
Abstract
The dorsal root entry zone is often used in research to examine the disconnection between the central and peripheral parts of the nervous system which occurs following injury. Our laboratory and others have used transplantation of olfactory ensheathing cells (OECs) to repair experimental spinal cord injuries. We have previously used a four dorsal root (C6–T1) transection model to show that transplantation of OECs can reinstate rat forelimb proprioception in a climbing task. Until now, however, we have not looked in detail at the anatomical interaction between OECs and the peripheral/central nervous system regions which form the transitional zone. In this study, we compared short- and long-term OEC survival and their interaction with the surrounding dorsal root tissue. We reveal how transplanted OECs orient toward the spinal cord and allow newly formed axons to travel across into the dorsal horn of the spinal cord. Reconstruction of the dorsal root entry zone was supported by OEC ensheathment of axons at the injured site and also at around 3 mm further away at the dorsal root ganglion. Quantitative analysis revealed no observable difference in dorsal column axonal loss between transplanted and control groups of rats.
Introduction
Sensory neurons within the olfactory system have a limited lifespan and are continually replaced throughout life 1,2 . The pathway along which new olfactory axons grow is formed by olfactory ensheathing cells (OECs) 3,4 . This ability to act as a conduit for growing axons is one of the reasons which led to their use in the repair of central nervous system (CNS) injuries. Transplantation of cultured cells from the olfactory bulb has been shown to induce axonal regeneration and restore function in a number of experimental models of spinal cord injury 5 –9 .
Evidence from a number of laboratories has highlighted the advantages of OEC transplantation: modulation of the growth-inhibitory glial scar, long-distance axonal regeneration, axonal myelination, removal of cell debris, or attenuation of the immune response are just some of the findings that have been reported 10 –13 .
We have previously used a rat dorsal root injury model to assess the return of upper limb proprioception after OEC transplantation. Electrophysiological recordings showed that animals with reinstated proprioception also had improved synaptic transmission from the periphery to the cuneate nucleus 5 . In this study and our previous studies, we observed OECs acting as a bridge to allow regenerating dorsal root axons to cross the glia–pial surface and then ascend in the myelinated tracts of the dorsal columns 14 . Until now, however, the precise mechanism underlying repair of injured dorsal roots is not fully known.
In this current study we have investigated the anatomical changes associated with OEC transplantation after dorsal root injury. We performed a histological examination by immunostaining methods of the dorsal root entry zone at 1 week and 6 weeks following injury to see how the transplanted cells interacted with the peripheral and central compartments of the transitional zone. Axonal tracing and quantitative analysis of axon loss within the adjacent dorsal horn and dorsal columns was also carried out.
Materials and Methods
All animals used according to the UK Home Office regulations for the care and use of laboratory animals, the UK Animals (Scientific Procedures) Act 1986 with the ethical approval of the University College London Institute of Neurology.
Cell Culture
A detailed description of the cell culture protocol is given elsewhere 15 . Briefly, the nerve and glomerular layers of the olfactory bulbs from adult female Albino Swiss rats (AS, locally bred, London, UK, body weight 200–220 g) were dissected out, dissociated using 0.1% trypsin (Thermo Fisher Scientific, Grand Island, NY, USA), and cultured in a medium consisting of DMEM/F-12 and 10% fetal bovine serum supplemented with penicillin-streptomycin (100 U, 100 µg/ml; all from Invitrogen, Paisley, Scotland, UK). The cultures were monitored until they reached confluence (at 14–16 days) where OECs and olfactory nerve fibroblasts were seen in roughly equal numbers and were embedded within an endogenous matrix. These cells were infected by lentivirus with the green fluorescent protein (GFP) transgene 3 days before transplantation.
Surgery
As described in detail elsewhere 5 , hemilaminectomies from C4–T2 were carried out on 18 AS rats, body weight 200–220 g under isoflurane (Abbott Laboratories Ltd., Berkshire, UK) with the aid of an operating microscope to expose the spinal cord. Through a posterior surgical approach, an incision was made through the dorsal midline, the paraspinal muscles were separated, and the spinous process of the second thoracic vertebra (T2) was located. Following the hemilaminectomy and identification of segmental levels, microscissors were used to transect dorsal roots C6, C7, C8, and T1 close to the spinal cord.
Roots injury alone (n=6) and injury with transplantation of OECs (n=7) for immunohistochemistry
The matrix containing cultured OECs expressing GFP was scraped from a Petri dish, applied between the cut ends of dorsal roots and their original entry points into the spinal cord, and retained in place with fibrin glue (Tisseel Kit, Baxter, Thetford, UK). Six rats received the injury alone, i.e., without the OEC transplant and were used as controls.
Roots injury with transplantation of OECs for BDA tracing study (n=5)
A further series of five rhizotomized rats with OEC transplants were used for tracing sensory afferents by injection of biotin dextran amine (BDA) into the ganglia. To avoid tracer passing through anastomoses to adjacent intact roots, extended laminectomies were carried out and then six roots from C5 to T2 were sectioned. The four roots from C6 to T1 were transplanted with OECs. Then 4 weeks after transplantation, about 0.5 µl of 10% BDA in saline was injected by micropipette at three points into each of the two middle (C7 and C8) DR ganglia. Two weeks later, the rats were perfused and fixed with 4% phosphate-buffered paraformaldehyde.
Tissue Processing and Immunohistochemistry
The rats were terminated under deep carbon dioxide anesthesia and perfused transcardially with 50 ml 0.1 M phosphate buffer solution followed by 400 ml of 4% paraformaldehyde buffered with phosphate solution for around 25 min. The vertebral columns were dissected out from the craniocervical junction to the upper thoracic level and left to harden in the same fixative for 48 h 16 .
Under a dissecting microscope, the spinal cord and the associated roots were carefully dissected from the bony skeleton while preserving continuity across dorsal roots and transplants to the spinal cord. For cryoprotection, tissues were then sequentially placed into 10% and 20% sucrose solution until sinking before they were frozen with CryoSpray (Cell Path, Newton, Wales, UK) and sections cut on a cryostat (Leica CM3050).
For double immunostaining, 16 µm sections were incubated in 2% milk containing 1:1000 anti-glial fibrillary acidic protein (GFAP) antibodies (ms, Millipore, Watford, UK) and 1:500 anti-laminin antibodies (rabbit polyclonal, Sigma, Hertfordshire, UK) overnight at 4°C. For neurofilament staining, anti-neurofilament 1:500 heavy chain and light chain polyclonal rabbit antibodies (68kD and 200kD, rb, Abcam, Cambridge, UK) were incubated overnight. Secondary antibodies were 1:500 anti-rabbit and anti-mouse (Alexafluor 546 or 488, Thermo Fisher Scientific), incubated for 1–2 h in the dark at room temperature. For detection of BDA labeling, 30 µm sections were incubated in 1:400 streptavidin conjugate (Alexafluor 546, Thermo Fisher Scientific) overnight at 4°C.
Loss of Dorsal Column Axons: Quantification Analysis (n=13)
To quantify the extent of axon loss within the ascending dorsal columns, the numbers of neurofilament-positive axon bundles in a region of interest (ROI) within the cuneate fasciculus of transverse sections at cervical level C7–C8 were counted. This was to determine whether OEC transplantation had an effect on the degree of axon loss caused by dorsal root rhizotomy.
Tissue from 13 rats was used for this analysis; seven were from the rats which had injury and received OEC transplants and six were “injury only,” and all had survived 6 weeks post-surgery. For each animal, cryostat sections at cervical level C7–C8 were immunostained as described above for neurofilament (light and heavy chains). Cervical levels C7–C8 were used because this was in the center of the C6–T1 dorsal root transection area, ensuring we did not analyze adjacent, uninjured segments. For the ROI, a square area of size 600 µm by 600 µm was imaged immediately dorsal to the medial part of the substantia gelatinosa (see Fig. 3A–D). For subsequent analysis, images were taken at this ROI on both the injured and non-injured sides of the spinal cord section.
The software ImageJ (National Institutes of Health, Bethesda, MD, USA) was used to count the number of axon bundles. First, ROI images from injured and non-injured sides of a tissue section were manually thresholded. Next, using the “Particle Analysis” function, two distinct size ranges were counted: small axon bundles with a diameter of 2–4 µm, and large bundles 4–10 µm. We used these size ranges in case Wallerian degeneration had a differential impact on different bundle sizes. This analysis was done by someone blind to whether the tissue came from Responder or Non-Responder groups.
Statistical Analysis
For the quantification of axon loss, results were expressed as axon number means ± SEM, with statistical comparison between groups made using a one-way analysis of variance (ANOVA), to determine F-ratio significance. Post hoc analysis was with Bonferroni multiple comparisons and IBM SPSS Statistics 22.0 software. Statistical significance was accepted at p<0.05.
Results
Short Term: OECs and their Early Interaction with the Dorsal Root Entry Zone
One week following injury to the dorsal roots, transplanted OECs were seen as a tightly packed mass of cells lying close to the entry zone. In transverse sections, around 40 GFP-labeled cells had entered the spinal cord and were found at around 200 µm from the transplantation site (see Fig. 1A, D).

Transplanted OECs retained at the entry zone one week after dorsal root rhizotomy (n=12). At low (1A) and high-power views (1B–F). High magnification shows GFP-labeled cells at the dorsal root entry zone interacted with GFAP+ and laminin+ (PNS) tissue following injury (1B, E). OECs intermingled with or ensheathed some neurofilament+ fibers (1C, D, F). Scale bars: 200 µm (A, B), 100 µm (C–F). Survival time 1 week. Transverse sections (A, B, D, E) and horizontal sections (C, F).
Higher magnification showed that the outer layer of the cluster of transplanted OECs had thick, elongated processes which were aligned at right angles to the surface of the spinal cord (asterisk in Fig. 1D). Closer to the entry zone, however, the processes of OECs were much shorter and appeared to “turn in” toward the CNS (arrowheads in Fig. 1D). Many neurofilament-positive fibers were found ensheathed by OECs in both the outer surface and inner core of the cell transplant. As with the OECs, short axonal fibers seen at the entry zone closest to the spinal cord were oriented toward and intermingled closely with the astrocytes of the CNS.
These GFAP+ astrocytes are shown in Fig. 1B, E where, particularly at the outermost edge of the spinal cord, a large number had ramified, fibrous processes which protruded outwards toward the periphery. At the injured transitional zone, much of the GFAP+ tissue overlapped with laminin-positive staining (arrowheads in Fig. 1E) but relatively little laminin-positive tissue was seen in the peripheral part of the entry zone.
In horizontal sections, transplanted OECs were found in close apposition to the spinal cord, with several parallel layers of OECs extending processes as far as 100 µm (Fig. 1C, F). A proportion of these OECs had ensheathed neurofilament-positive fibers which often ran in parallel to the spinal cord (in Fig. 1F) when they had entered the spinal cord or when they were aligned at right angles to the spinal cord at the entry zone (arrowheads in Fig. 1C).
Long Term (i): OECs act as a Bridge for Regenerating Axons at the Dorsal Root Entry Zone
Six weeks after injury, there was prominent survival of substantial numbers of transplanted cells at the dorsal root entry zone (Fig. 2A, C). Although a few of the OECs had migrated out of the dense transplant mass, most had remained close to their original point of transplantation. The majority of the OECs had elongated processes which spanned up to 100 µm in various directions at the injury site. There were around 6000 GFP-labeled cells in each transplant at 6 weeks after transplantation. Lentivirus labeling in the present study labeled around 20% of total cell population. Therefore the number of cells including the ones without GFP labeling was estimated around 30,000 in each transplant. The double immunostaining revealed integration of CNS tissue and peripheral nervous system (PNS) tissue at the transitional zone (Fig. 2B, D).
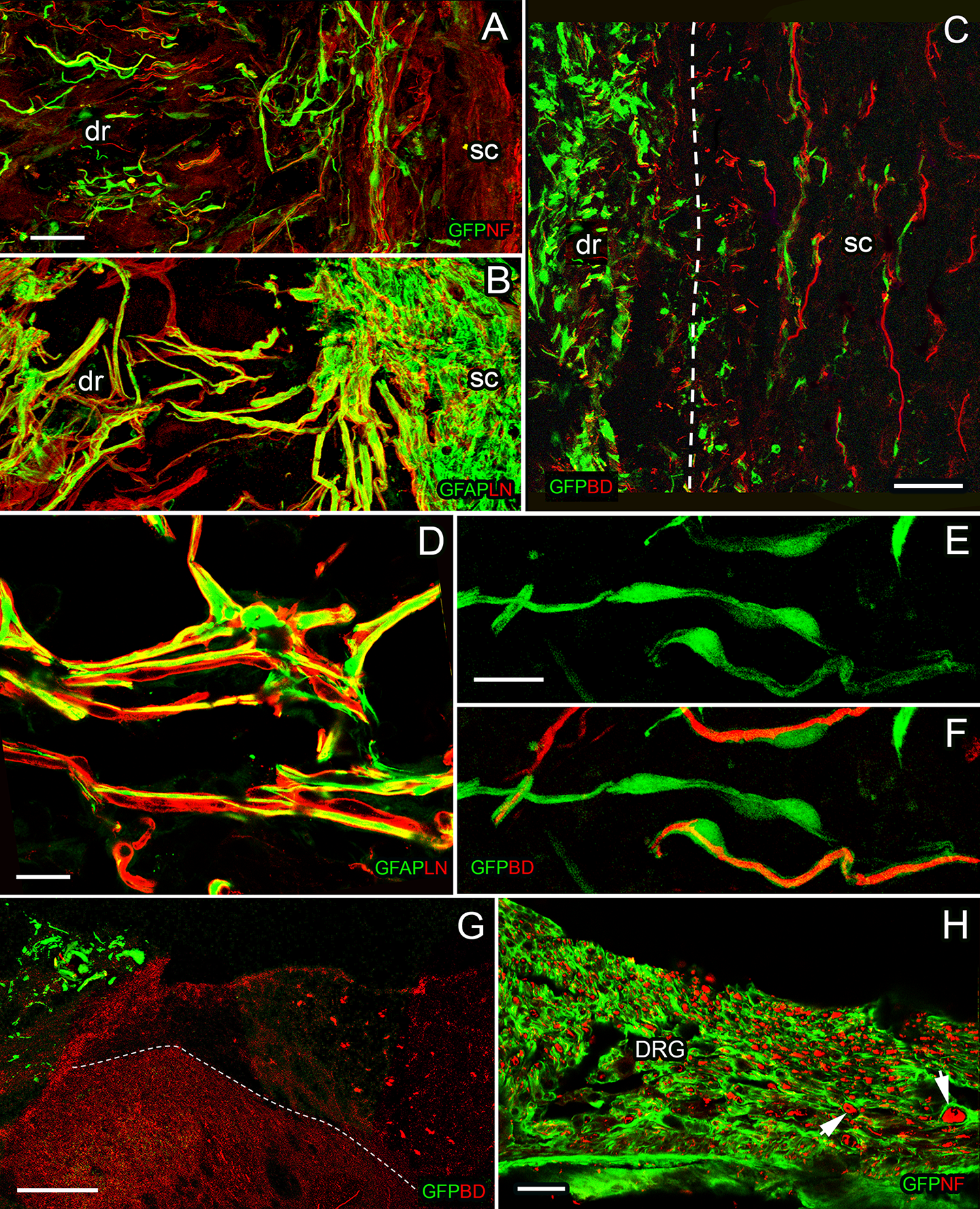
Transplanted OECs and BDA-labeled fibers at the entry zone 6 weeks after dorsal root rhizotomy (n=12). Images (Fig 2A, C) show GFP-labeled cells bridging between the peripheral and central parts of the transitional zone after injury. BDA-labeled fibers intermingled with and at times ensheathed by GFP-labeled OECs (Fig 2C, E, F) as they travel through to the injury site and enter the spinal cord (Fig 2C, dotted line to show the border between the dorsal root and the spinal cord). Migration and ensheathment of OECs in a dorsal root ganglion (Fig 2 H). High magnification of cross-section image (Fig 2G) showed GFP-labeled cells (green) retained at the entry zone and BDA-labeled regenerating fibers travel through to the dorsal column of the spinal cord (red, dotted line to show the border between the dorsal column and the dorsal horn). High-power images revealed a one-to-one ensheathment of BDA-labeled fibers (red) by transplanted OECs (E, F). Integration of CNS and PNS tissue at the transitional zone (Fig 2A, B, D). Neurofilament/GFP for Fig A, H; BD/GFP for C, E, F, G; GFAP/LN for B, D. Scale bars: 50 µm (A, B, H), 20 µm (D, E, F), 100 µm (G). Survival time, 6 weeks. Transverse section: 2G; horizontal sections: 2A, B, C, D, E, F, H.
In tissue from animals which received a BDA tracer injection and were perfused 6 weeks after surgery, extensive axonal regenerating axons had elongated across the damaged transitional zone. BDA-labeled fibers were found passing from the periphery and into the astrocytic compartment of the CNS (Fig. 2C, G). Some of these fibers which passed into the spinal cord were ensheathed by OECs, with most of these being small diameter axons (Fig. 2C, F).
High-magnification images revealed that there was a one-to-one ensheathment of BDA-positive axons by OECs. The elongated processes of the OEC had a central lumen containing a BDA-labeled regenerating axon. OECs did not often travel long distances within the spinal cord; most of them remained at the injured site, at the junction between what was the PNS–CNS border.
Long Term (ii): Migration of OECs and Regenerating Axons in the Dorsal Horn 6 Weeks Post-Injury
Six weeks after injury, some transplanted cells had migrated quite extensively. The transplanted cells migrated peripherally as far along the dorsal root as the dorsal root ganglia (up to 1.5–2 mm), as seen in Fig. 2H. GFP-labeled cells were found to intermingle with and ensheath many of the neurofilament-positive dorsal root ganglion (DRG) cells and their axons. They were also found to ensheath both small and large neurofilament-positive axons within the DRG, all apparently in the same one-to-one arrangement as seen at the 1 week post-injury time point.
Low-power images which outline large areas of the dorsal horn and dorsal columns revealed the route taken by regenerating axons after they crossed the dorsal root entry zone. Some axons took long meandering routes through the dorsal gray matter before appearing to terminate within the dorsal column (see Fig. 2G). Transverse sections showing cross-sections of numerous (up to 20) regenerated axon bundles were seen within this dorsal column white matter area. Some axons were bifurcated and may have terminated at more than one site. Axons were found within the cuneate fasciculus in a number of sections more rostral to the injury level, suggesting they traveled longitudinally to synapse at the cuneate nucleus.
Long Term (iii): Quantification of Dorsal Column Axon Loss 6 Weeks after Injury
As we saw a number of regenerating axons within the cuneate fasciculus, we went on to compare axon numbers in this region between control and transplanted groups at 6 weeks after injury. There was an obvious loss of axons within the dorsal columns at cervical level C7–C8 following the dorsal root injury. The regular array of tightly packed small and large axons seen on the normal side (Fig. 3A, B) contrasted with the numerous empty vacuole-like spaces seen at 6 weeks following injury (Fig. 3C, D). On the non-injured side of the spinal cord, we counted around 500 axon bundles (476 ± 78) within the dorsal column ROI. After injury, however, this number decreased by 43% to 271 (± 50) axon bundles. There was no significant difference between axon numbers found in the “injury-only” group and the transplanted group of rats.

Quantification of axon loss within the dorsal columns 6 weeks following rhizotomy injury (Control group, n=7; OEC group, n=6). Representative images showing the region of interest (ROI) on non-injured (A, B) and injured sides (C, D) of spinal cord sections. Comparison of axonal counts at the ROI on the non-injured and injured sides of the cervical level C7–C8 spinal cord in control and transplanted animals at 6 weeks survival time (E). Axon counts split into small (2–4 µm) and large (4–10 µm) range of axon bundle sizes. One-way ANOVA, F (3,22)=3.22 p<0.05. Post-hoc Bonferroni multiple comparisons revealed no statistically significant results. Scale bars: 500 µm (A, C), 200 µm (B, D).
We distinguished between small (2–4 µm) and large (4–10 µm) axon bundles to see whether Wallerian degeneration had a differential impact on axon bundles according to their diameter. Prior to injury, there were 102 (± 25) large axon bundles, but this number decreased by 42% to around 61 (± 20) axon bundles after rhizotomy. For the small axon bundles, the percentage decrease was very similar (44%) from pre-injury levels of 374 (± 78) to 210 (± 20) at 6 weeks after rhizotomy. There was no apparent difference in loss of small or large axon bundles between injury-only and OEC transplant groups.
Discussion
Here we have examined the reconstruction of the injured dorsal root entry zone following transplantation of OECs. Several of our previous studies have shown axon regeneration along with functional recovery of the forelimb after rat spinal dorsal root transection and OEC transplantation 5,6,15 . In this current study, we used an anatomical approach to investigate how transplanted cells repair the delicate interface between the peripheral and central spinal cord compartments. Using BDA labeling, we have shown that the reconstituted entry zone acts as a bridge to allow sensory axon regeneration into the dorsal horn and then on to ascend in the dorsal columns.
For the first time, we have found that many migrating OECs extensively ensheath cells of the adjacent DRG. This is not surprising because it is known that OECs myelinate DRG axons in co-culture studies, where they show a similar bundling and alignment of axons as seen in the olfactory system 17 . Given that the cell bodies of the sensory axons reside within the DRG itself, it is crucial that, following injury, as many of these cells as possible continue to receive growth factors to survive 18 –20 . The tight, one-to-one ensheathment of ganglion cells could protect or even amplify the conduction of afferent signals from the DRG through to the dorsal horn and up to the cuneate nucleus 21,22 . Our previous electrophysiological study revealed improved synaptic transmission in injured rats which had received an OEC transplant and then had recovered forelimb proprioception 16 . Therefore, the beneficial function of OECs could be due to this extensive ensheathment and the close association of OECs with DRG cells that we have described here.
By examining the reconstituted entry zone, we observed how transplanted OECs interacted with the delicate interface between astrocytes, axons, and peripheral nerve tissue to allow a bridge for sensory neurons to cross. The “intertwining, periodic ladder-like structure” formed between the outgrowing astrocytic branches and processes arising from peripheral nerve tissue is similar to what we described in a previous study with lumbar-level dorsal roots 14 . The special relationship which OECs have with astrocytes has been described before 23 –25 , and may be explained by their normal role within the olfactory bulb. For example, following removal of an olfactory bulb, neonatal axons have been shown to redistribute their innervation to other areas such as the hippocampus 26 . This would involve OECs opening the astrocytic glia–pial lamina which forms a barrier for the regenerating axons to enter the brain. It is likely that this function has enabled OECs to interact positively with the astrocytic border of the spinal cord in this current study—allowing entry of sensory axons into the spinal cord.
The microglia reside in the CNS, but they can be altered after spinal root injury. Using time-lapse imaging in an obstetrical brachial plexus injury model, a recent study from Green et al. 27 showed that after the spinal root injury, the microglia squeeze through the spinal boundary and emigrate to peripheral spinal roots. Although both macrophages and microglia respond, microglia are the debris-clearing cell 27 . In a recent study we investigated microglia response to the roots injury alone or with transplantation of OECs. We compared microglial activation within the dorsal horn and we noticed that there was little difference was seen between the two groups 28 . We will look at microglia interaction with PNS tissue at the transitional zone in future studies. A study from Chen et al. 29 found that microglial cells were activated in spinal cord dorsal horn after nerve injury. Their results provide the evidence that the activation of microglia in the spinal cord after nerve injury is unlikely to be due solely to neuronal activity, non-neuronal factors are likely responsible for the activation of nerve injury-related microglial cells in the spinal dorsal horn.
Close inspection of high-power images revealed that GFP-labeled cells in the innermost layer were oriented in the direction of “turning in” toward the spinal cord. They were associated with short axons which also had the same orientation directed toward the astrocytes. This arrangement is similar to that seen in mice at late embryonic stages, where glial cells located at the dorsal root entry zone are thought to act as positive substrates for an axonal route into the spinal cord 30,31 . In this study and in others it is apparent that OECs can coordinate the entry of small-caliber axons at the dorsolateral fasciculus 32,33 . We did not investigate the precise molecular signals or guidance cues which enabled axons to cross into the dorsal horn and terminate in deep Rexed laminae or as far as the ascending dorsal horns 33 .
Since we saw a number of BDA-positive axons within the cuneate fasciculus, we counted axons in this region to compare axon loss in controls with that of the OEC transplanted group. Unexpectedly, axon counts were found to be very similar in these groups following injury, meaning that the presence of OECs did not significantly alter the rate or extent of Wallerian degeneration 34 . Since we assumed that small-caliber axons could be more susceptible to degeneration and given that regenerated axons often have a small diameter, we were surprised that there was no obvious difference in counts between small and large axons. It is possible that this counting method was not sensitive enough to capture the relatively few axons which would be required to reinstate transmission to the cuneate nucleus as seen in our previous electrophysiological study 16 . This quantification method also does not capture the extent of axon loss or any possible neuroprotective effects of OEC transplants in the crucial early couple of weeks following injury. We have previously performed some preliminary studies using MRI scans of ex vivo rat spinal cord tissue. Using this technique it is possible to estimate axonal loss between two treatment groups. Furthermore, we could monitor the loss of axons over time with in vivo studies on spinal cord injury with and without OEC transplants. We may use this method to qualify the axons in future study. From our previous work we know that functional forepaw recovery can occur when as little as 1% of corticospinal tract axons are present 35 . In the current study this would represent only 15–20 small axon bundles, which is most likely too small a number for the quantification method we have used.
In the present study, we used an anatomical approach to investigate how transplanted OECs repair the delicate interface between the PNS and CNS compartments. The results convince us that the reconstituted transitional zone with OEC transplant acts as a bridge to allow sensory axon regeneration into the spinal cord. For the first time, we have found that many migrating OECs extensively ensheath the DRG cells and their axons.
Footnotes
Ethical Approval
Animals were used with the approval of the University College London (UCL) Institute of Neurology.
Statement of Human and Animal Rights
Animals were used according to the United Kingdom (UK) Home Office regulations for the care and use of laboratory animals, the UK Animals (Scientific Procedures) Act 1986.
Statement of Informed Consent
Statement of Informed Consent is not applicable for this article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by funds from the UK Stem Cell Foundation and the Nicholls Spinal Injury Foundation.