
Abstract
Alpha-calcitonin gene-related peptide (αCGRP) plays a significant pathophysiological role in the regulation of bone metabolism. Our previous research indicated that αCGRP might have a potential application in enhancing osseointegration in vivo. To further uncover the intrinsic mechanism of its networks in bone regeneration, here we investigate the impact of αCGRP on osteogenic differentiation in bone marrow-derived mesenchymal stem cells (BMSCs) from both wild-type and αCGRP-/- mice. Considering the half-life of αCGRP in plasma is only 10 min, we applied αCGRP lentivirus and stably transfected it into BMSCs, followed by transfection identification and cell cycle assay. We further conducted a series of in vitro tests, and the results revealed that biological functions including migratory ability and osteogenicity exhibited positive correlation with BMSCs’ αCGRP expression. Meanwhile, this phenomenon was associated with an enhanced expression of YAP (Yes-associated protein), the key downstream effector of the Hippo pathway. To sum up, our data together with previous in vivo observations is likely to elucidate the intrinsic mechanism of αCGRP in bone remodeling, and αCGRP would appear to be a novel treatment to promote bone wound healing.
Keywords
Introduction
Although bone has the capability to completely repair and regenerate after simple fracture, there are a number of clinical situations in which auto- or allografts augment bone regeneration 1 . The issue of bone repair and regeneration has been the subject of heated discussion. It has recently been suggested that bone regeneration involves several partially overlapping phases 2 . However, the underlying problem with current techniques is that they often promote bone regeneration through direct osteogenesis 1 , which limits its further development.
Generally, cells like bone marrow-derived mesenchymal stem cells (BMSCs), osteoclasts, and osteoblasts are involved in bone metabolism. Recently, several studies have reported that osteoporosis is mainly attributed to the decreased osteogenic differentiation of BMSCs 1,3 , strengthening the importance of BMSCs and making them attractive candidates for clinical applications for bone remodeling. Moreover, as the common precursors for both osteoblasts and adipocytes 4 , it is critical to stimulate BMSC proliferation and differentiation into osteoblasts, thus improving therapy efficiency of bone wound healing. Later related studies have demonstrated that calcitonin gene-related peptide (CGRP), a 37-amino acid peptide, is able to stimulate the differentiation of BMSCs into osteoblasts in vitro 5 –9 .
CGRP is an important neuropeptide expressed in the regulation of bone metabolism, the cardiovascular system, the gastrointestinal system, and pain 10 –12 . There are two forms of CGRP: α- and βCGRP 13 . Traditionally, it is considered that αCGRP is a physiologic activator of bone formation, whereas βCGRP has little effect on this process 14,15 . Bone formation and osteopenia was downregulated in αCGRP-/- mice 16,17 . Our previous experiments suggested that αCGRP plays a significant pathophysiological role in bone development, metabolism, and remodeling around titanium implants via lentiviral transfection in vivo 17 .
The Hippo signaling pathway plays a critical role in multiple biological behaviors, such as organ development and tissue repair. Yes-associated protein (YAP) is the main transcriptional regulator in this pathway. When the Hippo pathway is activated, YAP is phosphorylated, resulting in its inhibition. Otherwise, YAP enters into the nucleus and activates downstream transcription. Yu et al. found in their study that calcitonin receptor-like receptor (CLR), a component of the Hippo signaling pathway receptor, prevents YAP phosphorylation; Therefore, CLR might promote YAP function during bone metabolism 18 .
We hypothesized that αCGRP plays a comprehensive role during osseointegration or a series of events involved in bone metabolism via coordinating various aspects. In addition, we tried to clarify the intrinsic mechanism regarding these events. In this study, we first investigated the impact of lentiviral vector overexpression αCGRP on osteogenic differentiation in BMSCs from both wild-type and αCGRP-/- mice, in order to determine the osteogenic effects of αCGRP, and further demonstrated its comprehensive intrinsic mechanism in bone regeneration.
Materials and Methods
Isolation and Culture of BMSCs
This study was approved by the Ethics Committee of West China Hospital of Stomatology, Sichuan University (No.WCHSRIB-D-2017-053). αCGRP+/+ mice (wild-type model) and αCGRP-/- mice (αCGRP-/- model) with the same background were purchased from RIKEN BioResource Center (Tokyo, Japan). Femoral and tibial bones were collected from both between 8 and 12 weeks of age and washed with phosphate-buffered saline (PBS). Then the femoral and tibial medullary cavities were flushed with α-MEM (Gibco, Grand Island, NY, USA) supplied with 10% fetal bovine serum (FBS; Gibco), 100 U/ml penicillin and 100 μg/ml streptomycin sulfate. The marrow cell suspension was aspirated repeatedly and filtered through a 70-µm cell strainer prior to culture in an incubator at 37°C with 5% CO2. Cell culture was continued with medium changes every 2 days until the cells were subconfluent. BMSCs between passages 2 and 4 were used for the following experiments.
Identification of BMSCs
Cells were identified with alizarin red staining for osteogenic differentiation and Oil Red O staining for adipogenic differentiation. To induce osteogenic differentiation, BMSCs were grown in osteogenic-inducing medium α-MEM containing 10% FBS, 100 U/ml penicillin, 100 μg/ml streptomycin sulfate, 50 μg/ml ascorbic acid (Sigma, St. Louis, MO, USA), 5 mM β-glycerol phosphate (Sigma) and 10-8 M dexamethason (Sigma) for 21 days. After that, cells were fixed in 4% paraformaldehyde for 30 min, followed by two washes with ice-cold PBS, were stained for 30 min with alizarin red, and then underwent two washes as previously described. The secreted osteocalcin level was then explored. To induce adipogenic differentiation, cells were cultured in the adipogenic differentiation medium, consisting of α-MEM supplemented with 10% FBS, 1 μmol/l dexamethasone, 0.05 mmol/l 3-isobutyl-1-methylxanthine, 10 μmol/ml bovine insulin and 200 μmol/l indometacin for 7 days. Oil Red O staining was performed to detect the lipid droplets. Briefly, cultured BMSCs were fixed in 4% paraformaldehyde for 30 min and washed with 60% isopropanol. They were stained with Oil Red O for 5 min, they then underwent a rinse with 60% isopropanol and two further rinses with ice-cold PBS.
Construction and Transfection of αCGRP Overexpression Lentiviral Vector
Our previous experiments have successfully constructed an αCGRP overexpression lentiviral vector system as previously described 17 . αCGRP+/+ BMSCs (wild-type BMSCs) and αCGRP-/- BMSCs (αCGRP-/- BMSCs) were divided into three groups. Wild-type BMSCs were allocated to three groups: (1) Wild-type group: αCGRP+/+ BMSCs without transfection; (2) Overexpression group: αCGRP+/+ BMSCs with αCGRP overexpression lentiviral vector transfection; and (3) Empty vector group: αCGRP+/+ BMSCs with αCGRP empty lentiviral vector transfection. In the same way, αCGRP-/- BMSCs were divided into αCGRP-/- group, overexpression group and empty vector group.
Empty vector group: a 40 μl quantity of αCGRP empty lentiviral vector containing GFP was added to 1 ml α-MEM and transfected into both wild-type BMSCs and αCGRP-/- BMSCs. Cells were cultured in an incubator at 37°C with 5% CO2 for 8–12 h.
αCGRP overexpression group: wild-type and αCGRP-/- cells were transfected with 40 μl αCGRP overexpression lentiviral vector via different multiplicity of infection (MOI) levels (10, 20, 40, 60, 80, and 100) and cultured as described above.
Untransfected cells were cultured as usual.
Identification of αCGRP Lentiviral Vector Transfection In Vitro
To further evaluate the expression of αCGRP via different MOI levels (10, 20, 40, 60, 80, and 100), western blot was performed. Briefly, cells of each group were washed three times with ice-cold PBS and then whole-cell protein extracts were obtained using lysis buffer (Keygen total protein extraction kit; Keygen Biotech, Nanjing, China) at 4°C for 30 min. This was followed by a 15-min centrifugation at 14,000 rpm at 4°C to obtain the supernatant; the bicinchoninic acid method was used as the quantitative assay. After boiling for 5 min, 50 μg protein of each group was applied to SDS-PAGE (10% polyacrylamide gel) at 80 V for 40 min and 100 V for 80 min. The separated proteins were then transferred in the gel to a polyvinylidene difluoride (PVDF) membrane (Millipore, Billerica, MA, USA) and blocked at room temperature for 1 h. After that, the membranes were probed with αCGRP antibody (1:1000, Abcam, Cambridge, UK) at 4°C overnight, followed by incubation for 1 h with horseradish peroxidase-conjugated secondary antibody (diluted 1:2000). Reactive bands were visualized using an enhanced chemiluminescence kit (Millipore). The densities of the bands were computer analyzed by a densitometer (Quantity One, Bio-Rad, Hercules, CA, USA).
Cell Cycle Assay
To confirm the effect and safety of αCGRP lentiviral vector transfection on wild-type and αCGRP-/- BMSCs, cell cycle was assessed by flow cytometry. Cells from different groups were collected and then centrifuged twice at 2,000 rpm for 5 min. Following fixation with 70% (w/v) ice-cold ethanol at 4°C overnight cells were washed with PBS, and there followed 8 min centrifugation at 1,000 rpm. To stain the cells, 100 ml of RNAase A (Keygen Cell Cycle Detection Kit, Keygen Biotech) was added at 37°C for 30 min, followed by incubation with 400 ml of propidium iodide (Keygen Cell Cycle Detection Kit, Keygen Biotech) at 4°C for 30 min. After that, flow cytometry (Beckman Coulter, Indianapolis, IN, USA) was conducted for each sample. The S-phase fraction of total cells (SPF) in each sample was calculated according to the formula:

The proliferation index (PI) was calculated according to the formula:

Cell Migration Assay
Migration of BMSCs was analyzed using transwell chambers. The pore size of the transwell membrane was 8 μm. A volume of 600 μl α-MEM containing 10% FBS was added to the lower chambers. Cells were plated into the upper chambers at the density of 2×104 cells/well in 200 μl α-MEM supplied with 1% FBS. The chambers were incubated at 37°C in 5% CO2 for 12 h and 24 h. After a wash with PBS, BMSCs that migrated to the lower face of the porous membrane were fixed with 500 μl cold methanol for 20 min at 37°C and stained with 500 μl 2 μmol/l Calcein-AM for 30 min in an incubator. Unmigrated BMSCs were removed with cotton swabs. Migrated BMSCs were photographed in five random fields per filter and measured by cell counting.
Immunofluorescent Staining
The effects of CGRP on expression of osteogenic phenotypes (alkaline phosphatase, ALP; collagen Type 1, COL-1) in BMSCs were examined. Cells in different groups were plated on coverslips in 6-well plates at the density of 1×104 cells/well until confluent, then the medium was removed. Cells were fixed with 4% paraformaldehyde for 30 min and permeated with 0.25% Triton X-100 for 8 min at room temperature. After incubation with 1% bovine serum albumin (Sigma) for 30 min, the cells were incubated with primary antibodies raised against ALP (diluted 1:50; Santa Cruz Biotechnology, Dallas, TX, USA) and COL-1 (diluted 1:100; Abcam) at 4°C overnight. Control slides were stained with only the secondary antibody. Following three washes with PBS, cells were incubated with fluorescein isothiocyanate-conjugated secondary antibodies (diluted 1:100; Zsjq Bio Co., Beijing, China) for 1 h. Nuclei were further stained with DAPI (diluted 1:100; Beyotime, Shanghai, China). The cells were then observed using fluorescence microscopy (Olympus, Tokyo, Japan).
Alizarin Red Staining
Alizarin red staining was conducted as the method described previously.
RNA Isolation and Real-time PCR Analysis
At 4, 7, and 14 days, total RNA from each group was extracted using Trizol Reagent (Invitrogen), according to the manufacturer’s instructions. The RNA was treated with DNase, and cDNA was synthesized using PrimeScript Reverse Transcriptase (Takara Bio Inc., Kusatsu, Japan). The detailed procedure for real-time RT-PCR was described previously 17 . Briefly, real-time PCR was performed in a 20 μl reaction mixture, using an ABI PRISM 7300 Real-time PCR System (Applied Biosystems, Waltham, MA, USA). Calculations of relative gene expression levels were conducted according to the 2- △△ Ct method, and presented as fold increase relative to the control group. Primer sequences for ALP, COL-1, osteocalcin (OCN), and osteopontin (OPN) are listed in Table 1; glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as internal control.
Primers and Probes used for PCR.
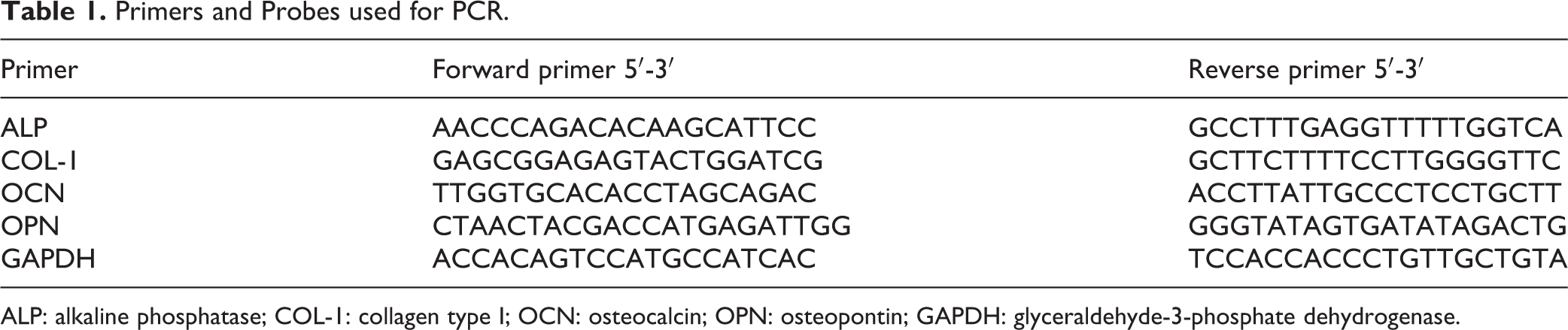
ALP: alkaline phosphatase; COL-1: collagen type I; OCN: osteocalcin; OPN: osteopontin; GAPDH: glyceraldehyde-3-phosphate dehydrogenase.
Western Blot Analysis
Western blot analysis was obtained as described previously. After blocking for 1 h in 0.1% Tween 20-Tris-buffered saline (TBST) containing 5% skim milk, membranes were incubated overnight at 4°C with anti-ALP (diluted 1:200; Santa Cruz Biotechnology), anti-COL-1 (diluted 1:500; Abcam, Cambridge, UK), anti-OCN (diluted 1:200; Santa Cruz Biotechnology), anti-OPN (diluted 1:1,000; Abcam), anti-YAP (diluted 1:1,000; Abcam) and GAPDH antibody (diluted 1:200; Zsjq Bio Co., China). Following three washes with TBST, membranes were incubated for 1 h with horseradish peroxidase-conjugated secondary antibody (diluted 1:2,000; Zsjq Bio Co., Beijing, China). Reactive bands were visualized using an enhanced chemiluminescence kit (Millipore). The densities of the bands were computer analyzed by a densitometer (Quantity One, Bio-Rad).
Statistical Analysis
Data are presented as mean ± SD. Statistical analysis was performed with SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA). Statistically significant differences were assessed by one-way analysis of variance (ANOVA) and Newman–Keuls post hoc tests. The value of p<0.05 was considered to be statistically significant.
Results
Identification and Expression of αCGRP via Different MOI Levels
Western blot was performed to evaluate the expression of αCGRP via different MOI levels (10, 20, 40, 60, 80, and 100) in wild-type BMSCs. Cells were allocated to the αCGRP+/+ group, overexpression group, and empty vector control group. As shown in Fig. 1A, αCGRP expression levels were almost undetectable under MOI 10, 20, and 40. With an increase to MOI 60, 80, and 100, αCGRP expression increased significantly in cells transfected with αCGRP overexpression lentiviral vector, while in the other two groups, αCGRP expression remained undetectable. MOI 100 showed the strongest target-gene expression in the overexpression group. The same test was performed in αCGRP-/- BMSCs; αCGRP expression did not rise obviously until MOI 40, 60, 80, and 100 in the overexpression group (Fig. 1B). According to the results, transfection via MOI 60 and MOI 40 might be the optimal condition in wild-type and αCGRP-/- BMSCs, respectively. In addition, the empty vector had no significant effects on those cells.

Western blot analysis of αCGRP expression via different MOI levels. (A) Wild-type BMSCs. (B) αCGRP-/- BMSCs.
Cell Cycle Assay
Flow cytometry was monitored to examine the effects of transfection on the cell cycle at MOI 60 of αCGRP+/+ BMSCs (Fig. 2A–C) and MOI 40 on αCGRP-/- BMSCs (Fig. 2D–F). The SPF and PI values in each sample were calculated according to the formula. There was no significant difference between the αCGRP+/+ group, overexpression group, and empty vector control group (p>0.05), thus transfection via MOI 60 had no adverse effect on αCGRP+/+ BMSCs’ cell cycle compared with the other two groups (Fig. 2A–C), as was the case also with the αCGRP-/- group, overexpression group, and empty vector control group via MOI 40 transfection (Fig. 2D–F). Taken together, MOI 60 and MOI 40 confirmed both the transfection efficiency and safety in wild-type and αCGRP-/- BMSCs, respectively.

Results of cell cycle assay. (A and D) The SPF parameter in each group. (B and E) The PI value in each group. (C and F) Proliferation activity was reflected by the cell cycle distribution in the logarithmic growth phase.
αCGRP Promotes BMSC Migration
Based on the above information, the empty vector had no significant effects on BMSCs. The cells were divided into four groups with different expression levels of αCGRP: (1) αCGRP-/- control group: Untransfected αCGRP-/- BMSCs; (2) αCGRP-/- recovery group: αCGRP-/- BMSCs+αCGRP transfection; (3) αCGRP+/+ control group: Untransfected wild-type BMSCs; (4) αCGRP+/+ overexpression group: wild-type BMSCs+αCGRP transfection. Western blot analysis indicated that the expression levels of αCGRP were different in the four groups (Fig. 3A), which demonstrated that they could be compared in the following assays.

Results of Transwell assays. (A) Western blot analysis. (B) The migratory response of BMSCs at 12 h and 24 h in each group. (C) Quantitation of the migratory response of BMSCs. Scale Bar=50 μm. *p<0.05.
Transwell assays were conducted to observe the migratory response of BMSCs to αCGRP. We noted that BMSCs could be found in the lower chambers at 12 h in all groups, although these cells did not have characteristic morphology until 24 h (Fig. 3B). As demonstrated in Fig. 3C, the migration of the αCGRP+/+ overexpression group was much higher than the αCGRP-/- control group (p<0.05). Most cells in the αCGRP-/- control group had not even reached the underside of transwell chambers at 24 h.
Immunofluorescent Staining
The expression of the osteoblastic markers ALP and COL-1 was assessed by immunofluorescent staining at day 7. As shown in Fig. 4, cells in the αCGRP+/+ overexpression group displayed obviously increased expression of ALP in the cytoplasm. The expression of COL-1 was similar to ALP. With the increase of αCGRP expression levels in BMSCs, COL-1 expression level became more obvious (Fig. 5).
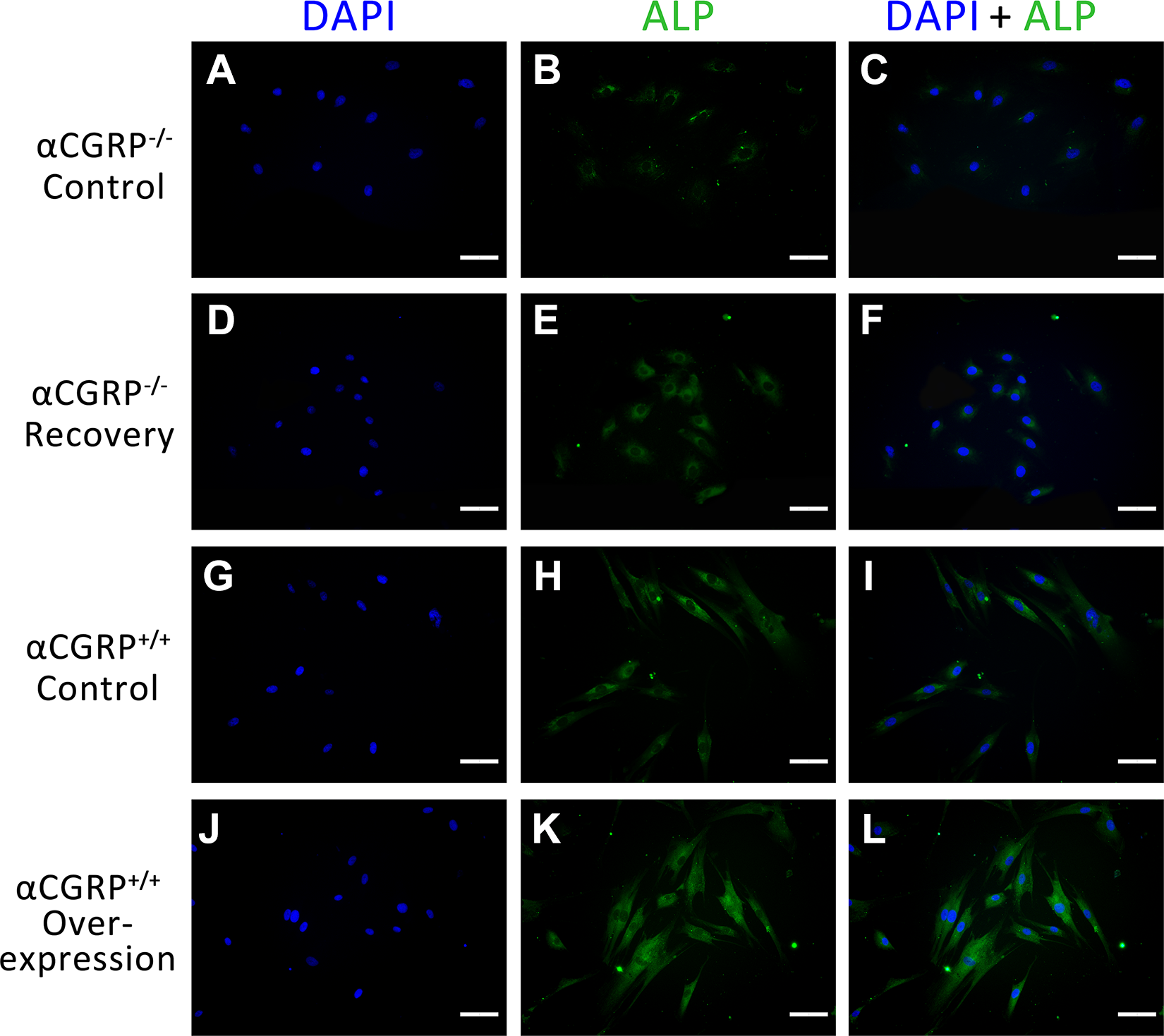
Immunofluorescent staining of ALP. Scale Bar=50 μm.

Immunofluorescent staining of COL-1 in each group. Scale Bar=50 μm.
Alizarin Red Staining
Alizarin red staining was conducted to further confirm the effects of αCGRP on osteogenic terminal differentiation in BMSCs. From Fig. 6A–D, we could clearly observe that alizarin red-positive nodules appeared more evenly distributed in the αCGRP+/+ overexpression group. The results also indicated that compared with the αCGRP+/+ control group and αCGRP+/+ overexpression group, αCGRP-/- control groups showed a dramatically decreased mineral nodule formation, which was in accordance with the quantitative analysis in Fig. 6E (p<0.05). On the other hand, although the regions of calcified nodule in the recovery group were more apparent than those in αCGRP-/- control group, there was no statistical significant difference (p>0.05).

Alizarin red staining of BMSCs. (A–D) Microscopy observation of alizarin red-positive mineral nodules in the αCGRP-/- control group (A), αCGRP-/- recovery group (B), αCGRP+/+ control group (C), and αCGRP+/+ overexpression group (D), respectively. (E) Quantitation of the density of alizarin red-positive nodules. Scale Bar=500 μm. *p<0.05.
Osteoblast-associated Phenotype Expressions
The gene expression levels of ALP, COL-1, OCN and OPN were examined with real-time PCR at 4, 7, and 14 days, in order to evaluate the condition of the early and terminal osteogenic differentiation. As revealed in Fig. 7A, compared with the αCGRP+/+ control group and αCGRP-/- recovery group, ALP gene expression was downregulated in the αCGRP-/- control group at 7 days (p<0.05). At the same time, COL-1 gene expression increased during the entire observation period, although that trend did not reach significant statistical difference (p>0.05) (Fig. 7B). αCGRP also enhanced the expression level of OCN (Fig. 7C), and they were positively correlated at day 14 (p<0.05). In terms of OPN, the αCGRP-/- recovery group presented a noticeable elevation compared with the αCGRP-/- control group at day 7 and tended to be close to the αCGRP+/+ control group (Fig. 7D). The peak OPN mRNA expression level in the αCGRP+/+ overexpression group at day 14 was about 2.2 times that of the αCGRP-/- control group. Briefly, the expression levels of osteoblast-associated markers ALP, COL-1, OCN, and OPN exhibited positive correlation with BMSCs’ αCGRP expression.

Osteoblast-associated phenotype expressions. Levels of (A) ALP, (B) COL-1,(C) OCN, and (D) OPN. (E) Western blot analysis. All data are expressed as mean±SD from three independent experiments. *p<0.05.
We further investigated the effect of αCGRP on protein expression of the osteogenic phenotypes. The data (Fig. 7E) suggested that the expression of osteogenic phenotypes was downregulated dramatically in the αCGRP-/- control group compared with the other three groups, including a decrease of ALP, COL-1, OCN, and OPN. αCGRP+/+cells transfected with lentiviral vector containing αCGRP showed increased expression of osteogenic phenotypes, especially a noticeable elevation of OCN. Due to the stable transfection of αCGRP, the αCGRP-/- recovery group exhibited more ALP, COL-1, and OCN protein expression compared with αCGRP-/- control groups and almost reached the same expression level as the αCGRP+/+ control group. Further investigation revealed that the enhanced expression levels of osteoblast-associated markers were associated with an increase of YAP expression, which is the significant effector of the Hippo pathway.
Discussion
Skeleton is a living and continuously remodeling tissue, which exhibits abundant sensory neuron innervations 19,20 . During bone wound healing, CGRP and other neuropeptide substances are secreted by nerve endings in the bony callus, promoting bone metabolism and reconstruction 21 . Originally, αCGRP was shown to be a highly potent vasodilator 22 . Later, related studies suggested that αCGRP was found to be upregulated in regenerating axons within injury zones 23 –25 and capable of promoting proliferation of local Schwann cells 26 , which is critical for partnering during peripheral nerve regeneration 27 . Other researchers have also reported that αCGRP is a physiologic activator of bone formation 14,15 . Moreover, research shows that CGRP is able to promote inflammation and inhibit inflammation mediator release through its ability to influence the activity of inflammatory cells, including lymphocytes, Langerhans cells and macrophages 28 –32 . Based on these findings, our team hypothesized that αCGRP may be a novel treatment to promote the reconstruction of neural feedback pathways and microcirculation during the bone healing process, thus improving osseointegration or other events during bone wound healing 27 .
Our previous experiments have suggested that implant osseointegration significantly decreased in the αCGRP
In this study, we applied an αCGRP overexpression lentiviral vector system and stably transfected it into BMSCs from both wild-type and αCGRP-/- mice. As expected, this gene delivery system effectively promoted αCGRP expression in the target cells. Flow cytometry and western blot showed that transfection via MOI 60 and 40 might be an optimal condition not only possessing basic transfection efficiency, but also having negligible adverse effects on proliferation of αCGRP+/+ and αCGRP-/- BMSCs. In addition, no obvious differences in αCGRP expression were observed between cells untransfected or transfected with empty vector, which indicates that transfection with the lentiviral vector alone does not change αCGRP expression in BMSCs. Taken together, we successfully assessed different groups with different αCGRP expressions; thus we explored the effect of αCGRP expression levels on BMSCs and the potent molecular mechanisms.
Experimental evidence indicated that BMSCs can directly migrate to an injured site to proliferate and differentiate into osteoblasts 33 . More importantly, promoting BMSCs to migrate effectively to an injured site has been a difficult problem to solve 34,35 . Thus, we carried out transwell assays, which indicated that the overexpression of αCGRP in BMSCs can increase cell motility, stimulating the BMSCs’ migratory response in the early stage. In addition, ALP activity indicated that the αCGRP+/+ overexpression group showed the highest osteogenic ability among the groups at 7 days. Furthermore, more and larger mineralized nodules formed in the αCGRP transfected group compared with the control group. αCGRP transfection also stimulated expression of osteogenic phenotypes, including an increase of ALP, COL-1, OCN, and OPN. Meanwhile, similar phenomena were observed in western blot analysis. Moreover, the αCGRP-/- recovery group exhibited increased functional osteogenic ability compared with the αCGRP-/- control group and almost reached the same level as the αCGRP+/+ control group. Collectively, we have demonstrated that αCGRP plays a prominent role in regulating bone remodeling.
It has been suggested that CGRP could stimulate proliferation, growth, and differentiation of cells involved in bone metabolism via receptors, producing a variety of biological effects 36 –40 . A functional CGRP receptor consists of at least three proteins: calcitonin receptor-like receptor (CLR), single transmembrane-spanning receptor activity modifying protein 1 (RAMP1), and receptor component protein (RCP) 41,42 . The latest related studies suggested that the Wnt/β-catenin signaling pathway was involved in CGRP-mediated osteoblastic differentiation of BMSCs 43 ; others have indicated that CGRP-induced mouse BMSCs’ osteogenesis was associated with the Hippo pathway 44 . Yu et al. mentioned in their study that Hippo-YAP signaling was a downstream branch of G-protein coupled receptors (GPCR) 18 . Meanwhile, as the CGRP receptor belongs to GPCR, and it was observed that one of the receptor components CLR prevents YAP phosphorylation, therefore, CLR might promote YAP function during bone metabolism 18 . In this study, we explored whether CGRP-mediated BMSCs’ migration and osteogenesis were related to the Hippo-YAP pathway. The results indicated that the increased migration ability and osteogenic phenotype expressions were associated with an enhanced expression of YAP, the significant effector of the Hippo pathway. To sum up, our investigations indicate that the Hippo-YAP pathway may contribute to αCGRP-mediated BMSCs’ migration and osteogenesis. However, the intrinsic mechanism regarding this is still unclear and it has remained elusive whether other signaling pathways are involved.
In the meantime, it should be noted that besides the above-mentioned effects, we could not exclude other effects on bone remodeling caused by αCGRP. It may affect BMSCs’ and other bone-related cells’ angiogenesis, neuroregulation, inflammatory reaction, and so forth. A related study reported that CGRP signaling maintains bone mass both by directly stimulating stromal cell osteoblastic differentiation and by inhibiting RANKL-induced NF-κB activation, osteoclastogenesis, and bone resorption 15 . Others have also suggested that CGRP can promote the migration of BMSCs and stimulate the protein expression of vascular endothelial growth factor, and the release of cytokines and growth factors from other cell types; thus enhanced bone formation may simply be attributed to the known angiogenic and vasodilatory effects of CGRP 45 . To further uncover the molecular mechanism of αCGRP net effects in bone regeneration, we will conduct further studies in the next step.
Conclusion
In conclusion, αCGRP-mediated BMSCs’ migration and osteogenesis might relate to its downstream branch Hippo-YAP pathway. Our data together with previous results from animal models and further analysis is likely to elucidate the intrinsic mechanism of αCGRP in bone remodeling, and αCGRP would appear to be a novel treatment to promote osseointegration and other related events involved in bone metabolism.
Footnotes
Author Contributions
Bin Wang and Jie Lin carried out the experiments and analyzed results. Bin Wang and Qin Zhang drafted the manuscript. Ping Gong participated in the design of the study and preparation of the manuscript. Xinyuan Zhang and Hui Yu participated in the analysis and interpretation of data. Lin Xiang supervise of all the studies. All authors read and approved the final manuscript.
Ethical Approval
Ethical approval to report this study was obtained from Ethics Committee of West China Hospital of Stomatology, Sichuan University (No.WCHSRIB-D-2017-053).
Statement of Human and Animal Rights
All procedures in this study were conducted in accordance with approved protocols of the Ethics Committee of West China Hospital of Stomatology, Sichuan University.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the National Natural Science Foundation of China (No.81701007), Sichuan Science and Technology Program (No. 2018RZ0087), the Fundamental Research Funds for the Central Universities (No. 2018SCUH0006) China Postdoctoral Science Foundation Grant (No. 2018M631091), and the Basic and Applied Basic Research Projects of West China Hospital of Stomatology of Sichuan University (RD-02-201902).