, and the average number of cells was subsequently obtained from six independent experiments. The following formulas were applied: Percentage of NSCs differentiated into neurons or glial cells = (number of NSE-positive cells / number of Hoechst 33258-positive cells) × 100%; (number of GFAP-positive cells / number of Hoechst 33258-positive cells) × 100%; (number of O4-positive cells / number of Hoechst 33258-positive cells) × 100%.
Statistical Analysis
SPSS 17.0 was applied for processing data. Data were presented as the mean ± standard deviation (SD). Statistical differences were calculated using one-way analysis of variance (S-N-K). Differences between groups were considered to be significant when p < 0.05. SPSS17.0 (IBM SPSS software) was used for the analysis.
Results
Effect of Different Conditional HBO Treatments on the Differentiation of NSCs In Vitro
The NSCs maintained in DMEM/F12 medium containing 10% FBS were randomly divided into seven groups: The CON group (untreated, control group) and six HBO treatment groups; (i) 1 ATA-30 min group; (ii) 1 ATA-60 min group; (iii) 2 ATA-30 min group; (iv) 2 ATA-60 min group; (v) 3 ATA-30 min group; and (vi) 3 ATA-60 min group. The NSCs were stimulated with HBO, and subsequently cultured in DMEM/F12 medium containing 10% FBS for 24 h. After 24 h, NSE cell expression was examined and showed that NSE cell expression gradually increased (Fig. 1A). However, 1 ATA-30 min group and CON group were not statistically different (p > 0.05, Fig. 1B), whereas for the remaining HBO groups and CON group, under the same pressure, the 60 min and 30 min groups were statistically different (p < 0.05, Fig. 1B). NSE cell expression was the highest in the 2 ATA-60 min group (p < 0.05, Fig. 1B). These findings indicated that HBO treatment at 2 ATA for 60 min had the strongest effect on inducing the differentiation NSCs into neurons in vitro.
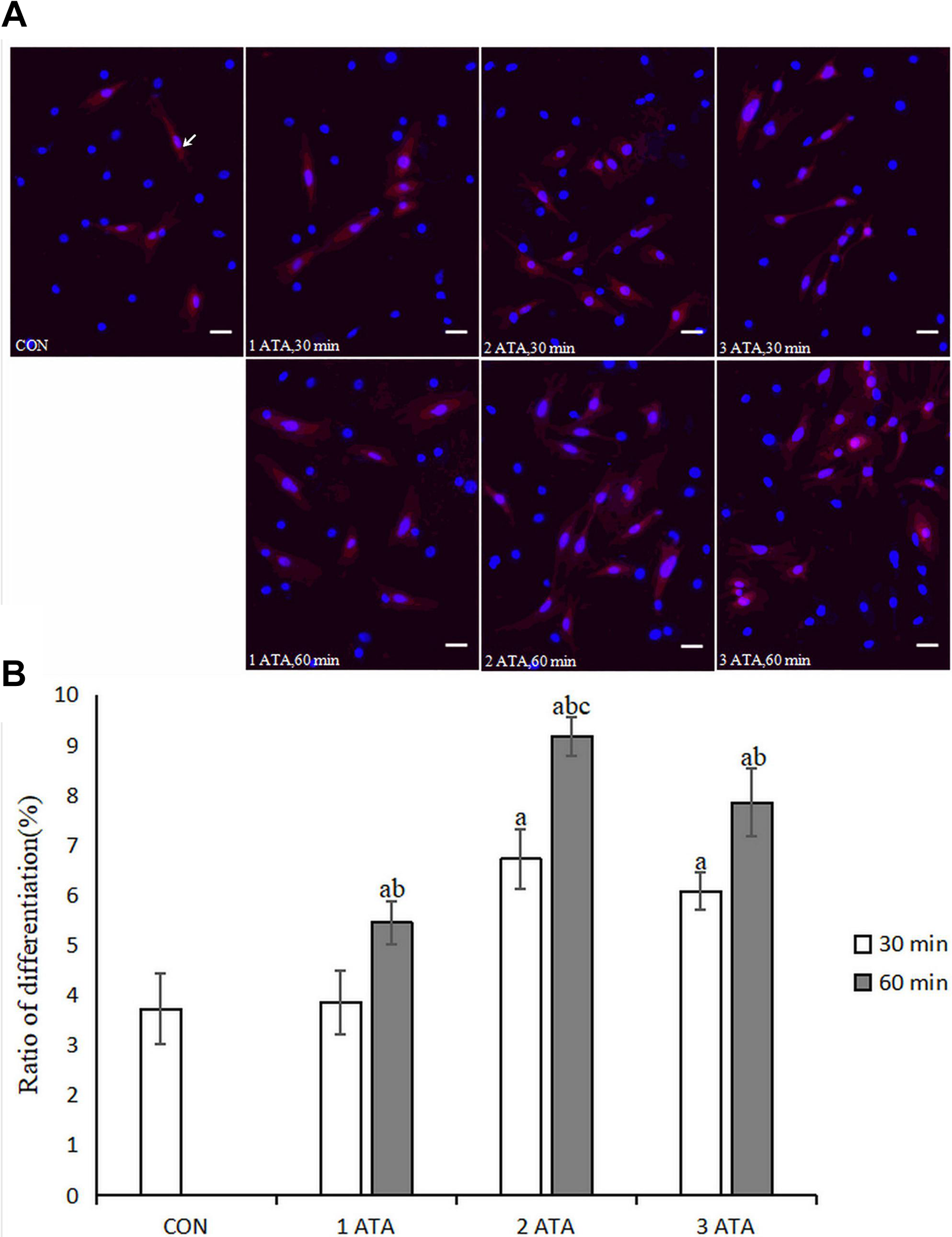
Differentiation of NSCs. (A) The differentiated NSCs were double labeled for CY3 (red) and Hoechst 33258 (blue) by immunofluorescence. NSE-positive NSCs were labeled with primary antibodies rabbit anti-rat NSE. Scale bar = 100μm. The expression of NSE in cells with HBO treatment (2 ATA, 60 min) exhibited the maximum number of differentiated NSCs. (B) Each bar represents mean±SD, n = 6. a p < 0.05 vs. the CON group, ANOVA test; b p < 0.05 vs. 30 min group under the same pressure, ANOVA test; c p < 0.05 vs. all groups, ANOVA test. NSCs: neural stem cells; NSE: neuron-specific enolase; HBO: hyperbaric oxygen; CON: control.
HBO Promotes the Differentiation of NSCs into Neurons and Oligodendrocytes
Next, we evaluated the effects of HBO treatment as well as how brain tissue extracts influence the differentiation of NSCs. Cells were randomly divided into five groups: (i) CON group: no treatment; (ii) HBO group: HBO treatment (2 ATA, 60 min); (iii) NBH group: medium containing normal brain tissue extracts; (iv) HIBH group: medium containing HIBD brain tissue extracts; and (v) HIBH+HBO group: medium containing HIBD brain tissue extracts for 1 h then subjected to HBO treatment (2 ATA, 60 min).
Compared with the CON group, the expression of NSE, MAP2, MBP, and O4 in the cells of the HBO, NBH, HIBH, and HIBH+HBO groups was gradually increased (p < 0.05, Fig. 2A) with the highest expression of NSCs in the HIBH+HBO group, suggesting successful differentiation into neurons and oligodendrocytes (p < 0.05, Fig. 2B). Interestingly, the expression of GFAP in NBH, HIBH, and HIBH+HBO groups was significantly lower than that in CON and HBO groups (p < 0.05, Fig. 2A). Taken together, these data indicated that HBO can increase the differentiation of NSCs into neurons and oligodendrocytes in vitro, without affecting their differentiation into astrocytes. Normal and hypoxic-ischemic brain tissue extracts increased NSC differentiation into neurons and oligodendrocytes and had the advantage of reducing the number of astrocytes. The effects of HIBD brain extracts were better than those associated with normal brain tissue extracts, and the effects of HIBD and HBO together were the most effective. Used in combination, HIBD and HBO were the most successful in causing NSCs to differentiate into neurons and oligodendrocytes and had the advantage of reducing the number of astrocytes.
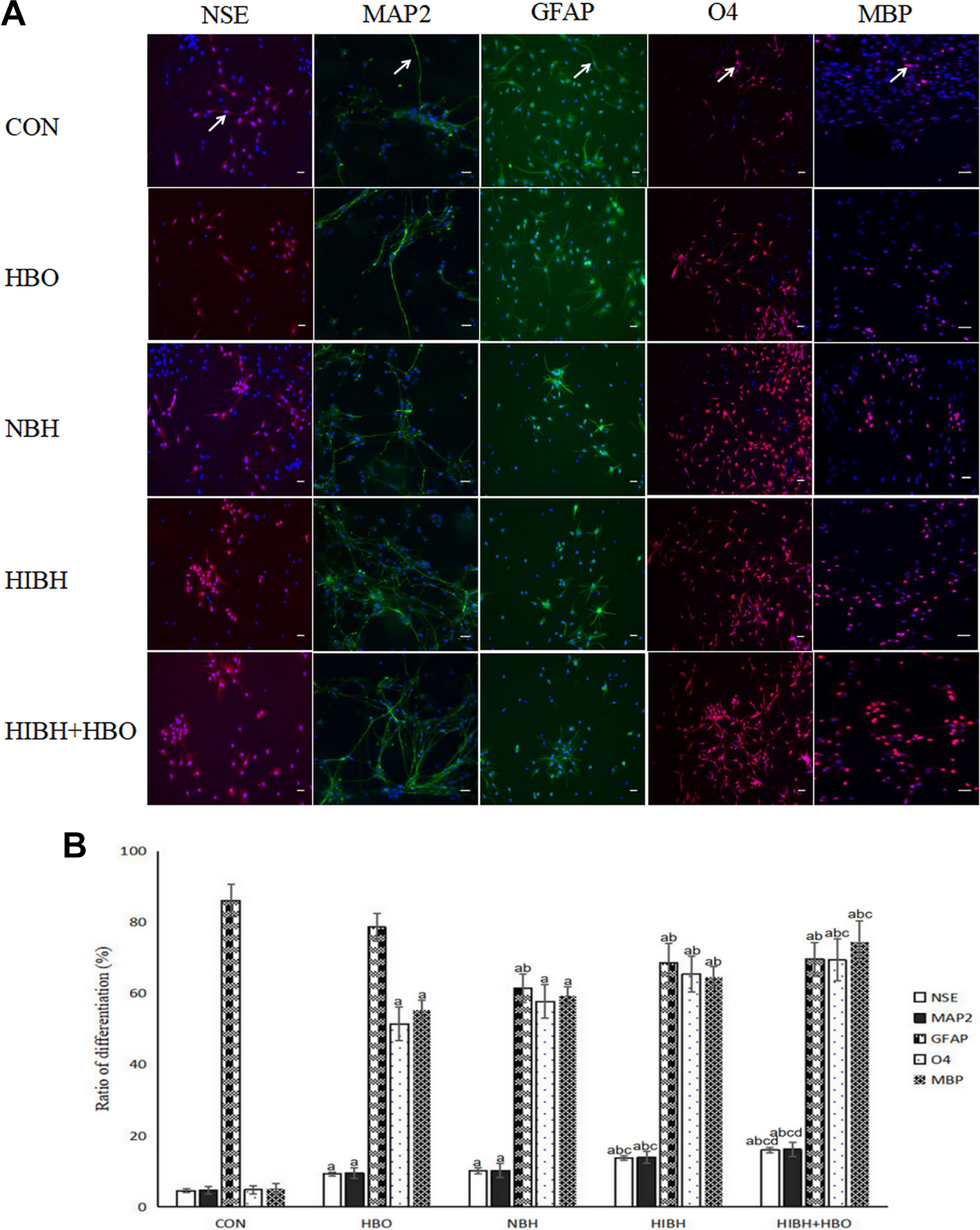
Differentiation of NSCs under different conditions. (A) The differentiated NSCs were double labeled for Fluor 555 (red) or Fluor 488 (green) and Hoechst 33258 (blue) by immunofluorescent staining. NSE-positive NSCs were labeled with primary antibodies against rabbit anti-NSE; MAP2-positive NSCs were labeled with primary antibodies against rabbit anti-MAP2; GFAP-positive NSCs were labeled with primary antibodies against rabbit anti-GFAP; O4-positive NSCs were labeled with primary antibodies against mouse anti-O4; and MBP-positive NSCs were labeled with primary antibodies against rabbit anti-MBP. Scale bars=20μm. HBO increased NSC differentiation into neurons and oligodendrocytes in vitro, but it did not affect its differentiation into astrocytes. When used in combination, HIBD and HBO were the most effective in causing NSCs to differentiate into neurons and oligodendrocytes and had the advantage of reducing the number of astrocytes produced. (B) Each bar represents mean±SD, n = 6. a p < 0.05 vs. the CON group, ANOVA test; b p < 0.05 vs. the HBO group, ANOVA test; c p < 0.05 vs. the NBH group, ANOVA test; d p < 0.05 vs. the HIBH group, ANOVA test. NSCs: neural stem cells; HBO: hyperbaric oxygen; CON: control; NBH: normal brain tissue extracts; HIBH: hypoxic-ischemic brain damage tissue extracts; NSE: neuron-specific enolase; MAP2: Microtubule-Associated Protein2; GFAP: glial fibrillary acidic protein; O4: Oligodendrocyte Marker O4; MBP: Myelin Basic Protein.
Effect of Different Conditional HBO Treatments on the Proliferation of NSCs In Vitro
In order to further verify the effect of HBO on NSC proliferation, we conducted EDU detection on NSC proliferation. The results showed that NSCs cell proliferation activity levels of CON group, HBO group, NBH, HIBH, HIBH+HBO group were increased in turn; the differences between the groups were statistically significant (p < 0.05, Fig. 3A and B). The cell proliferation activity of HIBH + HBO group is the highest among these groups. The results showed that HIBD combined with HBO had the best effect on promoting the proliferation of neural stem cells.

Proliferation of NSCs. (A) The proliferation NSCs were double labeled for EDU (red) and Hoechst 33342 (blue) by immunofluorescence. Scale bar = 20μm. (B) Each bar represents mean±SD, n = 6. a p < 0.05 vs. the CON group, ANOVA test; b p < 0.05 vs. the HBO group, ANOVA test; c p < 0.05 vs. the NBH group, ANOVA test; d p < 0.05 vs. the HIBH group, ANOVA test. NSCs: neural stem cells; HBO: hyperbaric oxygen; CON: control; NBH: normal brain tissue extracts; HIBH: hypoxic-ischemic brain damage tissue extracts.
HBO Increases the Expression of Wnt3a, BMP2, and β-Catenin Nuclear Proteins
Western blotting analysis indicated that the expression of Wnt3a, BMP2, and nuclear β-catenin proteins were increased gradually across the CON, HBO, NBH, HIBH and HIBH+HBO groups (Fig. 4A and B). However, no difference was observed between the HBO and NBH groups (p > 0.05, Fig. 4C and D), whereas the remaining groups were statistically different when compared with each other (p < 0.05, Fig. 4C and D). Cytoplasmic β-catenin protein levels remained the same across the different groups (p > 0.05, Fig. 4B and D). These results indicated that HBO promoted the expression of Wnt3a, BMP2, and nuclear β-catenin proteins.
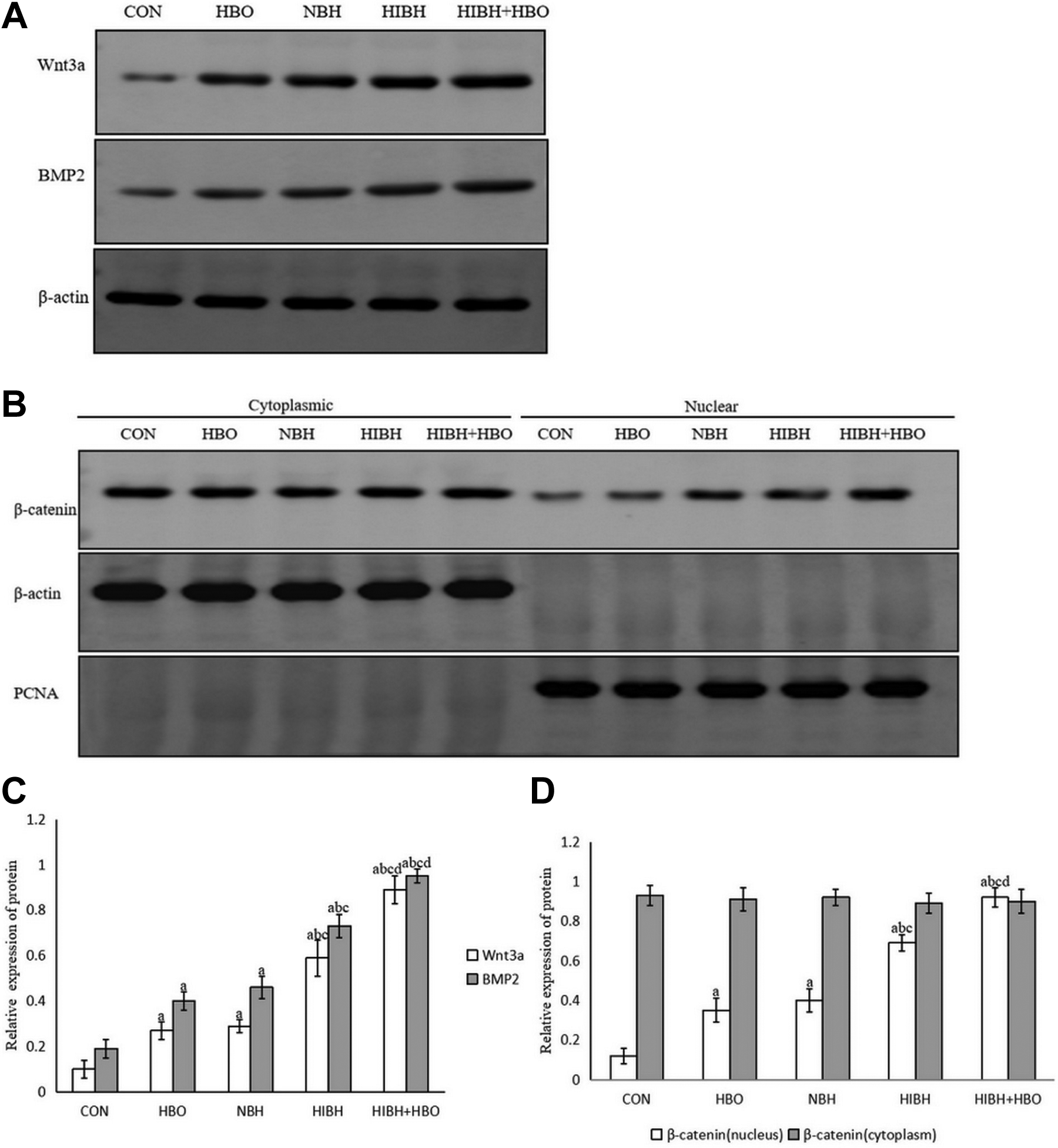
Expression of Wnt3a, BMP2 and β-catenin nuclear proteins in NSCs after HBO treatment by Western blotting. (A) Expression of Wnt3a and BMP2 protein: HBO treatment up-regulated the expression of Wnt3a and BMP2 proteins. (B) Expression of β-catenin nuclear and cytoplasmic proteins: HBO up-regulated the expression of β-catenin nuclear protein; however, the expression of β-catenin cytoplasmic protein was not affected. (C and D) Each bar represents mean±SD, n = 3. a p < 0.05 vs. the CON group, ANOVA test; b p < 0.05 vs. the HBO group, ANOVA test; c p < 0.05 vs. the NBH group, ANOVA test; d p < 0.05 vs. the HIBH group, ANOVA test. NSCs: neural stem cells; HBO: hyperbaric oxygen; CON: control; NBH: normal brain tissue extracts; HIBH: hypoxic-ischemic brain damage tissue extracts; BMP2: bone morphogenetic protein.
Inhibition of Wnt Attenuates HBO-induced NSC Differentiation into Neurons and Oligodendrocytes
To examine the effects of Wnt on HBO-induced NSC differentiation, Wnt inhibitor sFRP2/3 was used. The expression of NSE and O4 in the cells of the HIBD+HBO+sFRP2/3 group was significantly lower than that in the HIBD+HBO group (p < 0.01, Fig. 5). However, the expression of GFAP in the cells of HIBD+HBO+sFRP2/3 group was significantly higher than that in HIBD+HBO group (p < 0.01, Fig. 5). In addition, the expression of NSE and O4 in the cells of HBO+sFRP2/3 group was significantly lower when compared with that in the HBO group (p < 0.01, Fig. 5). However, the expression of GFAP in HBO+sFRP2/3 cells was not affected (p > 0.05, Fig. 5). These results suggested that inhibition of Wnt by sFRP2/3 decreased NSC differentiation into neurons and oligodendrocytes.

Differentiation of NSCs in the presence or absence of sFRP2/3. (A) The differentiated NSCs were double labeled for CY3 (red) or FITC (green) and Hoechst 33258 (blue) by immunofluorescent staining. NSE-positive NSCs were labeled with primary antibodies against rabbit anti-NSE; GFAP-positive NSCs were labeled with primary antibodies against rabbit anti-GFAP; and O4-positive NSCs were labeled with primary antibodies against mouse anti-O4. Magnification, ×100. Scale bar = 300 μm. sFRP2/3 decreased NSC differentiation into neurons and oligodendrocytes after HBO treatment. (B) Each bar represents mean±SD, n = 6. a p < 0.01 vs. the HIBD+HBO group, ANOVA test; b p < 0.01 vs. the HBO group, ANOVA test. NSCs: neural stem cells; HBO: hyperbaric oxygen; CON: control; HIBD: hypoxic-ischemic brain damage; NSE: neuron-specific enolase; GFAP: glial fibrillary acidic protein; O4: Oligodendrocyte Marker O4; sFRP2/3: secreted Frizzled-related protein 2 and 3.
Wnt Inhibition Reduces the Expression of Wnt3a, BMP2, and β-Catenin Nuclear Proteins after HBO Treatment
The role of sFRP2/3 in the Wnt/β-catenin signaling pathway after HBO treatment was evaluated by Western blotting and showed the expression of Wnt3a, BMP2, and nuclear β-catenin was significantly decreased after sFRP2/3 treatment (p < 0.05, Fig. 6A and B); whereas the expression of cytoplasmic β-catenin was not affected (p > 0.05, Fig. 6B and D). These results indicated that sFRP2/3 reduces the expression of Wnt3a, BMP2, and nuclear β-catenin proteins after HBO treatment.
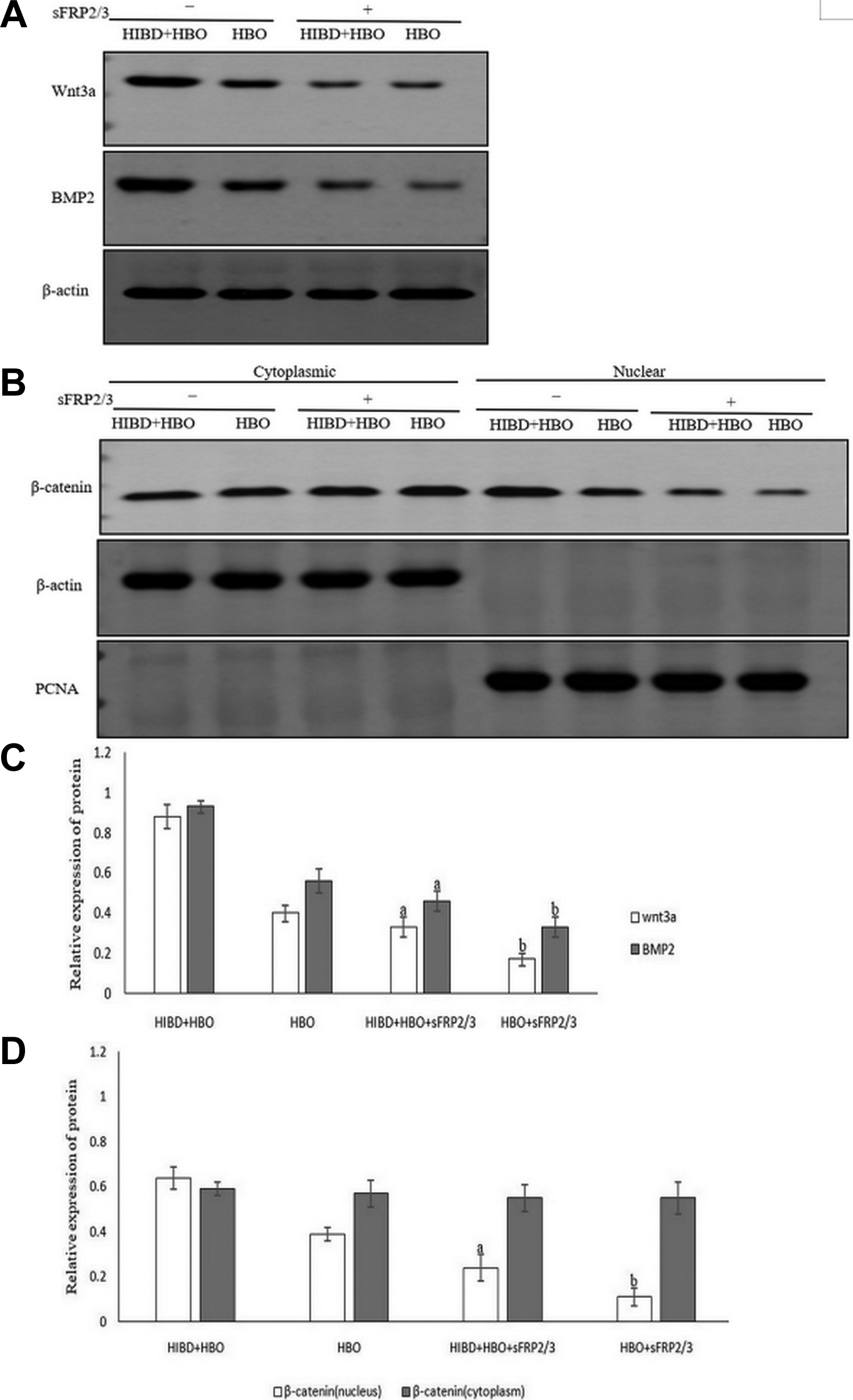
Expression, after HBO treatment, of Wnt3a, BMP2 and β-catenin nuclear proteins in the presence of sFRP2/3 was determined by Western blotting. (A) Expression of Wnt3a and BMP2 proteins: sFRP2/3 reduced the expression of Wnt3a and BMP2 proteins after HBO treatment. (B) Expression of β-catenin nuclear and cytoplasmic proteins: sFRP2/3 reduced the expression of β-catenin nuclear protein after HBO treatment; the expression of β-catenin cytoplasmic protein was not affected. (C and D) Each bar represents mean±SD, n = 3. a p < 0.05 vs. the HIBD+HBO group, ANOVA test; b p < 0.05 vs. the HBO group, ANOVA test. HBO: hyperbaric oxygen; CON: control; HIBD: hypoxic-ischemic brain damage; sFRP2/3: secreted Frizzled-related protein 2 and 3; BMP2: bone morphogenetic protein.
Noggin Reduces the Differentiation of NSCs and the Expression of BMP2
To investigate the effects of noggin on NSC differentiation, NSCs were treated with HBO in the presence or absence of noggin. In the presence of noggin, the expression of NSE, GFAP, and O4 was significantly decreased (p < 0.05, Fig. 7A and C). In addition, the expression of BMP2 protein was significantly decreased in the presence of noggin (p < 0.05, Fig. 7B and D). These results suggested that in vitro noggin reduced NSC differentiation into neurons, astrocytes and oligodendrocytes together with reduced expression of BMP2 after HBO treatment.

Differentiation of NSCs and the expression of BMP2 protein in the presence of noggin after HBO treatment. (A) Differentiation of NSCs: The differentiated NSCs were double labeled for CY3 (red) or FITC (green) and Hoechst 33258 (blue) by immunofluorescent staining. NSE-positive NSCs were labeled with primary antibodies against rabbit anti-NSE; GFAP-positive NSCs were labeled with primary antibodies against rabbit anti-GFAP; and O4-positive NSCs were labeled with primary antibodies against mouse anti-O4. Noggin reduced NSC differentiation into neurons, astrocytes and oligodendrocytes. Scale bars = 300 μm. (B) The expression of BMP2 protein by Western blotting: Noggin reduced the expression of BMP2 protein. (C and D) Each bar represents mean±SD. a p < 0.01 vs. the HIBD+HBO group, n = 6, ANOVA test; b p < 0.01 vs. the HBO group, n = 6, ANOVA test; c p < 0.05 vs. the HIBD+HBO group, n = 3, ANOVA test; d p < 0.05 vs. the HBO group, n = 3, ANOVA test. NSCs: neural stem cells; HBO: hyperbaric oxygen; CON: control; HIBD: hypoxic-ischemic brain damage; NSE: neuron-specific enolase; GFAP: glial fibrillary acidic protein; O4: Oligodendrocyte Marker O4; BMP2: bone morphogenetic protein.
Discussion
Hypoxic ischemia can cause a large number of ischemic cells to die from necrosis or apoptosis 33 . Previous studies have indicated that HBO treatment can promote long-term neurological recovery in neonatal HIBD rats by inhibiting neural apoptosis and promoting NSC proliferation 14,15 . However, the underlying mechanisms of neuroprotection after HBO treatment remain unclear. Micro-environments around NSCs and environmental oxygen levels can affect their proliferation and cell fate 34 –38 . HBO treatment at 2 ATA for 60 min was the most effective in inducing the differentiation of NSCs into neurons and oligodendrocytes in vitro without affecting their differentiation into astrocytes. Normal and HIBD tissue extracts increased NSC differentiation into neurons and oligodendrocytes and had the advantage of reducing the number of astrocytes, with better effects observed for HIBD brain extracts than normal brain tissue extracts. However, the effects of combination of HIBD and HBO treatment were the best, indicating that various micro-environments may modify the differentiation of NSCs into neurons and glial cells.
Mazumdar et al. 38 suggested that O2 availability may regulate stem cells directly via the Wnt/β-catenin signaling pathway. Consistent with this, in the present study, we found that HBO promotes the differentiation of NSCs possibly by regulating the Wnt3/nuclear β-catenin and BMP2 signaling pathways, as the expression of Wnt3a, BMP2 and nuclear β-catenin proteins was significantly increased after HBO treatment. However, inhibition of the Wnt signaling pathway by sFRP2/3 39,40 significantly decreased the differentiation of NSCs into neurons and oligodendrocytes together with reduced expression of Wnt3a, BMP2, and nuclear β-catenin proteins. In addition, inhibition of BMP signaling pathway by noggin, an extracellular antagonist of the BMP signaling pathway 26 , decreased NSC differentiation into neurons, astrocytes, and oligodendrocytes together with reduced expression of BMP2 protein. Recent research has revealed that canonical Wnt/β-catenin signaling can work synergistically with BMP signaling in order to influence the multiplication of NSCs and the alteration of cell types 41 –43 . In accordance with this, our present study indicated that Wnt3a signaling indirectly regulates the differentiation of NSCs through modulating BMP2 expression. Therefore, HBO treatment may influence neurogenesis and gliogenesis via cooperation between the Wnt3a and BMP2 signaling pathways.
HBO treatment has previously been shown to increase the resistance of tissue with reactive oxygen species, decrease the ultrastructural damage due to hypoxic-ischemia 13 , inhibit neural apoptosis 14 , and promote NSC proliferation 15 . The results of the present study support these previous findings, revealing that in vitro HBO promotes differentiation of NSCs to differentiate into neurons and oligodendrocytes and reduced the number of astrocytes produced when cultured with rat HIBD tissue extracts.
In conclusion, HBO promotes differentiation of NSCs into neurons and oligodendrocytes and reduced the number of astrocytes in vitro, possibly through regulation of Wnt3/nuclear β-catenin and BMP2 signaling pathways. HBO may serve as a potential therapeutic strategy for treating HIE.
Footnotes
Authors’ Contributions
Chongfeng Chen conceived, designed and performed the experiments, conducted the data analyses and wrote the manuscript. Yujia Yang contributed to the conception of the study and helped to perform the analysis via constructive discussions. Yue Yao performed the data analyses. All authors have read and approved the final manuscript.
Availability of Data and Materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethical Approval
All animals in the present study were purchased from the Animal department, XiangYa School of Medicine, and the experimental protocols were evaluated and approved by the Animal ethics committee of XiangYa School of Medicine, Central South University (Changsha, China).
Statement of Human and Animal Rights
All animals in the present study were purchased from the Animal department, XiangYa School of Medicine, and the experimental protocols were evaluated and approved by the Animal ethics committee of XiangYa School of Medicine, Central South University (Changsha, China).
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Natural science foundation of guangdong Province, China (grant no. S2013040012302).