
Abstract
Long-term graft survival is an ongoing challenge in the field of islet transplantation. With the growing demand for transplantable organs, therapies to improve organ quality and reduce the incidence of graft dysfunction are of paramount importance. We evaluated the protective role of a recombinant DNA repair protein targeted to mitochondria (Exscien I-III), as a therapeutic agent using a rodent model of pancreatic islet transplantation. We first investigated the effect of therapy on isolated rat islets cultured with pro-inflammatory cytokines (interleukin-1 β, interferon γ, and tumor necrosis factor α) for 48 h and documented a significant reduction in apoptosis by flow cytometry, improved viability by immunofluorescence, and conserved functional potency in vitro and in vivo in Exscien I-III-treated islets. We then tested the effect of therapy in systemic inflammation using a rat model of donor brain death (BD) sustained for a 6-h period. Donor rats were allocated to 4 groups: (non-BD + vehicle, non-BD + Exscien I-III, BD + vehicle, and BD + Exscien I-III) and treated with Exscien I-III (4 mg/kg) or vehicle 30 min after BD induction. Sham (non-BD)-operated animals receiving either Exscien I-III or vehicle served as controls. Islets purified from BD + Exscien I-III-treated donors showed a significant increase in glucose-stimulated insulin release in vitro when compared to islets from vehicle-treated counterparts. In addition, donor treatment with Exscien I-III attenuated the effects of BD and significantly improved the functional potency of transplanted islets in vivo. Our data indicate that mitochondrially targeted antioxidant therapy is a novel strategy to protect pancreas and islet quality from the deleterious effects of cytokines in culture and during the inflammatory response associated with donation after BD. The potential for rapid translation into clinical practice makes Exscien I-III an attractive therapeutic option for the management of brain-dead donors or as an additive to islets in culture after isolation setting.
Introduction
The majority of transplantable organs in the United States are recovered from brain-dead donors, and graft survival rates for these organs remain significantly inferior to those from living donors. 1 In the field of pancreatic islet transplantation as a therapy for type 1 diabetes mellitus (T1DM), success continues to be limited by the quality of the donor organ, the efficiency of the islet isolation process, the progressive loss of islet function over time, and the need for multiple donors to achieve insulin independence. 2,3 Currently, it is accepted that organs from brain-dead donors experience a cascade of nonspecific inflammatory events characterized by systemic release of pro-inflammatory cytokines such as tumor necrosis factor α (TNF-α), interleukin-1 β (IL-1β), and interferon γ (IFN-γ) that promote oxidative stress and ultimately lead to cellular injury. 4,5 The inflammatory response along with an endogenous overproduction and release of reactive oxygen species (ROS) has been suggested as a primary mechanism leading to pancreatic cellular injury and primary islet graft dysfunction. 6 –8 Inflammation can be triggered by multiple factors including pancreas donation after brain death (BD), cold ischemia during organ preservation, mechanical and enzymatic stress during islet isolation, and postinfusion activation of recipient innate immune and coagulation cascades known as instant blood-mediated inflammatory reaction (IBMIR). IBMIR is characterized by the activation of the host complement and coagulation cascades and infiltration of the graft by recipient leukocytes, resulting in the formation of cytotoxic ROS. 9 A variety of antioxidant, anti-inflammatory, and immunosuppressive mediators have been evaluated for improving islet quality and long-term graft survival after islet transplantation, but an optimal regimen of immunosuppression and islet support has yet to be established. 10,11 In the past, the overexpression of antioxidant enzymes has been shown to confer protection in insulinoma cell lines against the toxic effects of ROS, nitric oxide (NO), pro-inflammatory cytokines (IL-1β, TNF-α, and IFN-γ). 12 –15 Excessive generation of ROS has been also shown to interfere with adenosine triphosphate (ATP) generation and cellular capacity to repair damaged DNA. The mitochondrial genome encodes proteins vitally related to ATP generation through the electron transport chain. It is also particularly sensitive to oxidative damage, and ROS promotes oxidation of the phosphate/sugar backbone and bases in mitochondrial DNA (mtDNA). Oxidative damage to mtDNA is generally repaired by the mitochondrial base excision repair machinery, but capacity can be easily overwhelmed by an intense ROS burst in tissues such as pancreatic β cells that are especially susceptible due to lower levels of endogenous antioxidant enzymes. 16,17 Mitochondrial disruption allows increased amounts of endogenous ROS to leak into the cytoplasm and activates oxidases to produce more ROS, triggering a feedback loop of mtDNA damage and mitochondrial dysfunction that ultimately results in cellular apoptosis, necrosis, and/or autophagy. 18 Repair of mtDNA involves glycosylase and lyase activity that cleaves the sugar/phosphate backbone at apurinic/apyrimidinic (AP) sites on the 3′ side of the lesion. 19 Evidence suggests that the overexpression of the nuclear glycosylase/AP lyase, 8-oxoguanine DNA glycosylase (Ogg1), in the mitochondrial matrix protects cells from oxidative damage induced by oxidizing agents such as hydrogen peroxide or NO. 20,21 In vivo, selective overexpression of mitochondrial Ogg1 has been shown to increase viability of pulmonary artery endothelium exposed to ROS derived from xanthine oxidase enzymatic activity indicating a central role for lyase activity in mtDNA repair after oxidative damage. 22 Moreover, increasing mitochondrial Ogg1 expression and activity has been associated with a protective effect in human lung adenocarcinoma cells exposed to ROS and in models of ventilatory injury of the lung. 23 Another glycosylase able to target oxidized pyrimidines is the bacterial DNA glycosylase, Endonuclease III, which shares a common lyase activity with Ogg1 in cleavage of the AP sites on the 3′ side of the lesion. Targeted delivery of Endonuclease III to mitochondria of HeLa and breast cancer cells has been shown to promote mtDNA repair and increase cellular resistance to oxidative stress. 24,25 Exscien I-III is a novel mitochondrially targeted fusion protein engineered to allow its rapid uptake into the mitochondrial matrix, enhance the repair of mtDNA, and preserve cellular viability. Multiple studies have demonstrated that mitochondrially targeted Endonuclease III can significantly reduce mtDNA. 24,26 In the present study, we investigated the therapeutic impact of Exscien I-III on isolated pancreatic islets in vitro and in vivo. For the in vitro experiments, we supplemented Exscien I-III to isolated islets in culture. For the in vivo portion of our study, we evaluated the effect of pretreatment with Exscien I-III in a model of BD in rats followed by xenogeneic islet transplantation into immunodeficient diabetic-recipient mice. Our results indicate that pretreatment of BD donors with Exscien I-III and supplementation of Exscien I-III in culture has the potential to attenuate the deleterious effects of pro-oxidant cytokines on pancreatic islet cells and improve the viability and functional potency of isolated islets posttransplant.
Materials and Methods
Chemicals
Exscien I-III is a novel fusion protein construct developed at the University of South Alabama (Patent WO 2009099679 A2) and provided by the Exscien Corporation (Louisville, KY, USA). The fusion protein includes a mitochondrial targeting sequence and a protein transduction domain attached to Endonuclease III coupled to a TAT sequence to facilitate cellular uptake, a hemagglutinin tag and a histidine tail for detection and purification. 25 Purified protein was transported on dry ice and stored at −80 °C in our laboratory until use. Vehicle solution was provided by the Wilson Laboratory at the University of South Alabama. Recombinant human interleukin-1 β (IL-1β), tumor necrosis factor α (TNF-α), and interferon-γ (IFN-γ) were purchased from R&D Systems, Inc. (Minneapolis, MN, USA).
Animals
Inbred male Lewis rats (Harlan Sprague-Dawley, Indianapolis, IN, USA) weighing 250 to 400 g were used as donors and were arranged into treatment groups according to experimental conditions (Fig. 1). For the in vivo experiments, donor animals were divided into 4 groups: (1) Sham (non-BD) + vehicle (n = 6), (2) Sham (non-BD) + Exscien I-III (n = 3), (3) BD + vehicle (n = 6), and (4) BD + Exscien I-III (n = 4). For the in vitro portion of the experiments, 4 naive rats were used for each preparation (total 60 rats) and cultured islets that were then split up into different culture groups: (1) control + vehicle, (2) control + Exscien I-III, (3) cytokines, and (4) cytokines + Exscien I-III. Inbred male non-obese diabetic/severe combined immunodeficiency (NOD SCID) mice (Jackson Laboratory, Bar Harbor, MA, USA) were used as diabetic recipients in applicable experiments. For the in vivo experiments, transplanted animals were divided into the following groups: (1) Sham (non-BD) + vehicle (n = 10 recipients), (2) Sham (non-BD) + Exscien I-III (n = 8 recipients), (3) BD + vehicle (n = 9 recipients), and (4) BD + Exscien I-III (n = 8 recipients). For the in vitro experiments, recipients were divided as follows: control + vehicle (n = 16), control + Exscien I-III (n = 13), cytokines + vehicle (n = 15), and cytokines + Exscien I-III (n = 17). All experiments were performed in accordance with the guidelines established and approved by the University of Wisconsin-Madison Institutional Animal Care and Use committee.

Experimental design.
Rat Islet Isolation and Culture
Donor animals were anesthetized with a mixture of isoflurane (0.5%-2%) in oxygen, and the abdominal cavity was entered via midline incision. A polyethylene catheter (PE50, Braintree Scientific, MA, USA) was introduced into the abdominal aorta, the animal was exsanguinated and the organs flushed in situ with 30 mL of cold (4 °C) normal saline. The common bile duct was cannulated and injected with digestion solution–containing collagenase type V (Sigma-Aldrich Corp., St Louis, MO, USA) and DNase (Roche Diagnostics, Indianapolis, IN, USA). The pancreas was excised, and islets were isolated and purified by centrifugation on a discontinuous Ficoll gradient (Mediatech, Inc., Manassas, VA, USA). Purified islets were cultured at 37 °C and 5% CO2 in fetal-bovine serum (FBS)-supplemented CMRL1066 media (Mediatech, Inc.) and divided into 4 groups: (1) Control islets + vehicle, (2) Control Islets + Exscien I-III, (3) Cytokines + vehicle, and (4) cytokines + Exscien I-III. The islets were then cultured for a total of 48 h with media changed at 24 h postisolation.
Cytokines were supplemented at the following doses: TNF-α 1,000 U/mL, IFN-γ 1,000 U/mL, and IL-1β 50 U/mL. Exscien I-III was supplemented at a dose of 6.7 μg/mL, and vehicle control was supplemented at a matching volume. After 48 h, islets were washed, and quality parameters were assessed.
Brain Death Induction
Donor rats were anesthetized by a mixture of 0.5% to 2% isoflurane in oxygen using a precision vaporizer. An oral endotracheal tube was placed and connected to a mechanical ventilator for 6 h (Kent Scientific, Torrington, CT, USA). Blood–oxygen saturation was monitored via pulse oximeter on the left hind foot-pad, heart rate, and mean arterial pressure were monitored via PE50 catheter placed in the right femoral artery and connected to a transducer and recorder (SurgiVet, Smiths Medical, Waukesha, WI, USA). Hydration was supplied continually as normal saline injected via syringe-pump (0.25to1.0 mL/hr; Medfusion 2010i, Smiths Medical, Hythe, UK) through a PE50 catheter placed in the right femoral vein. Body temperature was maintained by a hot-water blanket (Gaymar TP-500, Orchard Park, NY, USA) and monitored with a rectal probe (Mon-a-therm Model 6510, Mallinckrodt Medical, St Louis, MO, USA). The parietal bone was exposed via a midline skin incision and an occipital burr hole to the subdural space. BD was induced via gradual inflation of a 4F balloon catheter placed through the hole to the subdural space, increasing intracranial pressure. The catheter was inflated to 0.5 ± 0.1 mL over the course of a 5-min period and maintained for the 6-h experimental period. BD was confirmed by the absence of central reflexes, maximally dilated and fixed pupils, and a positive apnea test. Nonbrain dead (sham) ventilated donors were used as controls in applicable experiments. After 30 min of mechanical ventilation, either vehicle or Exscien I-III (4 mg/kg) was delivered as a 0.5 mL bolus intravenously through the right femoral vein catheter. After 6 h, anesthetized animals were subjected to a laparotomy, and blood was collected from the abdominal aorta. Organs were then perfused with a flush of 30 mL cold normal saline in situ through the abdominal aorta allowing effusion through an incision in the vena cava. Islet isolation was performed on donor pancreata as described above. Purified islets were then cultured at 37 °C and 5% CO2 in FBS-supplemented CMRL1066 media (Mediatech, Inc.) for 24 h.
Viability Assessment with Fluorescent Dyes
An aliquot of cultured islets was transferred in phosphate-buffered saline (PBS) to 10 × 35 mm counting Petri dishes and stained in a solution of fluorescein diacetate (FDA; Sigma-Aldrich) as a fluorescent indicator of cellular viability and propidium iodide (PI; Thermo Scientific) as an indicator of necrosis. Stained islets were imaged under a fluorescent microscope assessed for cell viability by estimating the percentage of viable cells (green) versus the percentage of nonviable cells (red) within each islet. Each preparation was evaluated separately by 2 qualified and independent investigators. The average percentage of viable cells was then calculated.
Multiparametric Flow Cytometry
Islets were dispersed into single-cell suspensions via incubation, with 0.05% Trypsin-ethylenediaminetetraacetic acid Mediatech, Inc.) for 5 min at 37 °C and passaged through a narrow-gauge pipette tip, resuspended in modified Krebs buffer (KRB) containing 3.3 mM glucose and 0.25% bovine serum albumin (BSA), and stained separately with probes for apoptosis (Annexin V PE, Invitrogen, Carlsbad, CA, USA; VAD-FMK FITC, Promega, Madison, WI, USA), and mitochondrial membrane polarity (MMP; JC-1, Invitrogen). Washed cells were analyzed on a 5-color FACS Calibur flow cytometer (Becton Dickenson, Franklin Lakes, NJ, USA). ToPro3 (Invitrogen) was added to samples immediately prior to analysis to identify necrotic cells. Data were analyzed using FlowJo software (Version 7.6.5) (Treestar, Inc., Ashland, OR, USA).
Glucose-Stimulated Insulin Secretion and ELISA
Islet aliquots containing 100 islet equivalents (IEQs) were cultured in triplicate in modified KRB containing 2.8 mM glucose for 60 min. Islet aliquots were then transferred to fresh KRB containing 2.8 mM glucose and cultured for another 60 min. Islets were then transferred to fresh KRB containing 28 mM glucose and cultured for another 60 min. All cultures were kept at 37 °C, 5% CO2. Media was collected from each consecutive culture condition, and secreted insulin content was measured by enzyme-linked immunosorbent assay (ELISA; Millipore, Billerica, MA, USA). Average values of secreted insulin content in the 28 mM glucose media were compared to those of the second 2.8 mM glucose media to establish the stimulation index (SI). SI values were normalized to those of untreated control islets for comparison across groups.
In Vivo Islet Functional Assessment
NOD SCID mice purchased from the Jackson Laboratory (Bar Harbor, MA, USA) were used as transplant recipients. Diabetes was induced by single intraperitoneal injection of 150 mg/kg streptozotocin (Sigma-Aldrich Corp and blood glucose was monitored daily via tail-nick blood drops on a portable glucometer (Bayer, Burr Ridge, IL, USA). Mice were considered hyperglycemic after 3 consecutive days with blood glucose readings >350 mg/dL. On the day of transplantation, donor islets were washed and resuspended in aliquots of Connaught Medical Research Laboratories (CMRL) transplant media (Mediatech, Inc.). Diabetic mice were anesthetized by isoflurane inhalation, and islets were implanted underneath the renal capsule as described elsewhere. 27 Blood glucose was monitored for 30 d after transplantation. Graft-bearing kidneys were then removed from recipients who achieved normoglycemia, and blood glucose was monitored for restoration to hyperglycemia within 72 h.
Statistics
GraphPad Prism version 5.0 (GraphPad, San Diego, CA, USA) and SPSS (IBM SPSS Statistics for Windows, Version 22.0. Armonk, NY, USA) were used for statistical analysis. Differences between various groups were compared by use of 1-way analysis of variance (ANOVA) with the Bonferroni multiple-comparisons test for post hoc analyses. For data sets with nonnormal distribution, Kruskal-Wallis test and Dunn’s posttest correction were applied. All data are shown as mean ± standard error of the mean. Time to cure after transplant analyses were performed with Kaplan–Meier survival analysis using the log-rank test followed by pairwise comparisons to determine statistical difference between the groups. The level of statistical significance was defined as a P < 0.05.
Results
Culture Supplementation with Exscien I-III Protects Purified Islets from Pro-Inflammatory Cytokines
In order to establish the therapeutic effectiveness of Exscien I-III in the peri-transplant pro-inflammatory environment, we evaluated islets purified from anesthetized rats after 48-h culture in the presence of exogenous cytokines IL-1β, TNF-α, IFN-γ. Cultured islets were analyzed by flow cytometry for apoptosis and MMP as previously described. 28 The addition of cytokines to culture resulted in a significant decrease in MMP% as measured by JC-1 staining compared to control islets (control: 82.6 ± 1.3% vs. cytokines: 71.1 ± 2.6%, P < 0.01). We did not observe a statistically significant difference in MMP% between cytokine alone versus cytokine + Exscien I-III-treated islets (Fig. 2A). We also documented a significant increase in the number of cells staining positive for markers of apoptosis (Annexin V and VAD-FMK) from dispersed cytokine-treated islets when compared to the untreated control group (P < 0.01). Treatment with Exscien I-III decreased the apoptotic rate by 50% in islets exposed to pro-inflammatory cytokines in culture (cytokines: 2.8 ± 0.5 vs. cytokines + Exscien I-III: 1.4 ± 0.3, P < 0.05; Fig. 2B). Cultured islets were also analyzed by fluorescent microscopy for viability and necrosis. Islets from cytokine-treated cultures showed a marked decrease in viability as compared to untreated control islets. The administration of Exscien I-III protected pancreatic isolated islets against the viability loss observed in the cytokine only group (control: 96.1 ± 1.0%; control + Exscien I-III: 92.2 ± 2.4%; cytokines: 61.5 ± 2.7%; cytokines + Exscien I-III: 81.8 ± 2.7%, P < 0.05; Fig. 3A and B). Next, cultured islets were analyzed by static incubation for glucose-stimulated insulin secretion to assess the impact of culture with pro-inflammatory cytokines and treatment with Exscien I-III on islet functional potency. Islets incubated in the presence of cytokines exhibited a significant reduction in SI in response to in vitro glucose challenge when compared to control, control + Exscien I-III, and cytokines + Exscien I-III (P < 0.05; Fig. 4A). Cultured islets were transplanted under the kidney capsule of diabetic immunodeficient mice at a dose of 500 IEQ/recipient. In mice transplanted with islets exposed to cytokines, we observed a 46.6% (n = 7/15) cure rate within a week posttransplant, while 33.3% (n = 5/15) remained diabetic for the duration of the experimental period. In contrast, 76.4% (n = 13/17) of the recipients of cytokines + Exscien I-III islets achieved normoglycemia within 7 d, and average daily blood glucose values closely matched those of recipients of control islets, demonstrating a significant protective effect through in vitro treatment with Exscien I-III against the impact of pro-inflammatory cytokines. Recipients of control + Exscien I-III islets achieved normoglycemia within 5 d of transplantation, and this group showed the lowest average glucose level and superior efficiency of cure (Fig. 4B and C).

Islet cell apoptosis and mitochondrial membrane polarity (MMP) induced by cytokines (tumor necrosis factor α [TNF-α] 1,000 U/mL, interferon γ [IFN-γ] 1,000 U/mL, and interleukin-1β [IL-1β] 50 U/mL) in culture are prevented by Exscien I-III (6.7 μg/mL) treatment. (A) Flow cytometric analysis of dispersed islet cell populations assessed for positive JC-1 staining (MMP) and (B) apoptosis rate (Annexin V, VAD-FMK) after 48-h culture. Results express mean ± standard error of the mean for each group: control (n = 6), control + Exscien I-III (n = 6), cytokines (n = 7), and cytokines + Exscien I-III (n = 5). Statistical analysis performed using 1-way analysis of variance (ANOVA) and the Bonferroni post hoc test. *P < 0.05, **P < 0.01, and ***P < 0.001.
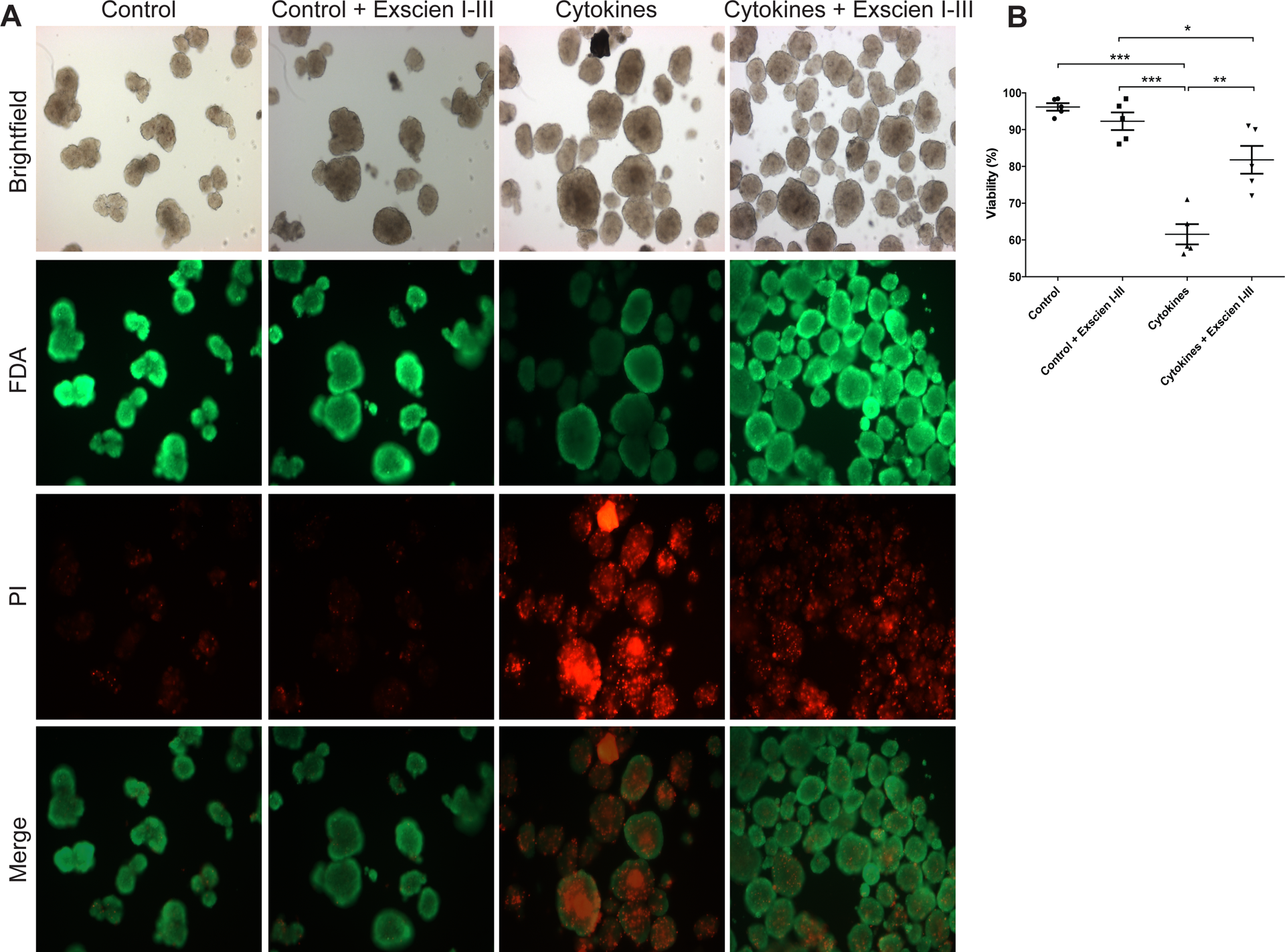
The addition of Exscien I-III in culture prevents cytokine-mediated islet necrosis. (A) Representative images (100×) of rat pancreatic islets stained in solution with fluorescein diacetate (FDA) as a fluorescent indicator of cellular viability (green) and propidium iodide (PI) as an indicator of necrosis (red). Islets in each group were cultured with cytokines (tumor necrosis factor α [TNF-α] 1,000 U/mL, interferon γ [IFN-γ] 1,000 U/mL, and interleukin-1β [IL-1β] 50 U/mL), Exscien I-III (6.7 μg/mL) or both for 48 h prior to assessment. (B) Quantification of viability of each islet preparation. Data are expressed as mean ± standard error of the mean for each group (n = islet preparations): control (n = 5), control ± Exscien I-III (n = 5), cytokines (n = 5), and cytokines + Exscien I-III (n = 5). Statistical analysis performed using 1-way analysis of variance (ANOVA) and the Bonferroni post hoc test. *P < 0.05, **P < 0.01, and ***P < 0.001.

Exscien I-III preserves the in vitro functional potency of cytokine-treated rat islets. (A) In vitro assessment of islet functional potency by glucose-stimulated insulin release (GSIR) after 48 h culture. Results are normalized to control group and expressed as percentage relative to control ± standard error of the mean. Statistical analysis performed using 1-way analysis of variance (ANOVA) and the Bonferroni post hoc test versus control. *P < 0.05, **P < 0.01, ***P < 0.001. (B) Streptozotocin-induced diabetic NOD SCID mice were transplanted with rat islets (500 islet equivalents [IEQs] each mouse, n = 5 per group) cultured for 48 h either alone or in combination with cytokines (tumor necrosis factor α [TNF-α] 1,000 U/mL, interferon γ [IFN-γ] 1,000 U/mL, and interleukin-1β [IL-1β] 50 U/mL), Exscien I-III (6.7 μg/mL) or both. Average daily blood glucose values for mice transplanted with islets from the respective treatment groups. (C) Cure rate (fraction of normoglycemic mice) in mice-receiving islets from each group. Data for blood glucose average expressed mean ± standard error of the mean, and statistical analysis was performed by 2-way ANOVA with *indicating a P < 0.05 versus all other groups. Cure rate data presented as fraction of animals achieving normoglycemia after transplantation and analyzed using Kaplan-Meier analysis (P < 0.001; log-rank test). All pairwise comparisons were significant at P < 0.05 except: control versus cytokines + Exscien I-III.
BD Model
To determine the therapeutic effect of Exscien I-III in a clinically relevant environment, we investigated the impact of treatment on islets using a model of inflammation after catastrophic brain injury. Hemodynamic parameters were monitored to ensure homogeneity in donor animal management and organ quality. Despite a trend toward lower mean arterial blood pressure and heart rate (HR) in the BD + Exscien I-III-treated animals, statistical analysis did not reveal significant differences when compared to animals in the BD + control group (P > 0.05; Fig. 5A to C).
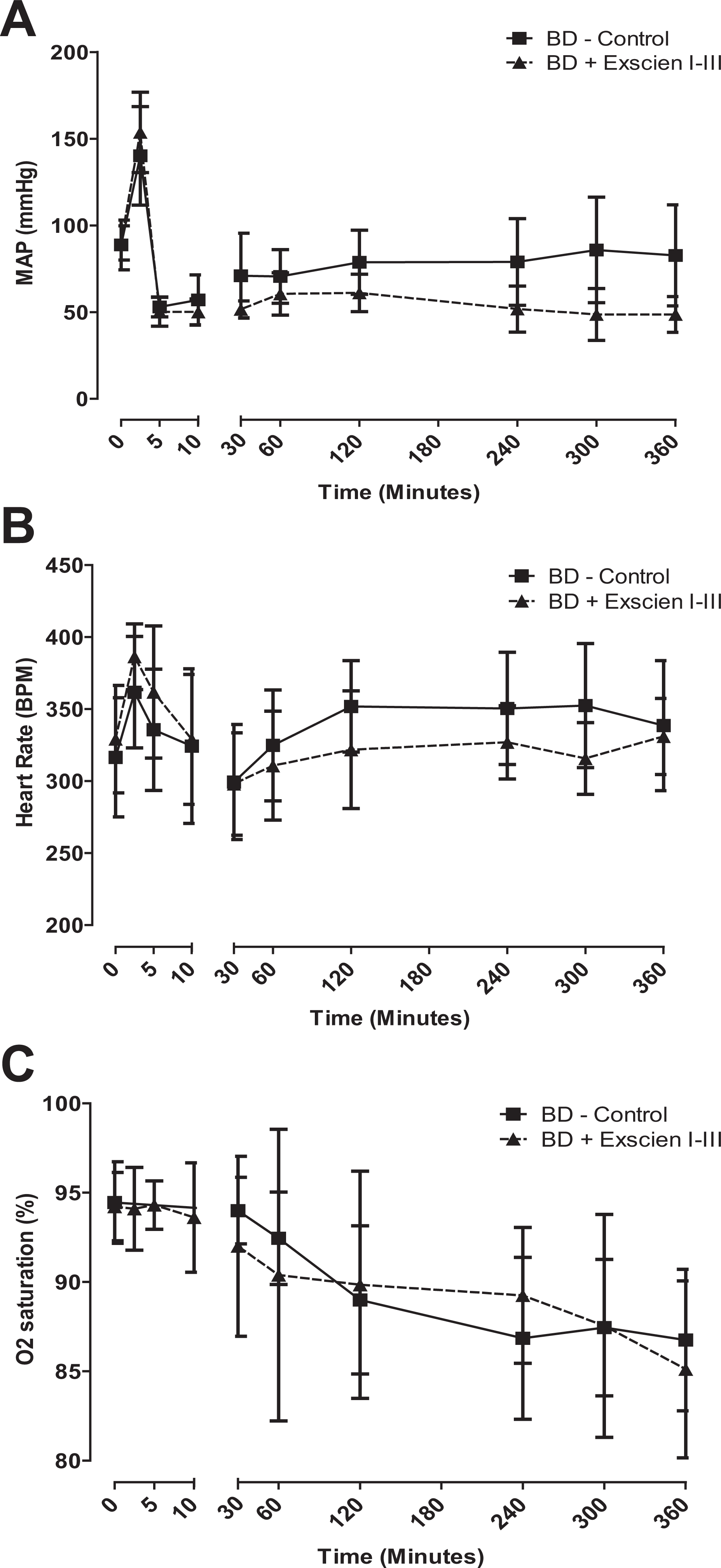
Hemodynamic monitoring of brain-dead rats. (A) Mean arterial pressure, (B) heart rate, and (C) oxygen saturation were monitored for a 6-h period following brain death induction in control (n = 34) and Exscien I-III (n = 22)-treated animals. Data are expressed as mean ± standard error of the mean. Statistical analysis did not reveal significant differences between the groups.
Donor Exscien I-III Treatment Prevents BD-Induced Impairment in Glucose-Stimulated Insulin Secretion
BD has also been shown to impact the functional capacity of pancreatic islets to respond to glucose stimulation. 6,29 Islets from BD and non-BD donors were evaluated for in vitro glucose-stimulated insulin secretion. Insulin secretion from the BD + control donors was significantly impaired relative to non-BD control levels, whereas BD + Exscien I-III donor islets were not impaired but rather exhibited improved insulin secretion relative to non-BD control, although this comparison did not achieve statistical significance (Fig. 6A).

Exscien I-III treatment restores functional potency of islets transplanted from brain-dead rats. (A) In vivo assessment of glucose-stimulated insulin secretion in islets isolated from either sham or brain-dead (BD) donors receiving vehicle or Exscien I-III treatment and cultured for 24 h. Results are normalized to non-BD control group and expressed as percentage ± standard error of the mean. Statistical analysis was performed using 1-way analysis of variance (ANOVA) and the Bonferroni post hoc test. (B) Blood glucose monitoring of streptozotocin (STZ)-induced diabetic NOD SCID mice transplanted with 250 islet equivalents (IEQs) donor rat islets under the kidney capsule. Blood glucose monitoring results are expressed as mean ± standard error of the mean, and statistical analysis was performed by 2-way analysis of variance (ANOVA) with *indicating a P < 0.05 versus all other groups. (C) Cure rate of NOD SCID mice after xenogeneic islet transplantation. Mice in the non-BD control (n = 10), non-BD + Exscien I-III (n = 8), BD-control (n = 9), and BD + Exscien I-III (n = 8) groups were followed for 30 d after implantation and monitored daily for return to normoglycemia (defined as random nonfasting blood glucose <250 mg/dL for 3 consecutive days). Cure rate data presented as fraction of animals achieving normoglycemia after transplantation and analyzed using Kaplan-Meier analysis (P < 0.001; log-rank test). All pairwise comparisons were significant at P < 0.05 except non-BD–control versus BD + Exscien I-III, non-BD−control versus non-BD + Exscien I-III, and BD + Exscien I-III versus Non-BD + Exscien I-III.
Donor Exscien I-III Treatment Prevents BD Induced Impairment of Allograft Functional Potency
Islets from BD and non-BD donors were assessed for in vivo functional potency by transplanting 250 IEQ under the kidney capsule of diabetic immunodeficient NOD SCID mice. In this experiment, 100% (10/10) recipients of islets from non-BD vehicle-treated donors achieved normoglycemia but with only 60% (6/10) achieved sustained normoglycemia by posttransplant day 7. In contrast, only 33% (3/9) of recipients transplanted with islets from vehicle-treated BD donors were able to achieve normoglycemia during the entire 30 d of the experiment and none within 7 d of the procedure. All recipients transplanted with islets from non-BD + Exscien I-III donors (8/8) and BD + Exscien I-III donors (8/8) achieved normoglycemia within 5 d after transplantation, exhibiting both superior cure rate and sustained functional potency (P < 0.001; Fig. 6B and C).
Discussion
In this study, we investigated the use of Exscien I-III, a mitochondrially targeted DNA glycosylase/AP lyase as a supplement for islets in culture or as a treatment strategy for brain-dead donors in the peri-transplant environment. Our data suggest that Exscien I-III has a robust protective effect on islet function and viability, and these findings are applicable to both in vitro and in vivo models of inflammatory injury. Our group and others have previously shown that the supplementation of pro-inflammatory cytokines to islets in culture induces apoptosis and decreases viability. 30 –34 The mechanisms by which pro-inflammatory cytokines induce β cell injury encompass both NO-dependent and independent pathways; in pancreatic islets, sustained exposure to IL-1β has been shown to induce NO production by direct upregulation of the inducible nitric oxide synthase (iNOS). 35 NO excess results in the irreversible inhibition of β-cell secretory function and can be genotoxic and proapoptotic by targeting mtDNA that encodes for mitochondrial proteins of the electron transport chain necessary for electron transport and mitochondrial ATP synthesis. 30,36 Our experimental results indicate that dispersed cells from islets cultured with cytokines for a 48-h period have a substantial increase in apoptosis (Annexin V and VAD-FMK staining), which is prevented by Exscien I-III treatment. This is in agreement with the observation that mtDNA is a target for NO-induced toxicity driving apoptosis in β-cells after exposure to free fatty acids. 37 In 2006, Rachek et al. conducted 2 similar studies in which they demonstrated that stable overexpression of 8-oxoguanine DNA glycosylase (Ogg1) in mitochondria of HeLa and the INS1 rat insulinoma cell line prevented apoptosis prevented apoptosis by limiting NO-mediated mtDNA damage. 21,38 The decrease in apoptosis observed in our experiments was associated with a parallel increase in functional potency manifested by a superior glucose-stimulated insulin secretion capacity in vitro, along with a reduced time to cure after transplantation and markedly improved glycemic control in immunodeficient mice receiving Exscien I-III-treated islets. These findings corroborate previous reports in which IL-1β antagonism blockade in vivo enhanced islet engraftment and improved the impairment in glucose tolerance observed in recipients of grafts exposed to the inflammatory environment. 39,40 A growing body of evidence suggests a role of the mitochondrial pathway in inflammatory cytokine–mediated pancreatic β-cell death. In rat RINm5F cells, a combination of IL-1β, IFN-γ, and TNF-α has been shown to induce mitochondrial membrane depolarization, and rat islets exposed to IL-1β were found to be highly susceptible to apoptosis mitochondrial cytochrome c release and caspase-9 activity. 41
In the clinical setting, a recent phase 3 clinical trial demonstrated that transplantation of human islets remains an effective treatment for T1DM and resulted in restoration of hypoglycemia awareness, elimination of severe hypoglycemic events, and normalization of glycemic control in approximately 88% of participants. 42 However, the widespread success of islet transplantation has been limited by the need for multiple donor pancreata and repeated islet infusions to achieve insulin independence in recipients. 43 Despite progressive refinement of the technique, islet transplantation is still limited by the supply of high-quality transplantable organs, and institutions and clinicians are turning to the use of lower quality expanded criteria donor (ECD) organs in an attempt to meet an ever-growing demand to increase the donor pool. 44 Cadaveric organs present a significant challenge to successful long-term graft function when compared to organs recovered form living donors. 45 As we gain better understanding of the mechanisms underlying organ dysfunction in deceased donors, we are discovering new targets for therapies to improve organ quality, and significant attention has been devoted to treatments that may protect tissue injury from oxidative stress. 46 –48 In islet transplantation, the donor pancreas is particularly susceptible to inflammatory and subsequent oxidative damage encountered during donation, cold storage, the isolation process, and engraftment. 9,49 Previous efforts have focused on reducing hypoxia and ischemia after organ recovery through advanced formulations of organ procurement solution and to reduce reperfusion injury through antioxidant and immunosuppressive therapies. 11,50,51 Moreover, there is increasing evidence of the negative impact on organ quality associated with the stable use of brain-dead pancreas donors and a strong correlation with suboptimal islet yields and potency.2, 8,52 In 2003, Contreras et al. provided experimental evidence suggesting lower postisolation islet yields and an increase in the apoptotic rate of pancreatic islets using a rodent model of BD pancreas donation. 6 Despite advances in management with vasoactive and hormonal therapies, currently there are no clinically effective strategies targeting inflammation and oxidative stress in brain-dead donors. 53 For this part of the study, we administered an intravenous bolus of Exscien I-III targeted to the mitochondria of pancreatic islets in pancreas donors and the treatment of BD rats prior to organ procurement resulted in a significant improvement in viability and functional potency after islets were transplanted into diabetic hosts. The mechanism by which Exscien I-III is protective in this model is not completely clear. Recently, multiple reports have suggested that oxidative mtDNA damage is important for reasons that extend beyond the cell harboring the damage. In this context, recent findings indicate that oxidative damage to mtDNA leads to its degradation or fracture into short sequences that are biologically active. There appears to be at least 3 destinations of the damage-related fragments. First, cytoplasmic mtDNA may autonomously activate endosome Toll-like receptor 9 (TLR-9) to provoke local inflammation and cellular dysfunction. 54,55 Second, cytoplasmic fragments may serve as coactivators in the nucleotide-binding oligomerization domain–like receptor family, pyrin domain containing 3 activation of caspase-1 and ensuing pro-inflammatory cytokine production. 56 Finally, there is evidence suggesting that mtDNA may have pro-inflammatory properties by acting as a damage-associated molecular pattern. 57 In various traumatic conditions including shock and severe traumatic brain injury, release of mtDNA has been shown to directly activate neutrophils by interaction with TLR9 and recent clinical studies in the trauma literature provide evidence that circulating mtDNA levels are associated with clinical outcome in severely injured human subjects. 58 –60 There is evidence that Exscien I-III may play a role in protection from ischemic injury by preventing the breakdown and release of pro-inflammatory mtDNA as recently shown in a model of ischemic injury to the myocardium.26 We believe that the treatment of the donor animal with the Exscien I-III reduced the mtDNA damage caused by brain death both in the islets and other tissues and caused a decrease in BD-induced inflammation. This treatment rendered the islets more healthy for transplantation. Accordingly, in animal models of BD donor, management with anti-inflammatory and β-cell protective agents has been shown to improve postisolation islet quality and function. 61 We have previously demonstrated that targeted blockade of the inflammatory response in a large animal model of BD using an IL-1 receptor antagonist (Anakinra) was associated with decreased pancreas and islet inflammation and significantly improved graft function in mice receiving islets from treated BD donors. 29 Moreover, Carlessi and colleagues used a rodent model of BD to show that islets from donor rats treated with Exendin-4, a glucagon-like peptide-1 analog with anti-inflammatory and antioxidant properties on β-cells, had a significant reduction in pancreatic IL-1β messenger RNA (mRNA) expression and stress-related genes. 49 Although they did not test the impact of Exendin-4 management on graft quality and survival in vivo, their data strongly suggest that early targeting of inflammation and oxidative stress in the BD donor exerts a beneficial effect through attenuation of oxidative damage, which may in turn prevent islet endoplasmic reticulum stress and β-cell necrosis. Recognizing the pivotal role of ROS in islet dysfunction, there has been a push to minimize oxidant levels in islets in culture through the addition of synthetic antioxidants, mimics of endogenous antioxidants (e.g., MnSOD),or through transfection of islet cells with MnSOD genes. 62,63 Most of these efforts have met with limited success due to inherent difficulty in achieving stable concentration levels in the mitochondria to protect cells from mtDNA induced by ROS and oxidative injury.
One of the main advantages of this treatment is that it is minimally invasive, and delivery of the protein to target organs is rapidly detectable and has lasting effectiveness. No cytotoxicity was detected as a result of the therapy, and in some instances donor management with Exscien I-III appeared to improve islet potency and transplant outcomes over that of untreated living-donor controls. The use of this protein presents potential clinical advantages for transplant recipients such as a reduction in the islet mass required to reverse diabetes, minimized dependence on exogenous insulin injections, and an overall improvement in graft function and graft survival. This enhanced functional capacity of islet grafts may in turn optimize the quality of organs transplanted from marginal donors and thereby broaden the donor pool.
There are a number of limitations to the study we conducted. Rodents are not the most reliable analogues for human patients nor does our model fully represent the myriad aspects of clinical BD and the transplant environment. Although we confirmed the presence of the protein in donor pancreata following intravenous administration, we did not conduct studies to identify the protein in our islet preparations following recovery and rely on a large body of literature investigating the kinetics of delivery, localization, elimination, and known mechanisms of action of Exscien I-III in vivo.
In summary, our findings indicate that the delivery of a bacterially derived mitochondrially targeted DNA repair enzyme results in improved islet viability postisolation, limits cytokine-induced apoptosis, and enhances islet functional potency, thus reducing the total islet-mass required to reverse diabetes in diabetic recipients. Altogether, this therapy holds promise for the treatment of conditions associated with inflammatory and oxidative injury, and further investigation is warranted to determine its effectiveness in the clinical transplant setting.
Footnotes
Authors’ Contribution
Juan S. Danobeitia and Peter J. Chlebeck contributed equally to this work.
Acknowledgments
The skilled technical and editorial assistance provided by L. Zitur and I. Segel is gratefully acknowledged. We would also like to thank Exscien corporation for kindly providing the investigational molecule used in this study. Finally, we thank Dr Brooks Rademacher for assistance with statistical analysis.
Ethical Approval
All experiments were performed in accordance with the guidelines established and approved by the University of Wisconsin-Madison Institutional Animal Care and Use committee.
Statement of Human and Animal Rights
Donor rats were humanely anesthetized as described in the Materials and Methods section above.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: This research was supported by funds from the National Institutes of Health Grants ES03456 (Glenn Wilson), T32 training T32AI125231 (J. Danobeitia), American Society of Transplant Surgeons Scientist Scholarship (J. Danobeitia), and internal funds of the Department of Surgery at the University of Wisconsin–Madison.