
Abstract
Streptozotocin (STZ) is a cytotoxic glucose analogue that causes beta cell death and is widely used to induce experimental diabetes in rodents. The sensitivity of beta cells to STZ is species-specific and human beta cells are resistant to STZ. In experimental islet transplantation to rodents, STZ-diabetes must be induced before transplantation to avoid destruction of grafted islets by STZ. In human islet transplantation, injection of STZ before transplantation is inconvenient and costly, since human islet availability depends on organ donation and frail STZ-diabetic mice must be kept for unpredictable lapses of time until a human islet preparation is available. Based on the high resistance of human beta cells to STZ, we have tested a new model for STZ-diabetes induction in which STZ is injected after human islet transplantation. Human and mouse islets were transplanted under the kidney capsule of athymic nude mice, and 10–14 days after transplantation mice were intraperitoneally injected with five consecutive daily doses of STZ or vehicle. Beta-cell death increased and beta-cell mass was reduced in mouse islet grafts after STZ injection. In contrast, in human islet grafts beta cell death and mass did not change after STZ injection. Mice transplanted with rodent islets developed hyperglycemia after STZ-injection. Mice transplanted with human islets remained normoglycemic and developed hyperglycemia when the graft was harvested. STZ had no detectable toxic effects on beta cell death, mass and function of human transplanted islets. We provide a new, more convenient and cost-saving model for human islet transplantation to STZ-diabetic recipients in which STZ is injected after islet transplantation.
Introduction
Streptozotocin (STZ, 2-deoxy-2-(3-(methyl-3-nitrosoureido)-D-glucopyranose) is a glucosamine-nitrosurea compound that selectively destroys pancreatic beta cells. The beta cell selectivity is conferred by the glucose moiety that allows the intracellular uptake in cells expressing the glucose transporter GLUT-2 1 –5 , and the toxicity is dependent upon the DNA-alkylating activity of its methylnitrosourea moiety 6 –8 . Thus, cells that do not express GLUT-2 are resistant to STZ 5,9 . Rodent pancreatic beta cells have GLUT-2 as the main glucose transporter and are highly sensitive to STZ damage. However, GLUT-2 expression is very low in human beta cells, and GLUT-1, that has very low or no affinity for STZ, is the predominant glucose transporter 5,10,11 . Thus, human beta cells are highly resistant to the toxic effects of STZ 12 –14 . The resistance, however, is not complete and STZ is used in oncology as a chemotherapeutic agent to treat metastatic insulinomas 15 –18 .
Reversal of diabetes after human islet transplantation to STZ-diabetic mice is considered the gold-standard method to assess the quality of human islet preparations, as it has shown the best correlation with graft function after islet transplantation to diabetic patients 19,20 . The model has been also used to evaluate the degree of maturation and function of insulin-producing cells differentiated from human stem cells 21 –23 . In experimental islet transplantation to rodents, STZ-diabetes must be induced before islet transplantation to avoid the concomitant destruction of grafted islets. Thus, STZ is first injected and once diabetes is confirmed several days later, islet transplantation is performed 24 –26 . The availability of rodent islet donors is at the discretion of the investigator, and islet isolation and transplantation can be planned well in advance and conveniently performed once diabetes has been confirmed in the recipient. In contrast, human islet isolation and transplantation can’t be planned ahead of time since pancreas availability depends on organ donation. Thus, prospective STZ-diabetic recipients of human islet transplantation, usually frail immunodeficient mice requiring more intensive care, including daily treatment with insulin, must be kept in the animal facility for unpredictable lapses of time, until a human islet preparation is available. Based on the well-known resistance of human beta cells to STZ toxicity, we have investigated a new model for experimental human islet transplantation in which islets are transplanted to healthy recipients, and STZ is injected several days later, after the islets’ engraftment. The model reduces the burden and cost of the procedure and increases the animal welfare.
Materials and Methods
Animals
Male athymic Nude-Foxn1nu mice (Harlan Laboratories, Horst, The Netherlands) housed under specific pathogen-free conditions were used as recipients of human and mouse islet transplantation. Male inbred C57Bl/6 mice (Harlan France SARL, Gannat, France) were used as donors of mouse islets for transplantation. Mice were maintained in the animal facility of Institut d’Investigació Biomèdica de Bellvitge (IDIBELL) (Association for Assessment and Accreditation of Laboratory Animal Care International accreditation number 1155). Experimental procedures were approved by the Ethical Committee for Animal Experimentation of IDIBELL.
Experimental Design
Athymic Nude-Foxn1nu mice were transplanted with 2000 (n = 14) or 800 (n = 9) human islets, or with 100–200 C57Bl/6 mouse islets (n = 15) and treated with STZ or vehicle 10–14 days later (Figure 1). Two thousand human islets are required to restore normoglycemia when transplanted to already diabetic, STZ-injected, mice 27 , and 800 islets are a clearly insufficient beta cell mass in this model 28,29 . The 2000 human islet and the mouse islet grafts were harvested 6 h after the last injection of STZ or vehicle. The 800 human islet grafts were not harvested after STZ injection, and blood glucose was monitored for an additional week to determine whether they could maintain normoglycemia. A group of non-transplanted mice treated with STZ (n = 9) was included in the study as controls for blood glucose.

Experimental design. Human and mouse islets were transplanted under the kidney capsule of athymic nude mice (Tx). Ten to 14 days after transplantation, mice were intraperitoneally injected with five consecutive daily doses of either STZ (70 mg/kg body weight) or vehicle (citrate). Grafts were harvested on the last day of STZ injection to determine beta cell death, mass and vascular density or followed for seven additional days to assess the metabolic evolution.
Streptozotocin (Sigma Immunochemicals, St Louis, MO, USA), freshly dissolved in citrate buffer (pH = 4.5), was injected intraperitoneally in five consecutive daily doses (70 mg/kg body weight, total dose 350 mg/kg). Blood was obtained between 9:00 and 11:00 a.m., in non-fasting conditions from the snipped tail and glucose was measured with a portable meter (Glucocard Memory, A. Menarini Diagnostics, Barcelona, Spain). Body weight was simultaneously determined. Animals were considered hyperglycemic when blood glucose values were >11.1 mmol/l on two consecutive measurements.
Human and Mouse Islet Isolation and Transplantation
Human pancreatic islets were isolated from six adult cadaveric organ donors, age 58 ± 6.6 years, by collagenase digestion (Collagenase NB1 Premium Grade with Neutral Protease NB, Serva Electrophoresis GmbH, Heidelberg, Germany) using the Ricordi method, purified on a refrigerated COBE 2991 cell processor (COBE BCT, Laekwood, Colorado, USA), and transplanted, as previously described 30 . Use of human islets was approved by the Ethics Committee of Hospital of Bellvitge, and written, signed consent was obtained from donor relatives. Mouse islets were isolated by collagenase (Collagenase P; Roche diagnostics, Mannheim, Germany) digestion of the pancreas 26 .
Islets were transplanted under the kidney capsule. To harvest the graft, the kidney capsule surrounding the graft was incised and removed with the graft. Grafts were immediately immersed in 4% paraformaldehyde-PBS, and after the removal of any excess paraformaldehyde by capillary action, weighed and processed for paraffin embedding 26 .
Immunohistochemical Studies
Beta cell death
Graft sections were double-stained for apoptotic nuclei using the TUNEL technique (In Situ Cell Death Detection Kit, ApopTag; Millipore, Temecula, CA, USA), and for insulin with a rabbit polyclonal anti-insulin antibody (Santa Cruz Biotechnology Inc., Dallas, TX, USA) (final dilution 1:100). For insulin labeling, the secondary antibody was a donkey anti-rabbit IgG conjugated with Alexa Fluor 555 (Life Technologies, Eugene, OR, USA; final dilution 1:400). Nuclei were stained with 300 nM DAPI (Life Technologies). TUNEL-positive beta cells were assessed using a Leica DM4000B microscope connected to a digital camera (Leica DFC310 FX) (Wetzlar, Germany) with a color monitor. Necrotic areas were excluded. Beta cell death was expressed as percentage of TUNEL-positive beta cells. A minimum of 1200 cells per sample were counted.
Beta cell mass and endocrine non-beta cell mass
Beta cell and endocrine non-beta cell mass was measured by point-counting morphometry on sections double-stained for insulin with a chicken anti-human antibody (Abcam, Cambridge, UK) (final dilution 1:1000) and for the endocrine non-beta cell mass with a cocktail of antibodies including rabbit anti-human glucagon (Cell Signaling Technology, Inc, Beverly, MA, USA) (final dilution 1:100), rabbit anti-human somatostatin (Dako, Carpinteria, CA, USA) (final dilution 1:500) and rabbit anti-human pancreatic polypeptide (ChemiconInternational, Inc., Temecula, CA, USA) (final dilution 1:2000) 31 . Insulin and endocrine non-beta cells were visualized with Alexa Fluor 488 and 555 (final dilution 1:400) under a fluorescent microscope. Nuclei were stained with 300 nM DAPI (Life Technologies). Beta cell mass and endocrine non-beta cell mass was obtained by multiplying the weight of the graft by the relative beta or endocrine non-beta cell volume respectively 31 .
Beta cell vascular density
Graft sections were double-stained for endothelial cells with a CD31 antibody and for beta cells with an insulin antibody. CD31 staining was performed using a mouse monoclonal anti-human antibody (DAKO, Carpinteria, CA, USA) for human grafts, and a rabbit polyclonal anti-mouse antibody (Abcam, Cambridge, UK) for mouse grafts, and visualized with the EnVision+ System-HRP (DAKO, Carpinteria, CA, USA). For insulin double-staining, sections stained for CD31 were incubated with a rabbit polyclonal anti-insulin antibody (Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and visualized with the EnVision+ System-HRP. Vascular density was determined by point counting morphometry, dividing the number of intercepts over CD31-positive endothelial cells located in the insulin-positive beta-cell area by the number of intercepts over the whole beta-cell area 32 .
Statistical Analysis
Results were expressed as means ± SEM. Statistics were performed using SPSS 14.0 (Chicago, IL, USA) for Windows or GraphPad Prism 4 software (La Jolla, CA, USA), and differences between means were evaluated using the Student’s t-test or the one-way analysis of variance (ANOVA) combined with Tukey’s test for post-hoc analysis, as appropriate. A p value of less than 0.05 was considered significant.
Results
Metabolic Outcome
Human islets induced a mild reduction in blood glucose after transplantation into normoglycemic mice, in agreement with the different range of normal glucose values in humans and rodents 33 . All animals transplanted with human islets remained normoglycemic after STZ injection, even when they were transplanted with only 800 human islets, and maintained blood glucose values similar to those of the vehicle-treated group (Figure 2(a)). When human islet grafts were harvested after last STZ injection, all animals developed acute and severe hyperglycemia, confirming that transplanted human islets were responsible for the maintenance of normoglycemia after STZ injection. Accordingly, the pancreases of transplanted and STZ-treated groups showed only a few insulin-positive beta cells at the end of follow up, consistent with STZ-induced death of most beta cells (supplementary Figure 1). All non-transplanted animals, and 87% of recipients transplanted with mouse islets, developed diabetes before graft removal (Figure 2(b)).

Metabolic evolution in mice transplanted with human and mouse islets, and treated with STZ. (a) Human islet transplantation. Green: Transplanted with 2000 islets, STZ treated (n = 8). Blue: Transplanted with 800 islets, STZ treated (n = 9). Red: Non-transplanted, STZ treated (n = 9). Orange: Transplanted with islets, vehicle treated (n = 6). Values are means ± SEM. ANOVA, p < 0.0001, with post-hoc Tukey’s test for multiple comparisons, *p < 0.05 v. STZ-treated and transplanted, and vehicle-treated and transplanted groups. (b) Mouse islet transplantation. Green: Transplanted, STZ treated (n = 8). Red: Non-transplanted, STZ treated (n = 9). Orange: Transplanted, vehicle treated (n = 7). Values are means ± SEM. ANOVA, p < 0.0001, with post-hoc Tukey’s test for multiple comparisons, *p < 0.05 v. STZ-treated and transplanted, and vehicle-treated and transplanted groups; †p < 0.001 v. vehicle-treated and transplanted group. Tx: transplantation day; STZ: streptozotocin; Arrows: STZ injection. Bold arrow: removal of the graft in transplanted (2000 islets) and treated with either STZ (green) or vehicle (orange) groups.
Beta Cell Death and Mass in Human and Mouse Islet Grafts
In transplanted mouse islets, beta cell death was increased (Figure 3) and beta cell mass was reduced (Figure 4(a)) after STZ injection. The 40% reduction in beta cell mass in mouse islet grafts after the fifth STZ injection is consistent with the previously reported islet loss in the endogenous pancreas in the multiple low-dose STZ model 34 . In contrast, beta cell death and mass in transplanted human islets was similar after STZ or vehicle injection. Similar to differences in initially transplanted islet mass in human and mouse grafts, beta cell mass was 10 times higher in human grafts harvested from both STZ- and vehicle-injected mice (0.91±0.13 mg and 0.73±0.081 mg respectively) than in mouse grafts transplanted to vehicle-injected recipients (0.080±0.017 mg), suggesting a comparable survival of human and mouse beta cells after transplantation. In mouse and human islet grafts, the endocrine non-beta cell mass was similar in STZ- and vehicle-injected mice (mouse islet graft: 0.017±0.0033 mg and 0.018±0.0043 mg); (human islet graft: 0.43±0.087 mg and 0.44±0.105 mg) (Figure 4(b)), confirming the beta cell selectivity of STZ toxicity.

Beta cell death in human and mouse islet grafts. (a) Representative images of mouse and human islet grafts from vehicle- and STZ-treated groups double stained for insulin (red) and TUNEL (green). (b) Quantification of beta cell death in human and mouse islet grafts after vehicle (n = 6 and 8 respectively, white bars) or STZ (n = 7 in each group, black bars) injection. Values are means ± SEM. Student’s t-test *p < 0.001.
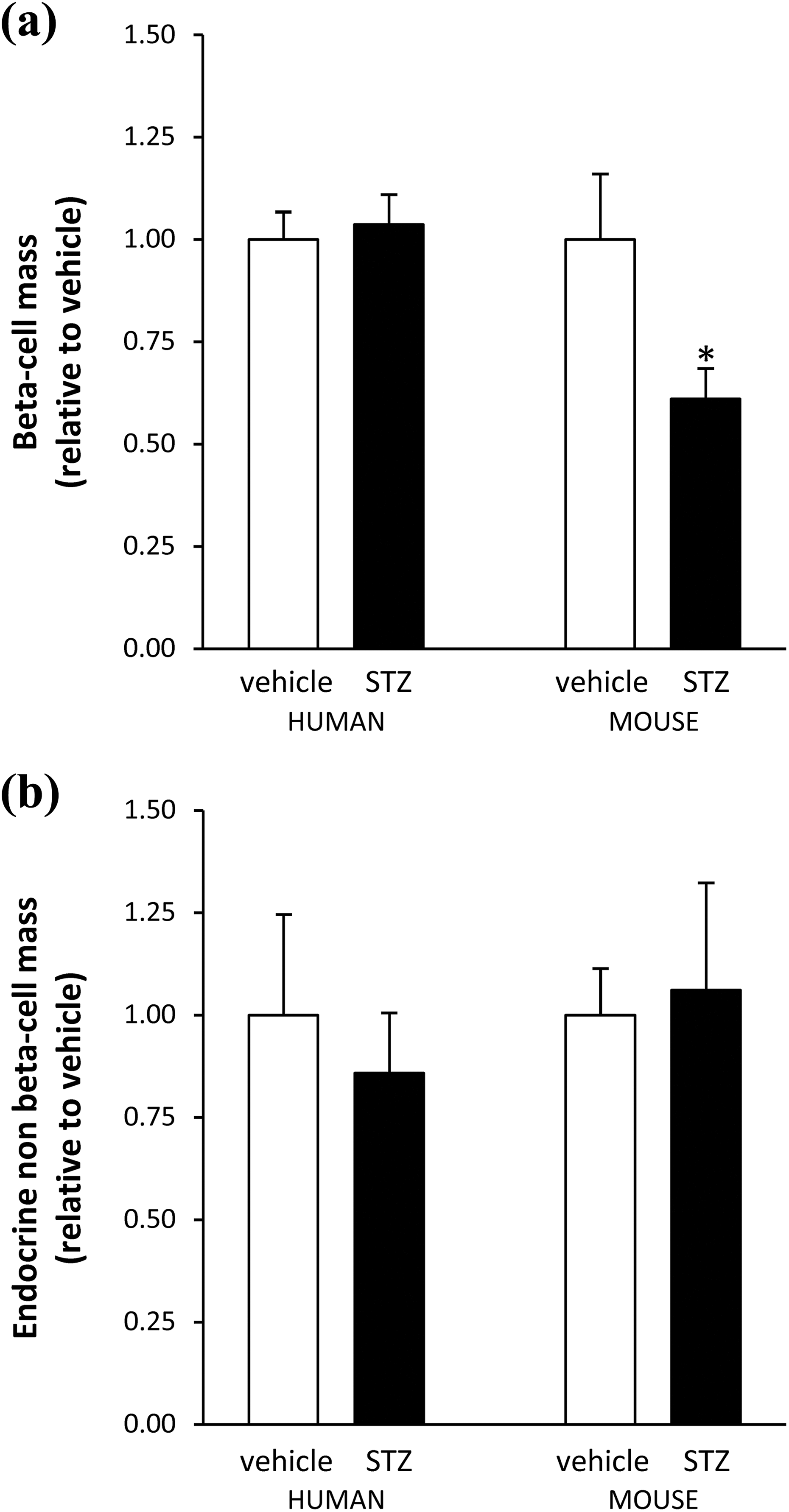
Beta cell mass and endocrine non-beta cell mass in human and mouse islet grafts. (a) Beta cell mass in human and mouse islet grafts after vehicle (white bar) or STZ (black bar) injection. (b) Endocrine non-beta cell mass in human and mouse islet grafts after vehicle (white bar) or STZ (black bar) injection. Values are means ± SEM. Student’s t-test *p < 0.02.
Beta Cell Vascular Density in Islet Grafts
Beta cell vascular density was determined in islet grafts harvested 6 h after the last STZ injection to ensure that they were revascularized when STZ was injected. Vascular density was similar in STZ- and vehicle-injected groups, both in human and mouse islet grafts (Figure 5). Vascular density was lower in human compared to mouse islet grafts.

Beta cell vascular density in human and mouse islet grafts. Quantification of beta cell vascular density in human and mouse islet grafts after vehicle (white bar) or STZ (black bar) injection. Values are means ± SEM.
Discussion
In this study, we describe a model for experimental human islet transplantation into STZ-diabetic mice in which, taking advantage of the well-known resistance of human beta cells to STZ damage 12 –14 , STZ injection was performed after transplantation when islets were already engrafted and vascularized. We found no detectable toxic effects of STZ on transplanted human islets, as indicated by the maintenance of normal fed blood glucose and the absence of changes in beta cell death and mass in islet grafts after STZ injection. The model is more convenient, cost-saving, and increases the welfare of the animals.
Differences in beta cell susceptibility to STZ damage between humans and rodents, as well as among different rodent strains, are well known 13,14,35 . Rodent beta cells are very sensitive to the toxic effects on STZ, while human beta cells are highly resistant to STZ. The resistance of human transplanted islets to high doses of STZ was reported by Yang et al. 14 that showed sustained normoglycemia and maintained beta granulation in islet grafts after STZ injection. However, they did not quantify beta cell death nor mass in the grafts, and recurrence of hyperglycemia after graft harvesting was not confirmed, an essential step to prove that transplanted islets were responsible for the maintenance of normoglycemia after STZ injection. In experimental human islet transplantation this assessment is particularly important because it has been reported that insulin treatment prevents the diabetogenic action of STZ in rodents, probably by a combination of decreased cellular activity of beta cells and reduced expression of GLUT-2 in insulin treated animals 36 . Thus, the insulin secreted by human transplanted islets, which induces a mild reduction in blood glucose, could provide some protection to the endogenous beta cells of recipient mice when STZ is injected. In our model, the development of hyperglycemia in all STZ-injected animals after graft harvesting confirmed that the endogenous pancreatic beta cells have been severely damaged by STZ injection, and that normoglycemia was maintained by the preserved beta cell function of transplanted human islets. Moreover, we show that transplanted beta cell death was not increased and beta cell mass was preserved after STZ injection. As expected, animals transplanted with mouse islets showed increased beta cell death and reduced beta cell mass in the graft, and developed severe hyperglycemia after STZ injection.
Graft vascularization was determined to confirm that injected STZ could reach the islet grafts, and that preservation of transplanted beta cell function and survival was not due to lack of exposure to STZ. We show that islet grafts were already vascularized when STZ was injected, and that vascular density was similar in STZ- and vehicle-treated animals in both mice and human grafts, indicating that STZ did not modify the islet-graft vasculature. Vascular density was lower in human islet grafts, in agreement with known data about vascularization of human and mouse islets in the endogenous pancreas 37 . The results provide additional indirect support to the concept that the absence of deleterious effects of STZ injection in islet grafts was due to the resistance of human beta cells to STZ toxicity.
The model that we describe has several advantages. First, it is not necessary to maintain a pool of chronically hyperglycemic immunodeficient recipients for undetermined periods of time, waiting for a human islet preparation to become available. These animals require additional care, including daily treatment with insulin, and they increase the burden and cost of the experiments. Second, the model reduces the number of human islets that must be transplanted. It is well established that restoration of normoglycemia in STZ-diabetic recipients requires the transplantation of a higher islet number than that needed to maintain normoglycemia 38 –40 . In our model normoglycemia was maintained with the transplantation of just 800 human islets, a clearly insufficient islet mass to restore normoglycemia when transplanted to STZ-diabetic recipients 28,29 . Third, the model can also reduce the number of animals used in the experiments. The recipients of human islet transplantation are usually frail immunodeficient mice. After STZ injection, and despite insulin treatment, their condition often deteriorates, and mortality is increased 41 . Thus, the survival, and also the welfare, of the animals is increased in the current model. Moreover, the number of animals to be injected with STZ is precisely known, since they have already been transplanted, and this may additionally reduce the use of animals. Fourth, the model is closer to the usual practice in clinical islet transplantation, where normoglycemia is maintained before and after the procedure to avoid the deleterious effects of hyperglycemia on beta cells. In brief, the procedure is convenient, reduces the workload of animal caretakers and investigators, reduces the number of islets and of animals required for transplantation, and it increases the comfort of the animals.
In summary, we have shown that the administration of STZ after human islet transplantation does not damage the grafted beta cells and is a more convenient and affordable model for pre-clinical human islet transplantation. The model may also be valuable to assess the function of insulin-producing cells differentiated from human embryonic stem cells or other human cell types.
Supplemental Material
Fig._Supplementary_1 - A model for human islet transplantation to immunodeficient streptozotocin-induced diabetic mice
Fig._Supplementary_1 for A model for human islet transplantation to immunodeficient streptozotocin-induced diabetic mice by Elisabet Estil·les, Noèlia Téllez, Montserrat Nacher, and Eduard Montanya in Cell Transplantation
Footnotes
Acknowledgments
We thank Cristofer Garcia for skillful technical assistance.
Ethical Approval
The isolation and the usage of human islets were approved by the Ethics Committee of Hospital de Bellvitge. All procedures in the animal experiment described in the study were reviewed and approved by the Ethical Committee for Animal Experimentation of the Bellvitge Biomedical Research Institute (IDIBELL).
Statement of Human and Animal Rights
Human Rights: The procedures and the experiments we’ve performed with appropriate consent and ethical approval from Ethics Committee of Hospital de Bellvitge. Animal Rights: The experiments on animals were conducted in accordance with local Ethical Committee laws and regulations as regards care and use of laboratory animals.
Statement of Informed Consent
The human islets were obtained with written informed consent from donors’ relatives.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from Instituto de Salud Carlos III (ISCIII) (P1600462), ACCIÓ (Catalonia Trade & Investment; Generalitat de Catalunya) in the context of ADVANCE.CAT (EM), and the European Community under the ERDF operational program (European Regional Development Fund, FEDER) ‘a way to build Europe’. CIBER of Diabetes and Associated Metabolic Diseases (CIBERDEM) is an initiative from ISCIII.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.