
Abstract
Alzheimer’s Disease (AD) is the leading cause of dementia in the elderly. In healthy individuals, amyloid precursor protein (APP) is cleaved by α-secretase, generating soluble α-amyloid precursor protein (sAPPα), which contributes neuroprotective functions in the neuronal environment. In contrast, in the neurodegenerative environment of AD patients, amyloid-β-peptide (Aβ) of either 40 or 42 residues are generated by increased activity of β- and γ-secretase. These proteins amalgamate in specific regions of the brain, which disrupts neuronal functions and leads to cognitive impairment. Human umbilical cord blood cells (HUCBC) have proven useful as potential immunomodulatory therapies in various models of neurodegenerative diseases, including AD. Our most recent work studied the impact of umbilical cord blood serum (CBS) on modulation of sAPPα production. Heat-sensitive CBS significantly promoted sAPPα production, indicating that heat-sensitive factor(s) play(s) a role in this process. Liquid chromatography with tandem mass spectrometry (LC-MS/MS) analysis was used to determine the molecular source of α-secretase in purified CBS and aged blood serum (AgBS) fraction. Of the proteins identified, the subunits of C1 complex (C1q, C1r, and C1s) and alpha-2-macroglobulin showed significantly greater levels in purified α-CBS fraction (α-CBSF) compared with the AgBS fraction (AgBSF). Specifically, C1 markedly increased sAPPα and alpha-carboxyl-terminal fragment (α-CTF) production in a dose-dependent fashion, whereas C1q alone only minimally increased and C3 did not increase sAPPα production in the absence of sera. Furthermore, C1q markedly increased sAPPα and α-CTF, while decreasing Aβ, in CHO/APPwt cells cultured in the presence of whole sera. These results confirm our initial assumption that APP α-secretase activity in human blood serum is mediated by complement C1, opening a potential therapeutic modality for the future of AD.
Keywords
Introduction
Alzheimer’s Disease (AD) is the most common neurodegenerative disorder, characterized by the presence of intracellular neurofibrillary tangles, extracellular β-amyloid plaques, and neuronal loss throughout the entire brain. Aβ, a 4 kDa peptide, is one of the major components of β-amyloid plaques. Under physiological conditions, α-secretase cleaves APP within the Aβ domain, thereby preventing generation of Aβ and releasing soluble neurotrophic sAPPα
1
. sAPPα has been shown to reduce Aβ generation and plaque deposition by interfering with beta-site APP cleaving enzyme 1 (BACE1), also known as β-secretase
2,3
. In addition, sAPPα has also been shown to inhibit tau phosphorylation through modulation of the GSK3β signaling pathway
4
. In the neurodegenerative environment of AD, sequential cleavage of APP by β- and
Human umbilical cord blood cells (HUCBCs) have emerged as a potential immunomodulatory therapy in various diseases, including ischemia and neurodegeneration 8 . The many benefits of HUCBC include attenuation of amyloidogenic APP processing and reduction of Aβ levels in PSAPP and Tg2576 mice models of AD 9 . Importantly, multiple low-dose HUCBC infusion has also been shown to improve cognitive functions in transgenic PSAPP mice 10 . Our most recent work showed that cord blood serum (CBS) significantly promoted sAPPα production, whereas heat-inactivation eliminated this effect. Furthermore, sAPPα production was found to be increased by the removal of serum immunoglobulins and salts. To further understand what exactly in CBS was promoting sAPPα, we isolated the specific fraction of CBS found to have the highest α-secretase activity (α-CBSF) by a three-step chromatographic process. The proteolytic α-secretase activity in α-CBSF was found to not be mediated by tumor necrosis factor-α converting enzyme (TACE) or a disintegrin and metalloproteinase domain-containing protein (ADAM) 10 . Immunoprecipitation studies indicated the presence of an α-secretase-like enzyme that physically interacts with APP 11 . As first reported, following the administration of α-CBSF by intracerebroventricular (ICV) injection, both Aβ levels and tau phosphorylation were reduced in 3xTg-AD mice. Amyloid plaque numbers were decreased in the neocortex and hippocampus of 5XFAD mice through α-CBSF intraperitoneal minipump administration in comparison with mouse brain treated with the corresponding AgBSF. Furthermore, ICV administration of α-CBSF enhanced non-amyloidogenic APP processing in 3xTg-AD mice, providing a neuroprotective effect. These results combined with reduced neuronal loss in the neocortex of the 5XFAD mouse model following α-CBSF administration reveal a therapeutic potential for improving synaptic plasticity and neuroinflammation 11 .
The complement system is a major part of the innate immune system involved in neuroprotection via activation of immunomodulatory pathways 12 . In fact, a growing body of evidence suggests that complement proteins have neuroprotective effects in the developing and adult brain 13,14 . Neurogenesis and synapse elimination are heavily influenced by the complement system, particularly in the early stages of neurodevelopment 13,15 . Complement protein C1 complex is most commonly known for its neuroprotective role in enhancing and refining the neural network through phagocytosis of apoptotic cells and immune complexes 16 . The C1 complex is composed of a C1q and two each of C1r and C1s molecules. C1q, the largest subcomponent of the cascade initiator of the C1 pathway, is composed of six each of A, B, and C chains, comprising 18 polypeptide chains 17 . In addition to a role in synaptic elimination, complement protein subcomponent C1q has been shown to increase neuronal survival and branching in absence of other complement components, 18 and this effect could be mediated via upregulation of genes associated with neurotrophic factors (neural growth factor, and neurotrophin 3), cytoskeletal (syntaxin-3), and cholesterol metabolism (cholesterol-25-hydroxylase) 19 . During development, C1q is expressed in synaptic regions, and synapse elimination is mediated by C1q during the period of synaptic pruning. Therefore, deficiencies in C1q or downstream complement components can lead to neuronal defects and other detrimental effects 15 . These data confirm a direct neuroprotective role of C1q in the physiological condition. In peripheral phagocytes C1q downregulates pro-inflammatory cytokine expression and supports the overall clearance of cellular debris. C1q binds to apoptotic cells, flagging them to be cleared out before they release neurotoxic components 20 . Despite several physiological functions, dysregulated activation of the complement pathways in the central nervous system (CNS) have usually been considered detrimental in various conditions, including AD 21 . Treatment with a cyclic hexapeptide C5a receptor antagonist (PMX205) has been shown to reduce fibrillary amyloid deposition and glial activation, while rescuing cognitive impairment in a passive avoidance task in Tg2576 Swedish mutant mice 22 . Neuronal C1q is normally downregulated in adult CNS, but is found to be upregulated in the advent of injury or early stages of disease 13 . Interestingly, C1q is found to be induced in the brain in response to neuronal injuries in AD and blocks fibrillary forms of Aβ neurotoxicity in vitro 23 . Benoit et al. demonstrated that C1q protects both immature and mature primary neurons against fibrillary and oligomeric Aβ toxicity 24 . Depending on the timing and environment, C1q can either be neuroprotective or induce an inflammatory environment, leading to neurodegeneration in the adult brain. Loeffler et al. reported that C3b and iC3b (i.e., proteolytically inactive product of C3b which prevents complement cascade activation) are also accumulated on AD-affected neurons, much like C1q 25 .
Given that sAPPα has anti-amyloidogenic properties, the primary known α-secretases that cleave APP are nonspecific, and stimulating them could generate unwanted off-target effects. Our preliminary data show that α-secretase-like enzymatic activity in CBS is not mediated by ADAM10 or TACE, the major α-secretases that cleave APP in AD. In addition, we purified and identified a CBS-specific fraction α-CBSF, with more than 1000-fold increased α-secretase activity, which markedly increased cerebral sAPPα, decreased Aβ production and abnormal tau phosphorylation, and ameliorated cognitive impairment in two (3xTg-AD and 5XFAD) AD mouse models 11 . In support of this hypothesis, we have tested further for the identification of the heat-sensitive protein factor(s) exerting this novel enzymatic function. Here, we have recently identified a novel role for complement protein C1 complex in the CNS. Multi-step fractionation followed by liquid chromatography with tandem mass spectrometry (LC-MS/MS) analyses of CBS have identified several proteins compared to AgBS, each of which could potentially generate sAPPα by cleaving APP in the cellular system. Utilizing in vitro CHO/APPwt, N2a/APPwt, and TgAPPwt primary neuronal cell lines, we found that α-secretase-like enzymatic activity promoting sAPPα may be in part mediated by complement protein C1 complex. These results point to a novel neurotrophic function of complement protein C1 complex, which requires further investigation.
Materials and Methods
Reagents and Antibodies
CBS was obtained from Lee Biosolutions (St. Louis, MO, USA). CBS was separated from umbilical cord blood by allowing clotting for 5–10 h in tubes without anticoagulant followed by centrifugation at 3500 rpm for 5–10 min and filtration through a membrane with a pore size of 0.22 µm. Individual CBS samples were prepared from a single lot of cord blood, and more than 10 samples of CBS were combined as “pooled CBS.” Normal human aged blood serum (AgBS, >75 years old) were obtained from Florida Blood Services (Tampa, FL, USA). All sera were heat-activated. Heat-inactivation of human sera was conducted at 56°C for 30 min. Complement proteins, complement depleted sera, and complement inhibitors were acquired from the following sources. Human complement protein C1 inhibitor (Cat# GF178), complement protein C1 from human serum (Cat# 204873), complement protein C3-depleted human serum (Cat# 234403), and complement protein C1q-depleted human sera (Cat# 234401) from MilliporeSigma (St. Louis, MO, USA). Complement protein C3 from human serum (Cat# A113) and complement iC3b from human serum (Cat# A115) from Complement Technology, Inc. (Tyler, Texas, USA). Complement protein C1q from human serum (Cat# 204876) from MilliporeSigma (St. Louis, MO, USA). Compstatin, complement protein C3b inhibitor human (Cat# SB 431542) was acquired from Tocris Bioscience (Minneapolis, MN, USA). Sera from C1qa deficient (def) mice (129P2/OlaHsd, 3 months of age) were kindly provided by Dr. Andrea J. Tenner (University of California, Irvine, USA). Antibodies used in this studies include: specific anti-sAPPα monoclonal antibody (2B3; IBL, Minneapolis, MN, USA, Cat# 11088 RRID: AB_494690), anti-APP C-terminal polyclonal antibody (pAb751/770; EMD Millipore, La Jolla, CA, USA, Cat# 171610, RRID: AB_211444), anti-N-terminal Aβ monoclonal antibody (6E10; Covance Research Products, Emeryville, CA, USA), and anti-β-actin monoclonal antibody (Cat# A5316, RRID: AB_476743) from MilliporeSigma (St. Louis, MO, USA). The human sAPPα ELISA assay kit was purchased from IBL AMERICA, Minneapolis, MN, USA.
CBS and AgBS Fractionation
In order to purify and characterize the α-secretase in CBS or AgBS, the Econo-Pack Serum IgG purification kit, and 10 DG columns (Bio-Rad, Philadelphia, PA, USA) were initially employed to remove highly abundant IgG and salts. The desalted serum was applied to DEAE Affi-Gel blue columns and residual IgG was eluted according to the instructions. Then, 20 additional protein fractions were collected by eluting with an ionic strength gradient of NaCl buffer ranging from 0.1 M to 2.0 M. The remaining proteins on the column were eluted by the regeneration buffer included in the kit and collected as the regeneration fraction. The 0.8 M NaCl eluted protein fractions were combined and sent to Moffitt Cancer Center protein purification core for further separation by size-exclusion chromatography, employing analytic Superdex 200 columns and eluting with phosphate-buffered saline (PBS), and ion-exchange chromatography, employing Q-Sepharose columns and eluting with 500 mM NaCl, 50 mM Tris, pH 7.6. The final enzyme containing fractions were exchanged to PBS buffer by Ultracel-10 membranes (10 kDa, Merck Millipore, Burlington, MA, USA) for further experimentation and referred to as α-CBSF or AgBSF (Fig. 1).

Tentative workflow for α-CBSF and AgBSF fractionation. Initially, CBS and AgBS were desalted followed by sequential chromatographic separation using DEAE column, size-exclusion, and ion-exchange chromatography. Each fraction was collected followed by efficacy testing by treating CHO/APPwt cells for sAPPα release. Eventually a fraction (α-CBSF) was chosen based on the highest capacity for sAPPα generation. LC-MS/MS was performed to identify the source of the protein function. AgBS, aged blood serum; CBS, cord blood serum; DEAE, diethylaminoethyl; LC-MS/MS, liquid chromatography with tandem mass spectrometry; sAPPα, secreted amyloid precursor protein-α.
LC-MS/MS
Briefly, each lane gel slices were destained using acetonitrile and subjected to tris 2-carboxyethy-phosphine (TCEP) reduction and iodoacetamide alkylation followed by in-gel trypsin digestion, extraction, and vacuum centrifugation concentration. A nanoflow liquid chromatograph (U3000, Dionex, Sunnyvale, CA) coupled to an electrospray ion trap mass spectrometer (LTQ-Orbitrap, Thermo, San Jose, CA) was used for tandem mass spectrometry peptide sequencing experiments. A 5 µl sample was first loaded onto a pre-column (5 mm × 300 µm ID packed with C18 reversed-phase resin, 5 µm, 100 Å) and washed for 8 min with aqueous 2% acetonitrile and 0.04% trifluoroacetic acid. Next, the trapped peptides were eluted onto the analytical column (C18, 75 µm ID × 15 cm; Pepmap 100, Dionex, Sunnyvale, CA). The 120-min gradient was programmed as: 95% solvent A (2% acetonitrile + 0.1% formic acid) for 8 min; solvent B (90% acetonitrile + 0.1% formic acid) from 5% to 50% in 90 min; then solvent B from 50% to 90% in 7 min and held at 90% for 5 min; followed by solvent B from 90% to 5% in 1 min and re-equilibrate for 10 min (60-min gradient setup). The flow rate on the analytical column was 300 nl/min. Five tandem mass spectra were collected in a data-dependent fashion following each survey scan. The MS scans were performed in Orbitrap to obtain accurate peptide mass measurement and the MS/MS scans were performed in a linear ion trap using 60 s exclusion for previously sampled peptide peaks. The LC-MS/MS data were submitted to a local MASCOT server Swiss-Prot Human database for MS/MS protein identification search via ProteomeDiscoverer software. Two trypsin missed cleavages were allowed; the precursor mass tolerance was 1.08 Da. The MS/MS mass tolerance was 0.8 Da. The incorrect detection rate in each LC-MS/MS analysis was set to be less than 1%.
Cell Culture
Chinese hamster ovary (CHO) cell line with stable expression of human wild-type APP (CHO/APPwt) was a generous gift from Drs. Stefanie Hahn and Sascha Weggen (University of Heinrich Heine, Düsseldorf, Germany). These cells were initially genotyped to confirm their genetic make-up. The cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum (FBS), 1 mM sodium pyruvate, and 100 U/ml of penicillin/streptomycin. For treatment, the cells were plated in a 24-well plate at 2 × 10 5 cells/well for overnight incubation, washed, and treated with CBS, heat-inactivated (56°C, 30 min) CBS, AgBS, or α-CBSF, AgBSF in DMEM. After treatment, supernatants were collected and the cells were washed with ice-cold PBS three times and lysed with cell lysis buffer (20 mM Tris, pH 7.5, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% v/v Triton X-100, 2.5 mM sodium pyrophosphate, 1 mM β-glycerolphosphate, 1 mM Na3VO4, 1 µg/ml leupeptin, and 1 mM PMSF; Cell Signaling Technology, Danvers, MA, USA). Both cell supernatants and lysates were used for sAPPα analysis by ELISA. In addition, murine N2a cells transfected with APPwt (N2a/APPwt) and primary neuronal cells were obtained from cerebral cortices of TgAPPwt mouse embryos, between 15 and 17 days in utero, as described previously 26 . For TgAPPwt mouse-derived cortical neurons, cerebral cortices were isolated from TgAPPwt mice between 15 and 17 days in utero and mechanically dissociated in trypsin (0.25%) individually after incubation for 15 min at 37°C. Cells were collected after centrifugation at 1200 g, suspended in DMEM supplemented with 10% fetal calf serum, 10% horse serum, uridine (33.6 mg/ml; Sigma-Aldrich, St. Louis, MO, USA), and fluorodeoxyuridine (13.6 mg/ml, Sigma-Aldrich), and seeded in 24-well collagen-coated culture plates at 2.5 × 10 5 cells/well. After reaching confluence (∼70–80%) these cells were treated with purified human complement C1 at 0, 5, 10, or 20 µg/ml for 4 h, supernatants were collected for sAPPα measurement by ELISA and cells were washed with ice-cold PBS three times and lysed with cell lysis buffer followed by western blotting using 6E10 and pAb751/770 antibodies.
Transgenic APPwt Mice
Transgenic wild-type B6.Cg-Tg (PDGFB-APP) 5Lms/J strain (TgAPPwt) male and female mice were purchased from the Jackson Laboratory (Bar Harbor, ME, USA). All mice were housed in the Animal Facility in the College of Medicine at the University of South Florida and maintained on a 12 h light/12 h dark cycle at ambient temperature and humidity with ad libitum access to food and water. All animal experiments involving mice were performed in compliance with the US Department of Health and Human Services Guide for the Care and Use of Laboratory Animals and in accordance with the guidelines of the University of South Florida Institutional Animal Care and Use Committee (IACUC reference number: IS00000438).
Western Blot Analysis and ELISA
Western blot (WB) analyses and quantification were performed as previously described 11 . Briefly, the proteins from the cell-free suspensions, cell lysates, and homogenized tissue were electrophoretically separated using 10% bicine/tris gel (8 M urea) for proteins less than 5 kDa or 10% tris/SDS gels for larger proteins. Electrophoresed proteins were transferred to nitrocellulose membranes (Bio-Rad), washed and blocked for 1 h at room temperature in tris-buffered saline containing 5% (w/v) nonfat dry milk (TBS/NFDM). After blocking, membranes were hybridized overnight with various primary antibodies, washed, and incubated for 1 h with the appropriate HRP-conjugated secondary antibody in TBS/NFDM and developed using the luminol reagent (Thermo Fisher Scientific, Waltham, MA, USA). Soluble sAPPα in the media was quantified with the IBL human sAPPα colorimetric sandwich ELISA kit according to the manufacturer’s instructions.
Statistical Analysis
Comparison between two groups were performed by Student’s t test analysis. For more than two groups, one-way analysis of variance (ANOVA) followed by least significant difference (LSD) post hoc analysis was used to compare each other for statistical significance. Alpha was set at 0.05 for all analyses. All molecular analyses were repeated three times in parallel. Data are expressed as mean ± SEM. The statistical package for the social sciences release IBM SPSS 23.0 (IBM, Armonk, NY, USA) was used for all data analyses.
Results
Heat-Sensitive APP α-Secretase Activity in CBS
In order to identify the source of α-secretase in CBS, we initially confirmed that CBS possesses heat-labile α-secretase activity. CHO/APPwt cells were treated with CBS (2%) for 0, 5, 10, 15, 30, 60, and 120 min, followed by determination of sAPPα in conditioned medium by ELISA. CBS caused a time-dependent increase in sAPPα release from CHO/APPwt cells that was significantly greater than the control treatment (Fig. 2a; *P < 0.05, **P < 0.01, ***P < 0.001). Similarly, CHO/APPwt cells were treated for 2 h with 0, 0.25, 0.5, 1, 2, and 5% CBS, followed by determination of sAPPα in conditioned medium by ELISA. CBS caused dose-dependent release of secreted amyloid precursor protein-α (sAPPα) in culture medium from CHO/APPwt cells that was significantly greater than the control treatment (Fig. 2b; *P < 0.05, **P < 0.01, ***P < 0.001). Next, we wanted to determine whether heat-inactivation of serum reduces the sAPPα-producing capacity of CBS. CHO/APPwt cells were treated with 2% whole or heat-inactivated CBS or AgBS (56°C for 30 min) for 2 h followed by determination of sAPPα in conditioned media by ELISA. Both CBS and AgBS heat-inactivation significantly limited sAPPα-releasing capacity (Fig. 2c; *P < 0.05, **P < 0.01, ***P < 0.001). Furthermore, CHO/APPwt cells were treated with 2% whole or heat-inactivated α-CBSF or AgBSF for 2 h followed by determination of sAPPα in conditioned media by ELISA (Fig. 2d; *P < 0.05, **P < 0.01, ***P < 0.001). As shown most recently, purification significantly enhanced the sAPPα-producing capacity of CBS and AgBS, which again was significantly limited by heat-inactivation. These results confirm that CBS possesses an enhanced level of heat-sensitive α-secretase activity.

CBS significantly enhances APP α-cleavage in CHO/APPwt cells. (a) CHO cells overexpressing human wild-type APP (CHO/APPwt cells) were treated for 0–120 min with CBS (2%). (b) CHO/APPwt cells were treated for 2 h with 0, 0.25, 0.5, 1, 2, and 5% CBS. (c) CHO/APPwt cells were treated for 2 h with PBS (0% serum, Ctrl), heat-inactivated CBS (Heat CBS), heat-inactivated AgBS (Heat AgBS), AgBS, or CBS at 2%. (d) CHO/APPwt cells were treated for 2 h with PBS (0% serum, Ctrl), heat-inactivated α-CBSF (Heat αCBSF), heat-inactivated AgBSF (Heat AgBSF), AgBSF, or α-CBSF at 2%, each purified as briefly described under “Materials and Methods.” Release of sAPPα into the conditioned medium after treatment was analyzed by ELISA. Data are presented as mean (±SEM) of sAPPα produced (ng/ml or ng/mg) from three independent experiments performed in triplicate. Significant differences from control treatment are indicated by asterisks (*P < 0.05, **P < 0.01, ***P < 0.001). AgBSF, aged blood serum fraction; CBS, cord blood serum; PBS, phosphate-buffered saline; sAPPα, secreted amyloid precursor protein-α; SEM, standard error of the mean.
Proteomic Analyses of CBSF and AgBSF
Having shown that CBSF possesses enhanced α-secretase activity, we next set out to determine the molecular source of α-secretase in CBSF and AgBSF by LC-MS/MS analysis. From 142 major proteins identified, we selected those most likely to exhibit α-secretase activity. Several of the major proteins identified in α-CBSF and AgBSF are shown in Fig. 3. Of these proteins, the subunits of complement protein C1 complex (C1q, C1r, and C1s) and alpha-2-macroglobulin showed significantly greater levels in α-CBSF compared with AgBSF (Fig. 3; *P < 0.05, **P < 0.01). Notably, the levels of complement proteins C3, C4, C5, C6, and C8 complexes, ApoE, ApoB100, and full-length APP (fAPP) were not significantly different between α-CBSF and AgBSF (P > 0.05). Complement proteins C1 and C3b complex have proteolytic activity, which can be inactivated at 56°C for 30 min. These proteins play important roles in neurodevelopment and provide early protection against stress or disease, including AD. Thus, we hypothesized that the heat-labile α-secretase activity in CBS is mediated in part by either complement protein C1 complex or C3b.
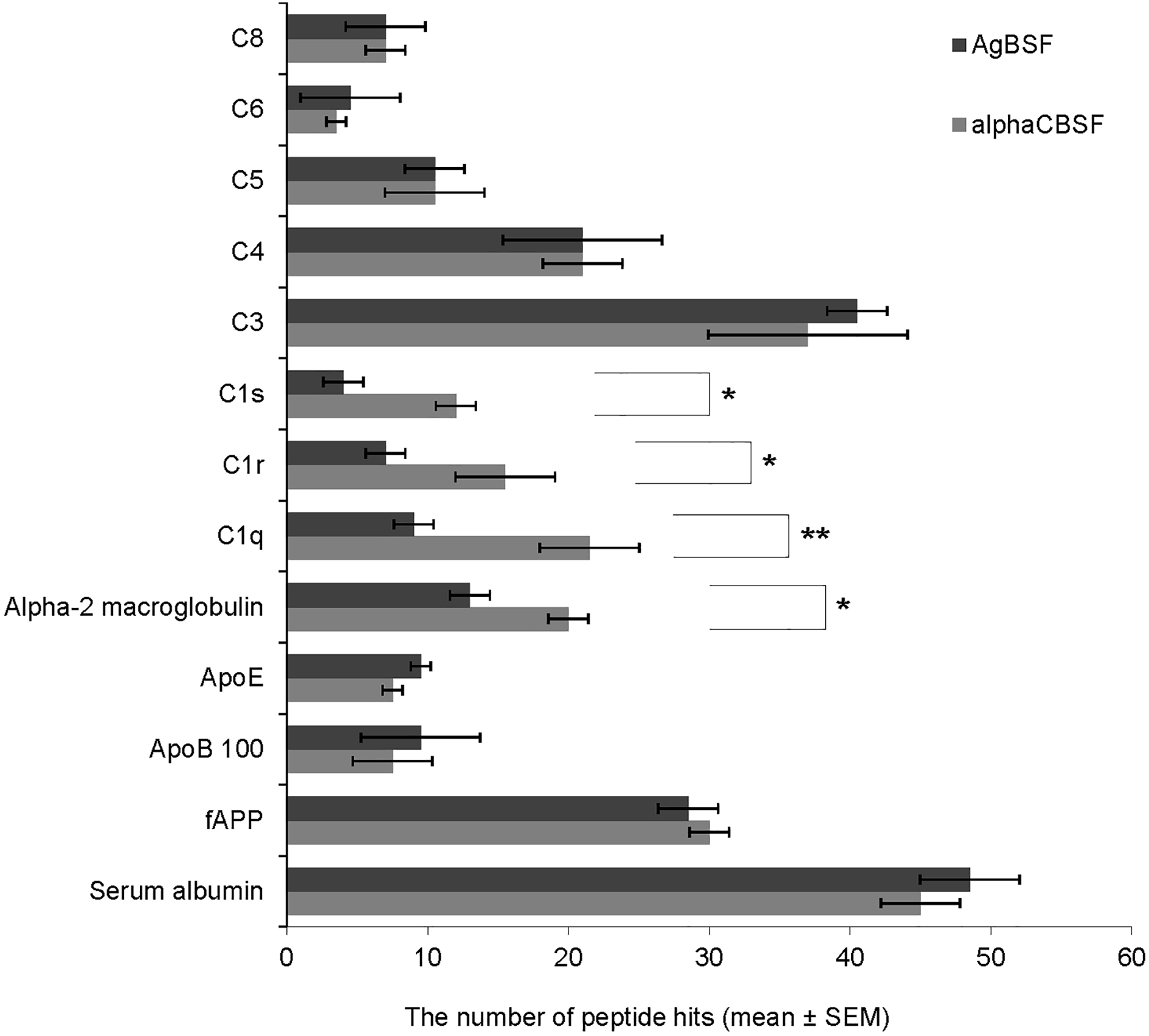
Identification of proteomic profile of α-CBSF and AgBSF. The proteome of α-CBSF and AgBSF were analyzed by LC-MS/MS. From 142 major proteins identified, those exhibiting proteolytic and therefore potential α-secretase-like activity were selected. Of these proteins, the subunits of complement protein C1q (**P < 0.01), C1r (*P < 0.05), and C1s (*P < 0.05), and alpha-2-macroglobulin (*P < 0.05) showed significantly greater levels in α-CBSF compared with AgBSF. Data are presented as mean (±SEM) of number of peptide hits from three independent experiments. Significant difference between AgBSF and α-CBSF indicated by asterisk (*P < 0.05, **P < 0.001, ***P < 0.01). AgBSF, aged blood serum fraction; LC-MS/MS, liquid chromatography with tandem mass spectrometry; SEM, standard error of the mean.
CBS-Mediated α-Secretase-Like Cleavage of APP is Independent of Complement C3b
In order to determine whether α-secretase activity of CBS is mediated by activation of complement protein fragment C3b, we investigated whether depletion of complement protein C3 could limit the α-secretase-like activity of CBS. CHO/APPwt cells were treated with C3-depleted CBS (2%) for 0, 15, 30, 60, and 120 min, followed by determination of sAPPα in conditioned medium by ELISA. Depletion of C3 complement protein did not change sAPPα-producing capacity of CBS in CHO/APPwt cells (Fig. 4d; *P < 0.05, **P < 0.01, ***P < 0.001). In order to confirm that C3b complement does not mediate α-secretase activity in CBS, we preincubated whole or C3-depleted CBS (2%) with the C3b inhibitor compstatin at 0, 6, 12, 25, 50, and 100 µM for 15 min at 37°C, and then treated CHO/APPwt cells with these premix complexes separately for 2 h. Compstatin did not alter sAPPα production significantly elicited by whole (Fig. 4a; P > 0.05) or C3-depleted serum (Fig. 4b; P > 0.05). Therefore, CBS α-secretase-like activity is independent of enzymatic activity of complement C3b protein.

CBS-mediated sAPPα release by CHO/APPwt cells is independent of complement protein C3b. C3b inhibitor compstatin at 0, 6, 12, 25, 50, and 100 µM was preincubated with CBS (2%) as well as C3-depleted CBS (2%) for 15 min. (a). CHO/APPwt cells were treated for 2 h with CBS (2%) + compstatin. (b) CHO/APPwt cells were treated for 2 h with C3-depleted CBS (2%) + compstatin. (c) In addition, CBS (2%) was preincubated with iC3b at indicated concentrations (0, 1.25, 2.5, 5, 10, and 20 µg/ml). CHO/APPwt cells were treated for 2 h with CBS + iC3b. (d). CHO/APPwt cells were treated with C3-depleted CBS (2%) for 0, 15, 30, 60, and 120 min. Release of sAPPα into the conditioned medium after treatment was analyzed by ELISA. Data are presented as mean (±SEM) of sAPPα produced (ng/mg) from three independent experiments performed in triplicate. Significant differences from control treatment indicated by asterisk (*P < 0.05, **P < 0.01, ***P < 0.001). CBS, cord blood serum; ELISA, enzyme-linked immunosorbent assay; sAPPα, secreted amyloid precursor protein-α; SEM, standard error of the mean.
In order to determine whether complement protein component C3b acts as an inhibitor of CBS α-secretase-like activity, CHO/APPwt cells were treated for 2 h with premixes of CBS (2%) and iC3b (0, 1.25, 2.5, 5, 10, and 20 µg/ml), the proteolytically inactive product of C3b that prevents complement cascade activation. The sAPPα-producing capacity of CBS was not altered by iC3b as determined by sAPPα ELISA (Fig. 4c; P > 0.05), indicating that complement C3b protein does not act as a potent CBS α-secretase inhibitor.
CBS α-Secretase-Like Activity is Mediated in Part by Complement Protein C1 Complex
To determine whether α-secretase-like activity in CBS is mediated by complement protein C1 complex, we determined CBS α-secretase activity in the presence of complement protein C1 inhibitor. CHO/APPwt cells were treated with α-CBSF (1 and 2%) in the presence of complement protein C1 inhibitor at 0, 50, 100, and 200 µg/ml for 2 h. C1 inhibitor markedly reduced sAPPα released by α-CBSF (1 and 2%) in a dose-dependent fashion, as determined by ELISA (Fig. 5a; *P < 0.05, **P < 0.01, ***P < 0.001) and WB (2% CBS; Fig. 5b; *P < 0.05, **P < 0.01, ***P < 0.001; quantification not shown), indicating that α-secretase-like activity in CBS is mediated in part by complement C1 protein. In addition, we determined the effect of purified human C1 complement protein on sAPPα production in N2a/APPwt and TgAPPwt/primary neuronal cells in the absence of sera. Complement protein C1 dose-dependently increased sAPPα and α-CTF production in N2a/APPwt cells (Fig. 5c–e; **P < 0.01, ***P < 0.001) and TgAPPwt/primary neuronal cells (Fig. 5f–h; *P < 0.05, **P < 0.01) in the absence of serum, as determined by ELISA and WB. These results indicate that complement protein C1 complex mediates α-secretase-like activity in CBS.

A specific complement protein C1 inhibitor significantly reduces APP α-cleavage induced by CBS. (a) CHO/APPwt cells were treated for 2 h with α-CBSF (1 and 2%) in the presence of C1 inhibitor at 0, 50, 100, and 200 μg/ml, followed by analysis of sAPPα in conditioned media by ELISA. (b) In addition, CHO/APPwt cells were treated for 2 h with α-CBSF (2%) in the presence of complement protein C1 inhibitor at 0–200 μg/ml, followed by analysis of sAPPα levels by WB. N2a/APPwt cells (c) and TgAPPwt/mouse primary neuronal cells (f) were treated with purified human complement protein C1 complex at 0, 5, 10, and 20 µg/ml in the serum-free condition for 4 h followed by analysis of holo APP, sAPPα and α-CTF in cell lysates by WB (c,f). The levels of sAPPα in conditioned media was also determined by ELISA
In order to confirm that complement C1 but not C3 protein has α-secretase-like activity, CHO/APPwt cells were treated with purified human complement C1 or C3, or C1q subunit, at 0, 2.5, 5, 10, and 20 µg/ml in serum-free media for 2 h followed by determination of sAPPα in conditioned medium by ELISA. As expected, complement protein C1 significantly increased sAPPα production in a dose-dependent fashion, whereas C1q alone minimally increased and complement protein C3 did not increase sAPPα production in serum-free medium (Fig. 6a; *P < 0.05, **P < 0.01). In contrast, both complement C1 and C1q protein component significantly increased sAPPα production in CHO/APPwt cells cultured in C1q-depleted sera (Fig. 6b; **P < 0.01, ***P < 0.001), and complement protein C1 significantly increased sAPPα and α-CTF level, while decreasing Aβ and β-CTF level in CHO/APPwt cells in the presence of whole sera as determined by WB (Fig. 6c).
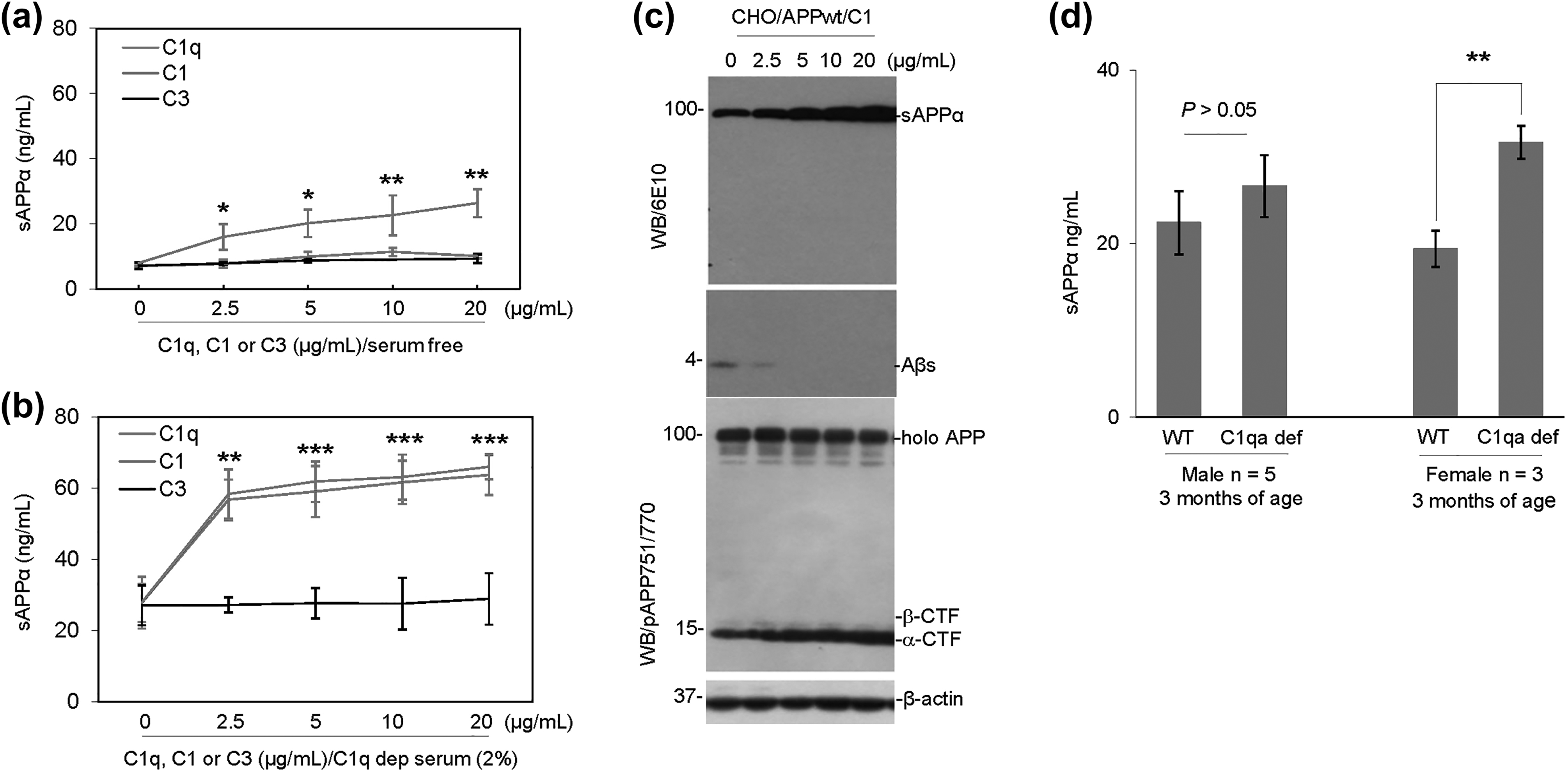
The purified human complement protein C1 complex promotes APP α-cleavage. (a) CHO/APPwt cells were treated for 2 h with purified human complement C1q subcomponent, C1 or C3, at 0, 2.5, 5, 10, and 20 µg/ml in serum-free media followed by analysis of sAPPα in conditioned media by ELISA. (b) CHO/APPwt cells were treated for 2 h with purified human complement C1q subcomponent, C1, and C3, at 0, 2.5, 5, 10, and 20 µg/ml in C1q-depleted serum followed by analysis of sAPPα in conditioned media by ELISA. (c) In order to further determine the effect of complement protein C1 on APP processing, CHO/APPwt cells were treated for 2 h with purified complement protein C1 at 0, 2.5, 5, 10, and 20 µg/ml in the absence of whole serum and cell lysates were prepared from each treated cell culture for analysis of holo APP, sAPPα, Aβ, and α- and β-CTF by WB. (d) CHO/APPwt cells were treated for 2 h with plasma (2%) obtained from C1qa-deficient mice or wild-type littermates at 3 months of age (n = 5 males or 3 females for each group) followed by determination of sAPPα in conditioned media by ELISA. For (a) and (b), data presented as mean (±SEM) of sAPPα produced (ng/ml) from three independent experiments performed in triplicate. Significant differences from control treatment indicated by asterisk (*P < 0.05, **P < 0.01, ***P < 0.001).
In order to investigate the role of complement protein C1 in mediating α-secretase activity in whole blood plasma, CHO/APPwt cells were treated with plasma from C1qa-deficient mice or age-matched wild-type littermates. Interestingly, the production of sAPPα elicited by plasma from male C1qa-deficient mice was not significantly different (Fig. 6d; P > 0.05), whereas that elicited by plasma from female C1qa-deficient mice was significantly greater than that elicited by age- and gender-matched plasma from wild-type littermates (Fig. 6d; **P < 0.01). Therefore, plasma α-secretase activity is not significantly altered by C1qa deficiency in male mice but significantly enhanced by C1qa deficiency in female mice. These results are consistent with C1-mediated α-secretase playing an important role in male and female individuals, but with other α-secretases compensating in its absence. In addition, the elevated sAPPα production capacity in female compared to male C1qa-deficient mice indicate an unknown function of this neurotrophic fragment in the peripheral system of females.
Altogether, these results confirm that APP α-secretase-like activity in human blood serum is mediated by complement protein C1 complex. The inability of C1q to rescue α-secretase activity in serum-free media is likely due to the absence of components C1s and C1r, which are required to complex with C1q in C1 for proteolytic activity.
Discussion
In our previous study, we have shown that peripheral administration of HUCBC 9 , as well as HUCBC-derived monocytes 27 , reduce cognitive impairments and β-amyloid plaques in PSAPP AD mice. In a subsequent study, we showed that human CBS also has α-secretase-like activity that likewise reduces cognitive impairment, tau hyperphosphorylation, and plaque deposition in AD mice 11,28 . The present study further investigated the component of CBS that attenuates β-amyloid plaques, especially through the increased production of the neurotrophic sAPPα. Our results confirm the presence of α-secretase-like enzyme activity within the CBS, reflected by elevated production of sAPPα. Interestingly, heat-inactivation of α-secretase activity of CBS at 56°C for 30 min inspired us to assume that α-secretase-like activity is mediated by a complement protein. This is further reinforced by the result of LC-MS/MS data, which indicate the dominance of complement protein C1 subcomponents (C1q, C1r, and C1s), C3b, and C4 proteins in α-CBSF as well as AgBSF (Fig. 3). Further investigations using complement inhibitors and C1 complement protein-depleted serum reveal that endogenous C1 complex enhances sAPPα production in cell culture and primary neuronal cell line, including CHO/APPwt (Fig. 5b), N2a/APPwt (Fig. 5c–e), and TgAPPwt/primary neuronal cells (Fig. 5f–h). As previously mentioned, complement protein C1 is composed of a C1q, and two each of C1r and C1s. C1q is composed of six each of A, B, and C chains, comprising 18 polypeptide chains 17 . Modifying various factors revealed that C1q depleted serum elicited less sAPPα production in comparison with the whole serum, highlighting the crucial nature of C1q-mediated α-secretase activity in CBS (Fig. 6a–c). Interestingly, our results indicate that the inability of C1q to rescue α-secretase-like activity in serum-free media is likely due to the absence of C1s and C1r subcomponents, which are required to form complexes with C1q in C1 for proteolytic activity (Fig. 6a). It should be noted that C1r and C1s are serine proteases that form the complete initiation complex of C1qC1r2C1s2 when activated by C1q binding to an immune complex 29 .
Initially, C1q synthesis serves to be protective by aiding in clearance of apoptotic cells and regulating the environment to prevent further neurotoxicity from spreading 23 . It has been proposed that C1q and C3 opsonization 30,31 accelerate clearance of Aβ. In addition, C1q-mediated protection against Aβ is associated with expression of low-density lipoprotein receptor-related protein 1B and G-protein-coupled receptor 6 as early as 2 months of age in the 3xTg-AD mouse model 24 . Despite the critical role C1q plays in immune regulation and neuroprotection, evidence has also suggested that C1q could potentially be proinflammatory 32 . C1q is synthesized when there is an injury in the CNS 33 . By binding with β-sheet fibrillary plaques, C1q activates the complement cascade that can have detrimental inflammatory consequences via production of the chemotactic factor C5a and recruitment and activation of microglial cells to the site of injury. This activation requires association of C1q with C1r and C1s as in the C1 complex. As the disease pathology continues to progress, it causes more of the complement pathways to be activated 34 . High density of negative charges in various regions can activate the complement system through an electrostatic interaction independent of antibodies 35 . This can be seen with β-amyloid fibrils binding to arginine-rich segments within the β-chain of C1q 36 . Consequently, the fragments not only create further inflammation, attracting glial cells, but can also activate the classical pathway, triggering the release of more pro-inflammatory components. In a study done with APPQ–/– mice, those lacking C1q were unable to activate the classical pathway because the fibrillary Aβ interaction with C1q is not possible 37 . In AD pathology, factors leading to neurodegenerative pathways of complement activation include C1q interaction with not only the β-sheet fibrils, but also hyperphosphorylated tau 35 . Though these fibrils would normally be cleared by microglia, binding of C1q to Aβ1-42 peptide strands makes it increasingly difficult for microglia to clear out neurotoxic debris 38 . The general trend toward increased complement activation causes the activated microglia to further exacerbate inflammation as C1q upregulation positively correlates with the quantity of reactive astrocytes, making the blood–brain barrier vulnerable 28 .
Collectively, our results indicate that CBS contains proteins that promote α-secretase-like enzymatic activity (Fig. 2). LC-MS/MS analysis in CBSF and AgBSF revealed the presence of 142 proteins, of which complement protein C1 subunits and alpha-2-macroglobulin showed significantly greater levels in α-CBSF compared with AgBSF (Fig. 3). Upon further study, complement protein C1 subunits showed enhanced sAPPα production and Aβ reduction in cell culture studies (Figs. 5 and 6). As discussed previously, C1q is known to have pro-inflammatory and detrimental effects on neuronal tissue, which will limit the therapeutic application of C1q. There may be other components in α-CBSF contributing to the beneficial effects observed in transgenic AD mice. In addition, C1 has α-secretase activity, which may also compete with the amyloidogenic β-secretase pathway and reduce β-cleavage of APP when α-CBSF is administered in mice. There is much to be learned about the exact mechanisms of action of C1q, especially how its neurotrophic effects may shift in AD pathology and instead lead to further acceleration of neuro-detrimental inflammation. Nevertheless, our research identified an additional and novel function in CBS by enhancing α-secretase-like activity and that the overall reduction in β-amyloid plaques incites a therapeutic potential for attenuating AD pathology.
Footnotes
Authors’ Note
Ahsan Habib, Darrell Sawmiller, and Huayan Hou contributed equally to this work.
Acknowledgments
We would like to thank Dr. Andrea Tenner for kindly providing us with murine C1qa-deficient blood sera and Dr. Robert Sprung for his technical support in size-exclusion and anion-exchange chromatography techniques. We would like to thank Dr. R. Doug Shytle for his helpful discussion.
Ethical Approval
All animal experiments involving mice were performed in compliance with the US Department of Health and Human Services Guide for the Care and Use of Laboratory Animals and in accordance with the guidelines of the University of South Florida Institutional Animal Care and Use Committee.
Statement of Human and Animal Rights
All animal experiments were performed in accordance with the guidelines of the National Institutes of Health and were approved by University of South Florida (USF) Institutional Animal Care and Use Committee (IACUC reference number: IS00000438). In addition, all mice were maintained on a 12-h light/12-h dark cycle at ambient temperature and humidity and housed in the Morsani College of Medicine Animal Facility at USF with ad libitum access to food and water.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
PRS is a cofounder and JT is a consultant for Saneron CCEL Therapeutics, Inc. JT and PRS are inventors on a patent application submitted by the University of South Florida. PRS was not involved in any data acquisition and analysis. All other authors report no biomedical financial interests or potential conflicts of interest.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the NIH/NIA (R21AG055116, JT and R01AG050253) and the Silver Endowment (JT).