
Abstract
As a putative marker of cerebral small vessel disease, cerebral microbleeds (CMBs) have been associated with vascular cognitive impairment. Both iron accumulation and amyloid protein precursor (APP) dysregulation are recognized as pathological hallmarks underlying the progression of CMBs, but their cross-talk is not yet understood. In this study, we found a profound increase of amyloid formation with increasing FeCl3 treatment, and a distinct change in APP metabolism and expression of iron homeostasis proteins (ferritin, Fpn1, iron regulatory protein) was observed at the 300 uM concentration of FeCl3. Further results revealed that extracellular iron accumulation might potentially induce binding of APP to BACE1 for amyloid formation and decrease the capability of APP/Fpn1 in mediating iron export. Our findings in this study, reflecting a probable relationship between iron dyshomeostasis and amyloid pathology, may help shed light on the underlying pathogenesis of CMBs in vascular cognitive impairment.
Introduction
Cerebral microbleeds (CMBs), regarded as a putative marker of cerebral small vessel disease (cSVD), have been associated with cognitive decline in patients with stroke and vascular cognitive impairment. Histologically, CMBs may not only represent the extracellular accumulation of hemosiderin from leakage through cerebral small vessels and pathological iron deposition following hemorrhage and ferritin breakdown, but also correlate with cerebral amyloid angiopathy 1 -4 . Furthermore, several observational studies support that CMBs may relate to amyloid deposition in dementia 5 -9 , and the key finding revealed that ferritin, as a major iron storage protein, adversely impacts dementia progression 10 .
Previous evidence indicated a key mechanism of Aβ toxicity underlying dysregulated processing of amyloid protein precursor (APP) via the switch from the non-amyloidogenic to the amyloidogenic pathway. In the amyloidogenic pathway, APP is cleaved by β-secretase (BACE1) and γ-secretase, leading to increased Aβ42 generation 11,12 . In the non-amyloidogenic pathway, the protective upregulation of APP by the iron regulatory protein (IRP) can be linked to the finding that active interaction of APP with surface ferroportin1 (Fpn1) could not only mediate excessive iron efflux but also maintain cognitive function 13 -16 . Since both the extracellular iron dyshomeostasis and amyloid deposition are recognized as pathological hallmarks of CMBs, insight into the relationship between them might not only increase understanding of the underlying pathogenesis of cSVD, but also convey their importance in vascular cognitive impairment.
Therefore, in the present study, we hypothesized APP as a key link between iron dysregulation and pathogenesis of CMBs, and analyzed the effects of extracellular iron treatment on APP metabolism and its association with the proteins of iron homeostasis including ferritin, Fpn1, and IRP. Considering that the APP/Fpn1 complex mediates iron export at the plasma membrane where APP combines with BACE1, we further investigated changes in the interaction of both complexes in response to iron treatments. Rather than storing a large content of iron, previous findings supported that microglia can rapidly de novo synthesize APP as an acute phase protein in response brain injury 17,18 ; therefore, we used microglia as a model cell line, not neurons. Our findings support a potential relation of extracellular iron dyshomeostasis with APP metabolism, and reflect the underlying pathogenesis of CMBs in the process of vascular cognitive dementia.
Materials and Methods
Cell
BV-2 cells (a microglia cell line) were obtained from the Peking Union Medical College (Beijing, China). Cells were maintained in Dulbecco’s Modified Eagle medium (DMEM; Gibco, Carlsbad, CA, USA) containing 10% fetal bovine serum (FBS; Gibco), 100 U/ml penicillin, and 100 μg/ml streptomycin in a 5% CO2 incubator at 37°C.
Iron Treatment
Cells were cultured for 48 h with variable concentrations of FeCl3 (Sangon Biotech, China) from 25 μM to 400 μM prior to Western blot analysis. Iron was kept soluble using L-ascorbic acid in Tris-HCl (pH = 7.5).
Western Blot
Cells were washed and lysed. Protein concentrations were determined using BCA protein assay kit (Bio-Rad, Hercules, CA, USA). Aliquots of cell lysates containing ∼20 mg of protein were subjected to 10% SDS-PAGE and subsequently blotted onto a PVDF membrane (Bio-Rad). After blocking in PBST with 5% bovine serum albumin for 1 h at room temperature, membranes were incubated with primary antibodies (anti-ferritin: 1:1000; anti-amyloid precursor protein: 1:20000; anti-ferroportin1: 1:200) at 4°C overnight. β-actin served as an internal reference, and the relative expression of target proteins was calculated. The protein bands were scanned into a computer with an Odyssey scanner (Li-COR, Biosciences, Lincoln, NE, USA).
Quantitative Real-Time PCR
Total RNA extraction and cDNA synthesis were performed using TRIZOL reagent (Invitrogen, Grand Island, NY, USA) and reverse transcription kit (TaKaRa, Tokyo, Japan) according to the manufacturers’ instructions. The specific primers used for PCR are: APP forward, 5’-TTCTACACCAGGAGCGGAT-3’, APP reverse, 5’-GGAACTTGTCGATGCCACAGG-3’; ferritin forward, 5’-GCCCCTGAATCTGGCATGG-3’, ferritin reverse, 5’-ATGCACTGCCTCAGTGACC-3’. Quantitative real-time PCR was performed using SYBR FAST qPCR Master Mix (KAPA) with appropriate TaqMan primers in ABI PRISM 7900HT Sequence Detection System (Kapa Biosystems, Wilmington, MA, USA). All mRNA expressions were normalized to that of β-actin as described previously 19 , and the detection was performed in triplicate.
ELISA
Microglia were seeded onto 96-well plates. Cells were treated with different concentrations of FeCl3 for another 48 h. Cell supernatants were collected, and the Aβ42 levels were determined by using an Aβ1-42 ELISA Kit (Roche Diagnostic GmbH, Mannheim, Germany).
siRNA Transfection
Cells were transfected with non-targeting siRNA (negative control) or special siRNA targeting rat APP (5’-CUCAACAUGCACAUGAAUGdTdT-3’) designed by Genepharma using Lipofectamine™2000 reagent (Invitrogen) according to the manufacturer’s instructions. The medium was refreshed 6 h later, and cells were harvested 48 h later.
Co-Immunoprecipitation (CoIP)
Cells were lysed in immunoprecipitation buffer (protease inhibitor and phosphatase inhibitor mixture). Equal amounts of protein (800 μg) from each sample were incubated with protein A/G agarose working solution for 10 min at 4°C under constant shaking. Then, 3 μg of the specified antibody (APP: 1:30; ferroportin1: 1:50; BACE1: 1:50) was added and incubated at 4°C overnight under constant shaking. After washing, the immune complexes in the supernatant were detected by Western blotting.
Statistical Analysis
Data are presented as mean ± SEM. Statistical analyses were completed using SPSS (SPSS Statistics v20; IBM, Armonk, NY, USA). All differences between the means were assessed by two-way analysis of variance (ANOVA), after which Bonferroni post hoc tests were conducted. P-values less than 0.05 were considered as statistically significant.
Results
To ascertain whether iron treatment resulted in Aβ42 accumulation, we first evaluated the levels of Aβ42 by ELISA in microglia treated with increasing doses of FeCl3. Considering that brain iron is present at a concentration of approximately 1 mM surrounding amyloid plaques 11 , we controlled the iron concentration under this level. After 48 h treatments, we did not find cell viability CCK8 assay at 300 μM concentration of FeCl3 reduced significantly as compared with control (Sup. 1). Next significantly increased levels of Aβ42 (p < 0.01) were found at the concentration of 300 μM (Fig. 1). We further characterized whether the increase in extracellular iron content was related to changes in the levels of iron transporters. Microglia treated with increasing FeCl3 concentrations for 48 h showed reduced levels of APP proteins and elevated ferritin protein levels below 300 μM treatment. When FeCl3 concentrations reached 300 μM, levels of APP protein in microglia significantly increased, while ferritin production was decreased (Fig. 2).

Effects of extracellular iron treatments on the changes in Aβ42 formation in microglia. Microglia was treated with increasing dose of FeCl3 for 48 h after which the levels of Aβ42 were analyzed by ELISA. A significant increase in the level of Aβ42 (p < 0.01) is observed at 300 μM FeCl3 compared with that at 200 μM. Error bars represent mean ± SEM (n = 3). *p < 0.05 and **p < 0.01 as compared with control; #p < 0.05 as compared with 200 μM FeCl3 treatment group.

Effects of extracellular iron treatments on the levels of APP and ferritin proteins in microglia. Microglia was treated with increasing dose of FeCl3 for 48 h after which we evaluated the levels of APP and ferritin proteins by Western blot. Upper panel (a) shows representative blots and lower panels represent the relative quantification of band density in APP (b) and ferritin (c), respectively. Values represent mean ± SEM (n = 4). *p < 0.05 and **p < 0.01 as compared with control; # p < 0.05 and ## p < 0.01 as compared with 200 μM FeCl3 treatment group.
To determine if changes in microglial protein levels were consistent with changes in mRNA levels, qPCR was conducted after treatment with iron for 48 h using specific primers for APP and ferritin (Fig. 3). Results showed that changes in APP mRNA and ferritin mRNA were similar to the changes in their protein levels. Microglia treated with iron concentrations below 300 μM presented decrease in APP mRNA content but increase in ferritin mRNA levels. APP mRNA increased significantly in microglia while decreased ferritin mRNA was observed following 300 μM FeCl3 treatment.
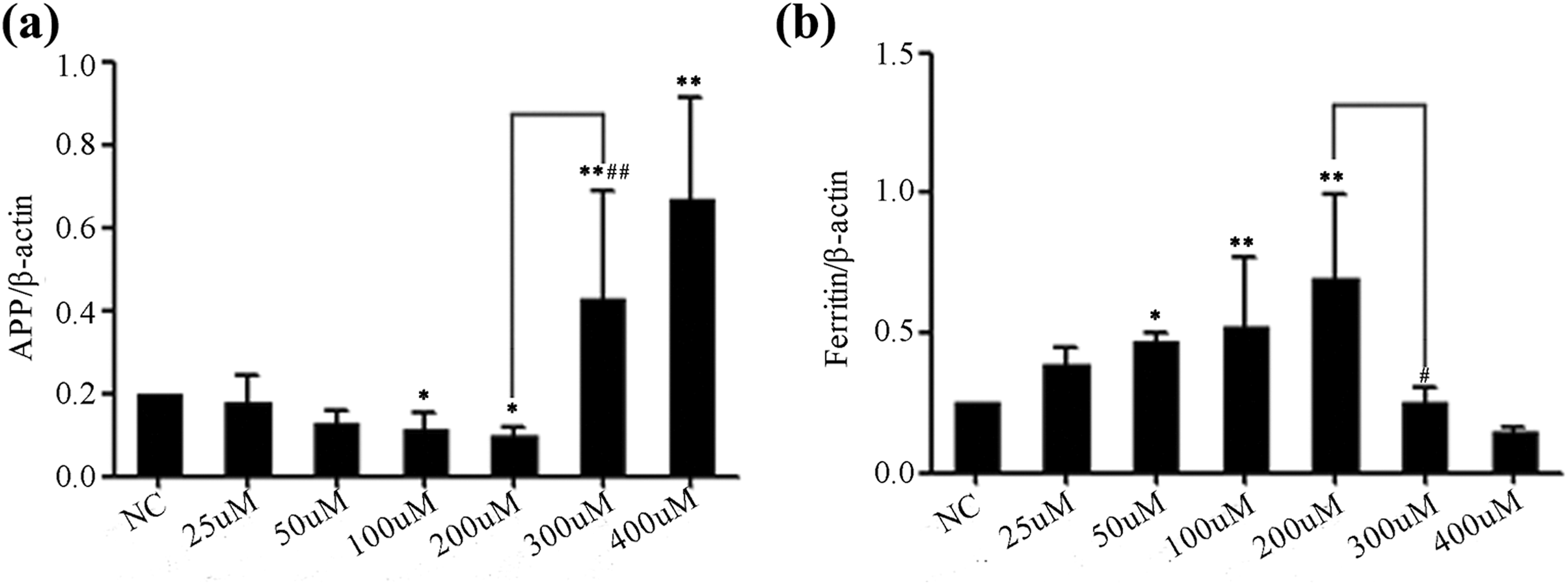
Effects of extracellular iron treatments on the changes in the mRNA levels of APP and ferritin in microglia. Microglia were treated with increasing doses of FeCl3 for 48 h. Relative mRNA expression levels of APP (a) and ferritin (b) were analyzed by RT-PCR. Values represent mean ± SEM (n = 4). *p < 0.05 and **p < 0.01 as compared with control; # p < 0.05 and ## p < 0.01 as compared with 200 μM FeCl3 treatment group.
Considering that APP is suggested to be associated with amyloidogenesis and iron dyshomeostasis, it is of interest to determine the putative changes in iron metabolism proteins such as ferritin, IRP, and Fpn1 in the absence of APP. To this end, we detected an increase in APP and ferritin proteins by 300 μM iron treatment, and found decreased levels of IRP and Fpn1 proteins compared with control groups. In the absence of APP mediated by siRNA, iron treatment also induced a significant decrease in IRP and Fpn1 proteins and elevated APP proteins, whereas ferritin levels remained unchanged (Fig. 4).
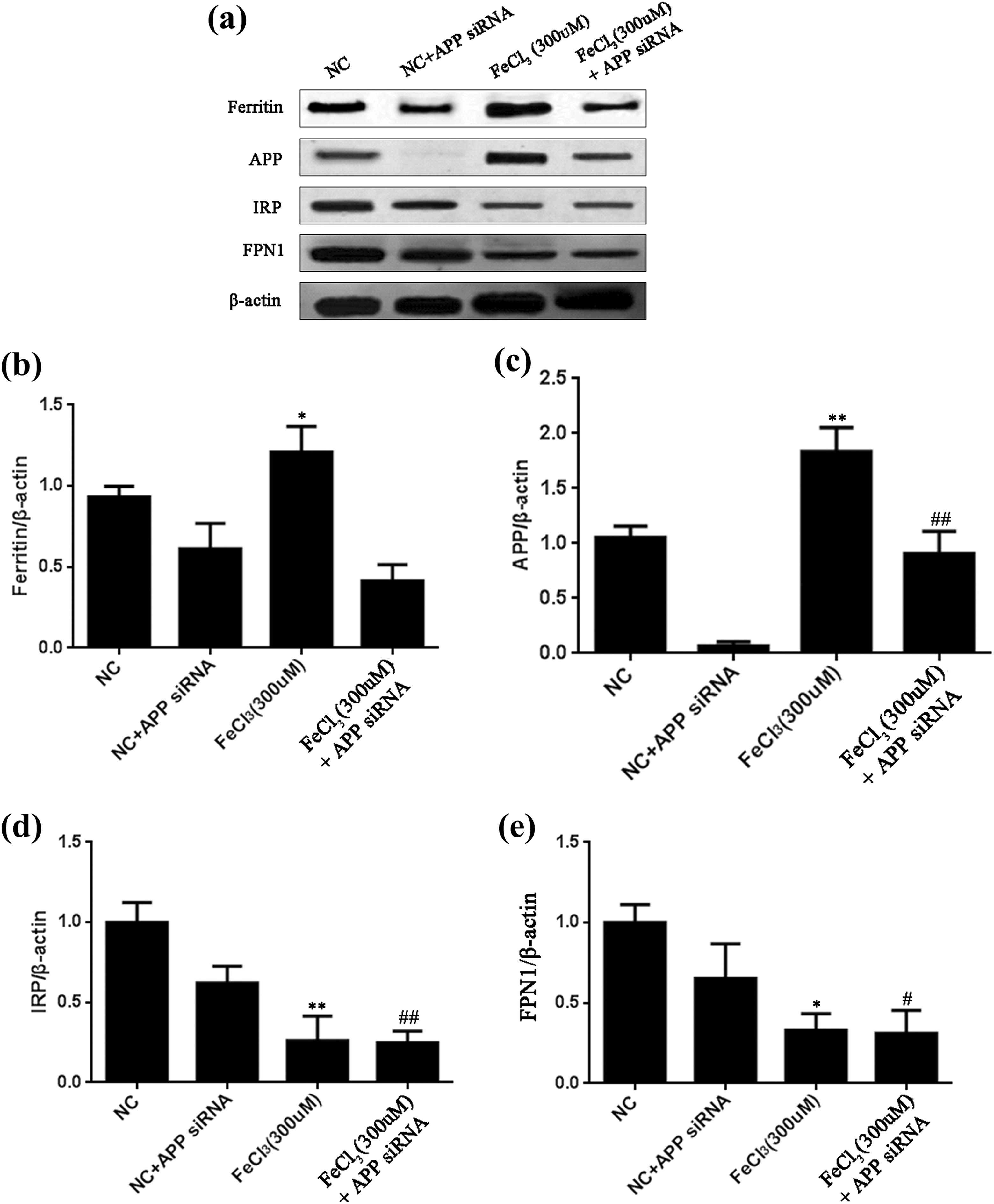
Extracellular iron treatments of microglia induce changes in the expression levels of iron metabolism proteins in the presence and absence of APP mediated by siRNA. Microglia was treated with 300 μM iron treatment for 48 h. Changes in iron metabolism proteins were determined by Western blot. Panel (a) shows representative blots and the panels below show the quantification of band density in ferritin (b), APP (c), IRP (d), and Fpn1 (e) in the presence and absence of APP, respectively. Values represent mean ± SEM (n = 4). *p < 0.05 and **p < 0.01 as compared with control groups in the presence of APP; # p < 0.05 and ## p < 0.01 as compared with control groups in the absence of APP.
According to the above changes, we next evaluated if microglia treated with iron could affect the interaction between APP and Fpn1 using a CoIP technique. Although CoIP results do not necessarily mean the interaction is direct, it suggests a probable binding capability of a complex. There was a reduced ratio of Blot: Fpn1/Input: Fpn1, likely due to a decreased affinity of APP/Fpn1, observed following iron treatment in treated groups compared with controls (Fig. 5). We also evaluated the change in binding of APP and BACE1 during the interaction APP/Fpn1, and found that iron treatment of microglia showed an increased interaction between APP/BACE1, without significant changes in the level of BACE1 (Fig. 6).

Extracellular iron treatments induce changes in the interaction of APP and FPN1 proteins in microglia. Microglia was treated with 300 μM iron. Changes in the interaction of APP and FPN1 were analyzed by coimmunoprecipitation (CoIP). Panel (a) shows representative blots and lower panels show the ratio of band density in Blot:FPN1/Input:FPN1(b), and quantification of band density in Input:FPN1 (c), respectively. Values represent mean ± SEM (n = 3). *p < 0.05 as compared with control.

Extracellular iron treatments induce changes in the interaction of APP and BACE1 proteins in microglia. Microglia was treated with 300 μM iron. Changes in the interaction of APP and BACE1 were analyzed by CoIP. Panel (a) shows representative blots and lower panels show the ratio of band density in Blot:BACE1/Input:BACE1 (b), and quantification of band density in Input:BACE1 (c), respectively. Values represent mean ± SEM (n = 3). **p < 0.01 as compared with control.
Discussion
The pathological hallmarks of CMBs in the brain are not only increased extracellular iron deposition, but also excessive amyloid plaques 5 . However, the molecular mechanisms of how amyloidosis and extracellular iron accumulation cross-talk remain unknown. Our results showed that extracellular iron treatment exercised profound effects on APP metabolism, and induced a significant increase of amyloid protein levels at the FeCl3 concentration of 300 μM. Furthermore, a distinct change in APP metabolism and expression of iron homeostasis proteins (ferritin, Fpn1, IRP) was observed at the 300 uM concentration of FeCl3. We supposed that this concentration was likely to reflect the microglia status switching from the homeostasis to dysregulation in the present experimental condition. Finally, the increase in amyloid formation might be partially explained if extracellular iron accumulation was related to the reduced iron export by the complex of APP/Fpn1 and higher affinity of APP/BACE1. Our data provide a potential mechanism for the previous findings that CMBs are associated with amyloid pathology in the brain, the latter being related to the ferritin in CSF 5,8,10 .
In the central nervous system, the largest iron stores are found within microglia and modulated by ferritin, which plays the crucial role of controlling iron storage and homeostasis. Thus, microglia are supposed to be naturally responsible for the maintenance of the extracellular iron status among other brain cell types 20 . Moreover, rather than clearance of amyloid plaques, microglia can induce APP production as an acute phase protein in response to brain attacks 17,18 . For these reasons, we used microglia as a model cell line in this study rather than neurons, which appeared to be commonly used in other experiments. In microglia, extracellular iron treatment at the concentration of 300 μM dramatically increased the levels of Aβ42. The rise in Aβ42 by accumulated iron may be a consequence of the activation of oxidative stress (OS) signaling pathways resulting in excess amyloid production 21 -24 . The cause for the slight increase in amyloid formation in response to iron treatment below 300 μM is unknown, but could be because of a compensatory response to reduce OS 25,26 .
Previous studies suggested that APP could serve as a cell-surface pro-inflammatory receptor on microglia, and that it regulates microglial activation 27 . To this end we assumed the transient decrease in APP could be a result of the neuroprotective response below iron concentrations of 300 μM. Likewise, the increase in ferritin could be the natural function of excess iron for storage in order to protect against the OS response by iron regulatory protein/iron responsive element (IRP/IRE) regulatory system 28,29 . However, microglia presented an inverse pattern, including significant increase in APP and reduction of ferritin, at iron concentrations of 300 μM and above. The levels of ferritin were reduced in the presence of iron treatment higher than 300 μM, probably as a consequence of ferritin degradation due to iron-dependent oxidation and reduced ferritin synthesis 29 . Increased APP in response to 300 μM iron treatment is consistent with the previous observation of IRP/APP IRE-driven abundant iron efflux, and may be a response to severe oxidative challenges 26 .
To test the effects of APP on major iron metabolism proteins such as ferritin, IRP, and Fpn1 in microglia, we assessed the changes of these proteins in the absence of APP mediated by siRNA. Consistent with previous studies, downregulated IRP levels following iron treatment could be a consequence of regulation of IRP/IRE iron homeostasis system 11,15 . Unexpectedly, it resulted in a significant reduction in Fpn1 protein, probably independent of the existence of APP. Although iron accumulation was thought to be associated with elevated Fpn1 levels, previous research showed that inflammatory stimuli caused by iron could decrease Fpn1 expression and trigger a positive feedback loop of elevated iron deposition in microglia 30 -32 . It can be inferred that high iron concentrations may alter the expression of Fpn1 as we observed in this study.
Following the changes in the above proteins, such as the reduced level of Fpn1, it was of interest to evaluate if the affinity of interaction between increased APP and decreased Fpn1 could be affected. We used CoIP to show that there was a decreased level of Fpn1 immunoprecipitated by APP antibody, partially suggesting reduced affinity of the APP/Fpn1 complex. However, rather than the potential decreased ability of interaction, this finding might be also attributed to the possibility of decreased input of Fpn1 protein amount itself. In addition, we also investigated whether the “relative excess” APP could turn to interact with BACE1, another membrane protein in microglia, during the binding of the APP/Fpn1complex. The cause for the observed increase in the interaction of APP/BACE1 complex without significant changes in the level of BACE1 is unknown, but the process of interaction is a key step in the amyloid pathway. Therefore, it could partially explain why there is a significant increase in Aβ42 levels at iron concentrations of 300 μM. Our findings might suggest a potential relationship for both complexes of APP/Fpn1 and APP/BACE1, providing new insights for further investigation; however, it should be carefully considered that CoIP does not mean the interaction is direct, since larger complexes may be involved.
In summary, extracellular iron accumulation induced a significant increase in amyloid formation, and was likely due to both the APP dysregulation and iron perturbation. Changes in extracellular iron accumulation might potentially induce the interaction between APP and BACE1 which is necessary for amyloid production, but also reduce the capability of APP/Fpn1 mediating iron homeostasis (Fig. 7). The data presented here establish a potential key link of extracellular iron dyshomeostasis and APP metabolism, supporting the role of CMBs in the pathogenesis of vascular cognitive dementia.

Schematic representation of changes in extracellular iron accumulation and amyloid formation in microglia. 300 μM iron treatments of microglia elevated the APP protein level but reduced the levels of Fpn1 and IRP. Furthermore, the iron treatment might reduce the capability of APP/Fpn1 complex for iron efflux and induce binding of APP to BACE1 for amyloid protein production.
Supplemental Material
Supplemental Material, sup_1 - Iron Dyshomeostasis Induces Binding of APP to BACE1 for Amyloid Pathology, and Impairs APP/Fpn1 Complex in Microglia: Implication in Pathogenesis of Cerebral Microbleeds
Supplemental Material, sup_1 for Iron Dyshomeostasis Induces Binding of APP to BACE1 for Amyloid Pathology, and Impairs APP/Fpn1 Complex in Microglia: Implication in Pathogenesis of Cerebral Microbleeds by Li Gong, Xiangzhu Tian, Jing Zhou, Qiong Dong, Yan Tan, You Lu, Jiayan Wu, Yanxin Zhao and Xueyuan Liu in Cell Transplantation
Footnotes
Ethical Approval
This study was approved by the Ethics Committee of Shanghai Tenth People’s Hospital (Shanghai, China).
Statement of Human and Animal Rights
This article does not contain any studies with human or animal subjects.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grant from National Natural Science Foundation of China (No.81771131, No.81571033), and by Science and Technology Commission of Shanghai Municipality (No.17411950100, No.17411967500).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.