
Abstract
The INK4-ARF locus includes the CDKN2B and CDKN2A genes and is functionally relevant in the regulation of both cell proliferation and senescence. Studies have reported modifications of DNA methylation in this locus by exposure to environmental contaminants including pesticides; however, until now, specific methylation profiles have not been reported in genetically conserved populations exposed to occupational pesticides. The aim of this study was to determine the methylation profiles of the CDKN2B and CDKN2A genes in a genetically conserved population exposed to pesticides. A cross-sectional and analytical study was carried out in 190 Huichol indigenous persons. Information related to pesticide exposure, diet and other variables were obtained through the use of a structured questionnaire. Blood and urine samples were collected for methylation test and dialkylphosphates (DAP) determination, respectively. DNA methylation was measured by the pyrosequencing of bisulfite-treated DNA and DAP concentrations by gas chromatography-tandem mass spectrometry (GC/MS). The most frequent metabolite in the population was dimethylthiophosphate. The farmer group presented a higher methylation percentage of CDKN2B than the non-farmer group, but no differences in CDKN2A were observed between groups. A positive correlation between methylation of CpG site 3 of CDKN2B and time working in the field was observed in the farmer group. An association between methylation percentage of CDKN2B and age was also observed in the non-farmer group. These results suggest that pesticide exposure and exposure time in Huichol indigenous individuals could modify the methylation pattern of the CDKN2B gene.
Highlights
1. Huichol farmworkers showed higher methylation of CDKN2B than non-farmworkers. 2. A correlation between the methylation of CpG site 3 of the CDKN2B gene and work time in the field was observed. 3. Occupational pesticide exposure and exposure time may modify the methylation of CDKN2B gene.
Introduction
Pesticides are widely used around the world in the agricultural sector to improve crop yields.1,2 Organophosphate pesticides (OPs) represent a class of extensively used pesticides with high toxicity and widespread use. 3 After absorption, OPs are metabolized into the more reactive oxon forms, which may bind to cholinesterases with their consequent inhibition, or be hydrolyzed into non-specific dialkylphosphate (DAP) metabolites, these are widely used as biological indicators of these compounds.4,5
OPs can alter cell components such as lipids and proteins, as well as DNA through genotoxic damage and epigenetic modifications.6–11 Epigenetic changes refer to chemical modifications of DNA or of the structural and regulatory proteins bound to it, which do not involve changes in nucleotide sequence and that are potentially transmissible to daughter cells. Examples of epigenetic modifications include DNA methylation, histone modifications, and microRNA expression.12,13 In the human genome, about 28 million CpG sites are susceptible to methylation changes, approximately 10% of which are in the promoter regions of genes. Such changes in methylation might alter the binding of transcription factors to DNA.14–16
Outside the promoter region, DNA methylation modification at the untranslated regions (UTR) is important in the regulation of gene expression.17,18 UTR contain various regulatory elements and are involved in the post-transcriptional regulation of gene expression, modulation of the transport of messenger RNA out of the nucleus, and control of translation initiation.18,19 Loss of epigenetic regulation manifests as global hypermethylation of DNA related to genomic instability, chromosomal aberrations, reactivation of transposable elements, loss of imprinting, and gene-specific hypermethylation, mainly of tumor suppressor genes.12,20,21
The inhibitor of cyclin-dependent kinase 4a/alternative reading frame (INK4-ARF) locus encodes two cyclin-dependent kinase inhibitors, CDKN2B and CDKN2A, and a regulator of the p53 pathway known as ARF. This locus is functionally important in regulating cell proliferation and senescence.22,23 The CDKN2B and CDKN2A proteins are cyclin-dependent kinase inhibitors (CDK) of the cyclin/Cdk4 and cyclin/Cdk6 complexes. These complexes in turn act to phosphorylate and inactivate retinoblastoma protein (pRb), a protein which is critical in the regulation of cell cycle progression from the G1 to the S phase. By inhibiting the cyclin/cdk4 and cyclin/cdk6 complexes, CDKN2B and CDKN2A activity thus results in the activation of pRb and the downstream events which drive cell cycle progression.24–26 Several investigations have examined the inactivation of CDKN2B and CDKN2A by gene deletion, mutation, and hypermethylation in the CpG islands of the promoter region.26–28
Some studies have evaluated the influence of race, ethnicity, and genetic ancestry on changes in DNA methylation patterns.29–31 However, to date there have been limited reports regarding the DNA methylation profiles of genetically conserved indigenous populations. The Huichol population is an indigenous group of Mexico whose members migrate to different regions of the country and who are mainly employed in jobs related to agriculture and the application of pesticides; further, they reported no use of personal protective equipment and have limited access to health care, among other factors that render them vulnerable to the toxic effects of these compounds.32,33 Therefore, in this study, the methylation profiles of the CDKN2A and CDKN2B genes was evaluated in a genetically conserved Huichol population, as was the relationship of these methylation profiles to pesticide exposure.
Materials and methods
A cross-sectional and analytical study was carried out in Nayarit, Mexico, in 190 conserved Huichol indigenous older than 18 years of age. All selected participants were required speak the indigenous language, share cultural traditions, and have blood type O (ABO system) and the Rh+ antigen, this a trait of genetically conserved indigenous communities and considered a genetic fingerprint in this population. All participate voluntarily in the study and all signed a written informed consent document. This study was conducted in accordance with the international guidelines of the Declaration of Helsinki 34 and was approved by the Bioethics Commission of the Nayarit State, Mexico (registry number CEBN/01/2016). The aims of the study were explained to the participants, who responded to a structured questionnaire that collected general information related to the population, including sex, age, body mass index (BMI), alcohol consumption, smoke exposure, medical history, diet, and pesticide exposure. The section on pesticide exposure included specific information on work activity, time spent working in the field, use of pesticides, period of pesticide exposure, and others.
In addition, the software program Evaluation of Nutritional Habits and Nutrient Consumption System (SNUT) 35 from the National Institute of Public Health and National Institute of Cardiology Ignacio Chávez, both of these in Mexico, was used for the determination of macro- and micro-nutrient intake according to a food frequency questionnaire.
Blood samples were collected in tubes with EDTA disodium salt for methylation analysis and morning urine samples were collected for DAP determination. Blood and urine samples were stored at −20°C until analysis.
Quantification of dialkylphosphate metabolites
Extraction of urinary analytes was performed according to the method of Valcke et al. 36 and that of Ramírez-Jiménez et al. 37 Urinary concentrations of diethylphosphate (DEP), diethylthiophosphate (DETP), diethyldithiophosphate (DEDTP), dimethylphosphate (DMP), dimethylthiophosphate (DMTP), and dimethyldithiophosphate (DMDTP) were determined using a gas chromatograph (Agilent 7820A GC; Agilent Technologies) tandem mass spectrometer (5975 Series MSD; Agilent Technologies), through selective ion monitoring. The internal standard was deuterated diethylphosphate (DETP-d10) and was determined in each set of samples. The retention time, lineal range and the limits of detection, and quantification of each metabolite can be seen in Table S1.
DAP concentrations were corrected by urinary specific gravity according to the method of Sokoloff et al. 38 and were expressed in ng/mL. DAP concentrations were analyzed as the sums of metabolites for the following groups: ∑DEP (DEP+DETP+DEDTP); ∑DMP (DMP+DMTP+DMDTP), and ∑DAP (DMP+DMTP+DMDTP+DEP+DETP+DEDTP), based on a previous study of a population of urban sprayers. 39
DNA methylation status of CDKN2B and CDKN2A genes
DNA extraction and bisulfite modification
Genomic DNA was isolated from the blood using a PureLink® Genomic DNA Mini Kit (Invitrogen) according to the manufacturer´s instructions. The purity and concentration of the extracted genomic DNA were determined by spectrophotometry (NanoDrop 2000c; Thermo Scientific). From 500 ng DNA, bisulfite conversion was carried out with the EZ DNA Methylation-Lighting™ kit (Zymo Research) according to the manufacturer’s protocol. Bisulfite-treated DNA samples were stored at −20°C until methylation analysis.
Polymerase chain reaction amplification and pyrosequencing
Primers and analyzed sequences of CDKN2B and CDKN2A genes.

F: forward primer; R: reverse primer; S: sequencing primer; Btn: biotinylated; PQS: pyrosequencing; bp: base pairs, PCR: polymerase chain reaction.
For pyrosequencing, 0.3 μM of sequencing primer was used with the PyroMark Q24 system (software version 2.0.7) in order to quantify the percentage of methylation on CpG sites according to the manufacture’s protocol and the sequencing plate was processed in the PyroMark Q24 Instrument. The results were calculated as the percentage of methylated cytosines (%5 mC). As a control for DNA methylation, a sample of 50% methylated DNA (QIAGEN; Qiagen) was included in each sequencing plate evaluated, and an external control for bisulfite conversion was added to each sample during the pyrosequencing analysis.
For the CDKN2B gene, the analyzed sequence was located in the 5′UTR region, and five CpG sites (+21 to +73 bp) were evaluated. For the CDKN2A gene, the analyzed sequence was located in the proximal promoter region and six CpG sites (−205 to −146 bp) were evaluated, according to Herrera-Moreno et al. 10 The NCBI reference sequences were NG_023297 and NG_007485 for CDKN2B and CDKN2A genes, respectively.
Statistical analysis
Data distribution was determined by Skewness Kurtosis test and descriptive statistical analyses were carried out. The student t-test and the Mann–Whitney U test were used to compare the farmer and non-farmer groups for data with parametric and non-parametric distributions, respectively. Methylation levels of the CpG sites measured were averaged to yield a mean %5 mC. The correlations between %5 mC of CDKN2B and CDKN2A genes with time working in the field and age were evaluated by Spearman rank correlation coefficients. A regression analysis of the %5 mC of CDKN2B and CDKN2A genes in the participants was carried out with covariates such as age, sex, and time working in the field, among others; the p-value was adjusted by the types of white blood cells (neutrophils, lymphocytes, eosinophils, basophils, and monocytes). All analyses were performed using version 14.0 of STATA software (Stata Corporation, College Station, TX, USA) and p <0.05 was considered statistically significant.
Results
General characteristics of the study participants
General characteristics of the study population.

ap-values were obtained by chi-square test.
bMann–Whitney U test and.
cBody Mass Index (BMI) according to World Health Organization (WHO) parameters: low weight (≤18.5 kg/m2); normal weight (>18.5 kg/m2 and ≤24.9 kg/m2); overweight (>24.9 kg/m2 and ≤29.9 kg/m2), and obesity (≥30 kg/m2). GM: geometric mean; M: arithmetic mean; SD: standard deviation; 95% CI: 95% confidence interval.
dStudent’s t-test.
Pesticides used by the study population.

aWorld Health Organization (WHO, 2020) classification: class Ia, extremely hazardous; class Ib, highly hazardous; class II, moderately hazardous; class III, slightly hazardous; class U, unlikely to present acute hazard.
Dialkylphosphate metabolites in the study population
The metabolite found most frequently in our sample of the study population was DMTP with a GM of 27.9 ng/mL (95% CI: 24.9–31.2 ng/mL) for both farmer and non-farmer groups, while DMP was found only in the farmers group with a GM of 10.6 ng/mL (95% CI: 5.8–19.5 ng/mL). Likewise, the ∑DMP was higher than the ∑DEP concentration in the study population; however, farmers and non-farmers had similar ∑DAP concentrations (Figure 1). Urinary concentration of dialkylphosphate in the reference and farmer groups of the study population. ∑DMP (DMP+DMTP+DMDTP); ∑DEP (DEP+DETP+DEDTP); ∑DAP (DMP+DMTP+DMDTP+DEP+DETP+DEDTP). The p-value was obtained by Mann–Whitney U test.
DNA methylation of CDKN2B and CDKN2A genes
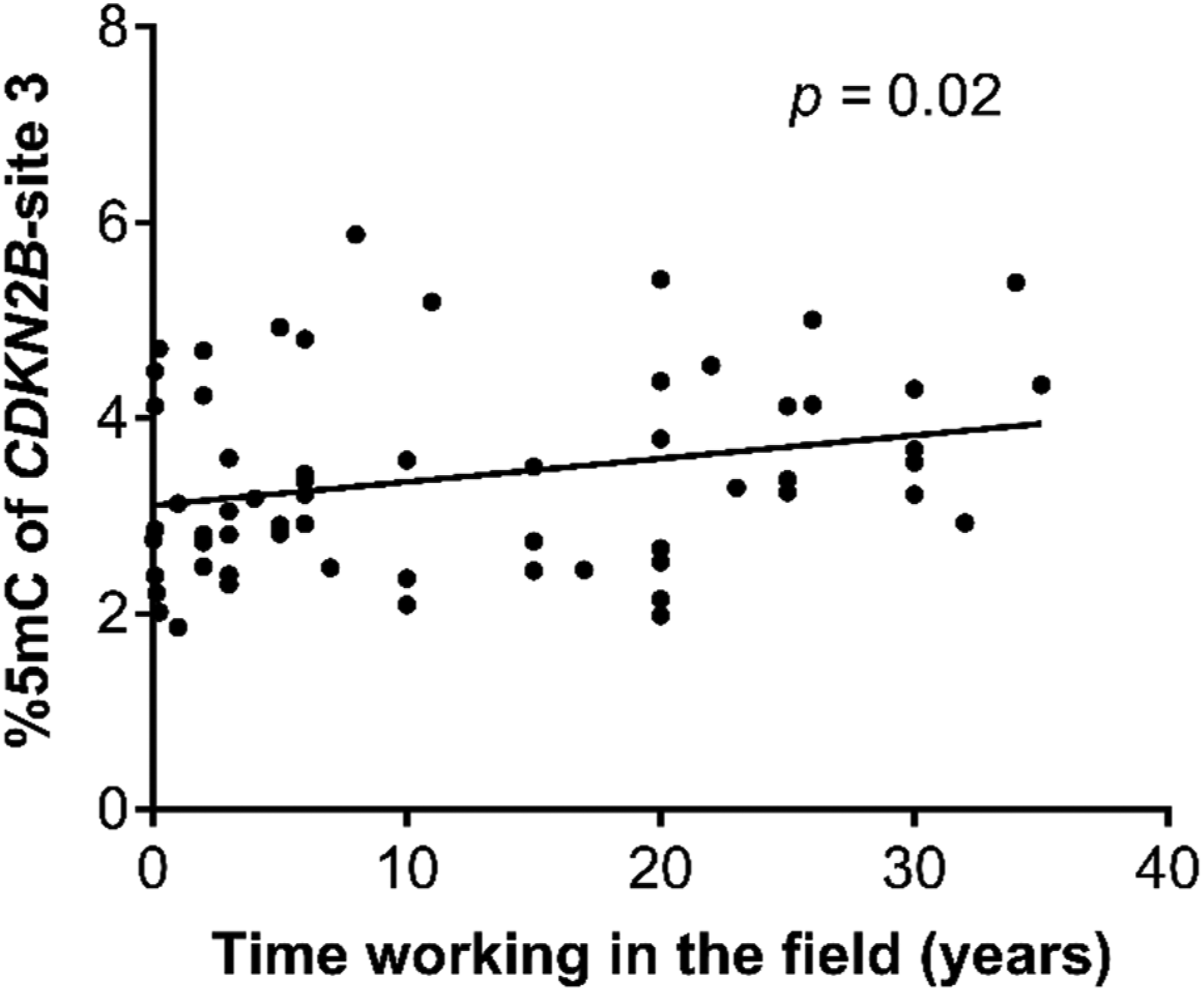
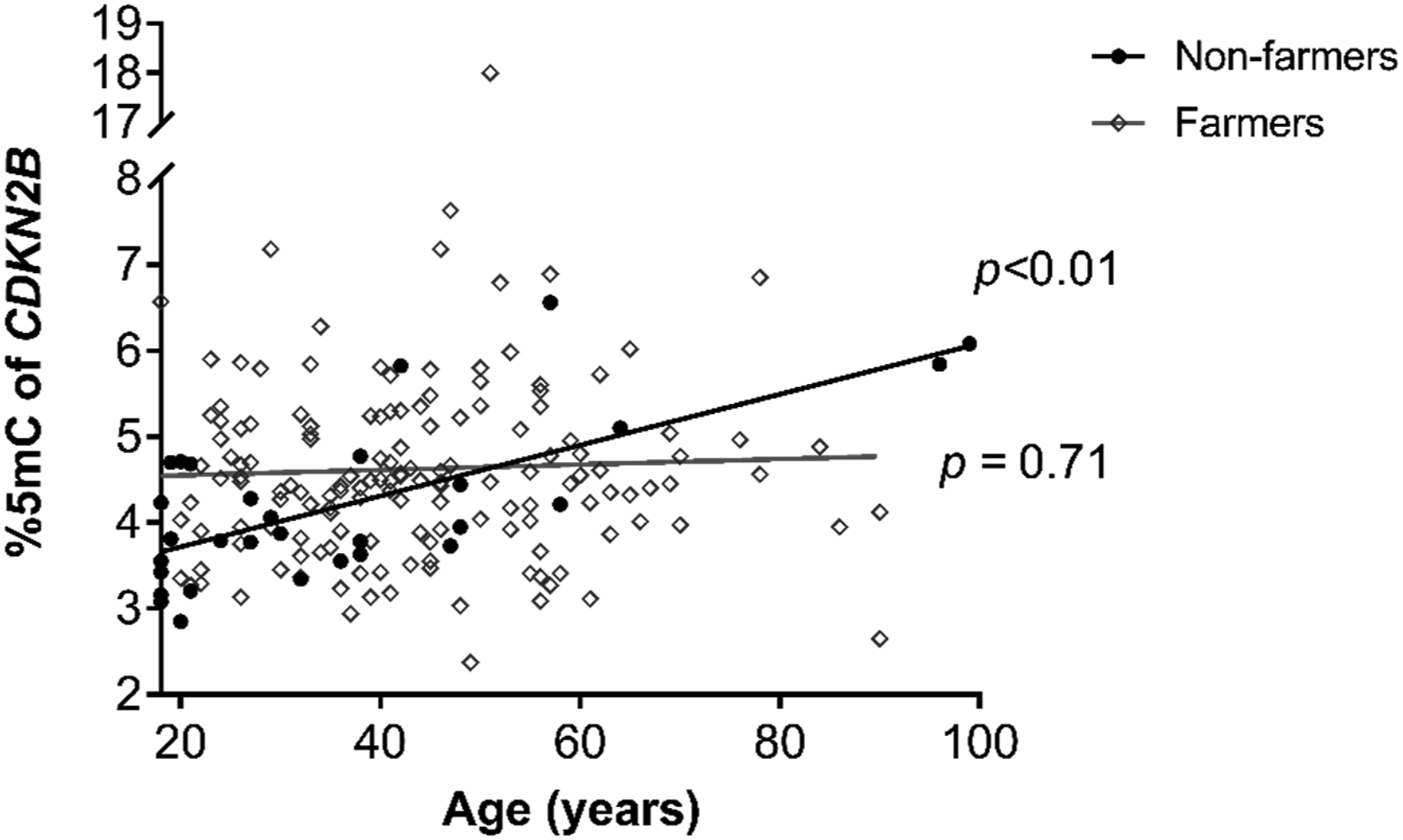
With respect to gene methylation patterns, the average percentage of methylation in the CDKN2B gene was 4.42 %5mC (95% CI: 4.27–4.57 %5mC), with the farmer group presenting a higher %5mC of the CDKN2B gene (4.48 %5mC, 95% CI: 4.31–4.65 %5mC) than the non-farmer group (4.11 %5mC, 95% CI: 3.80–4.44 %5mC) (Figure 2(a)). Analysis by CpG site found that the percentage of %5mC in CpG site 1 of CDKN2B was higher in the farmer group than the non-farmer group (Table 4). Meanwhile, no differences were observed in total methylation of the CDKN2A gene between the farmer (8.33 %5mC; 95% CI: 7.83–8.86 %5mC) and non-farmer groups (8.70 %5mC; 95% CI: 7.99–9.47 %5mC) (Figure 2(b)), nor were differences between these groups seen for site-specific methylation of the CDKN2A gene (Table 4), since there was a positive correlation between the methylation percentage of CpG site 3 of the CDKN2B gene and time working in the field in individuals between 18 and 55 years of age (Figure 3) in the farmer group. Also, an association between %5mC of the CDKN2B gene and age was observed, but only in the non-farmer group (Figure 4). A negative correlation was observed between %5mC of CDKN2B and %5mC of CDKN2A (Figure 5). No significant differences were observed in the methylation percentage of CDKN2B or CDKN2A genes in relation to general population characteristics (sex, BMI, and age), alcohol consumption, smoking status, and macro- and micro-nutrient intakes for the study population (Tables 2S–4S). Likewise, we did not find significant associations between urinary DAP metabolites with the methylation percentage of CDKN2B and CDKN2A genes. Percentage gene methylation in an indigenous farmer and non-farmer groups. (a) Methylation of CDKN2B and (b) CDKN2A genes. Bars represent the geometric mean and 95% confidence interval. Statistical analysis was performed with Mann–Whitney U test. Percentage methylation in CpG sites of the CDKN2B and CDKN2A genes. GM: Geometric mean; 95% CI: 95% confidence interval. The p-values were obtained by Mann–Whitney U test. p < 0.05 was considered statistically significant. Correlation analysis between time working in the farm and percentage methylation of the CDKN2B gene at CpG site 3. The p-value represents the significance of Spearman correlation test. Association between age and percentage methylation of the CDKN2B gene in farmer and non-farmer groups. The p-value represents the significance of association analysis by a generalized linear regression model. Correlation between averages of CpG sites analyzed from the CDKN2B and CDKN2A genes. The p-value represents the significance of Spearman correlation test.


Discussion
Several studies have demonstrated that alterations in the levels of DNA methylation can contribute to the development of different human diseases.44–46 Some of these DNA methylation changes could be influenced by environmental factors such as pesticide exposure.47–49
Previously, studies in vitro and in vivo have noted a decrease of global DNA methylation in response to pesticide exposure, as well as an increase in mutations, chromosomal instability, and cancer risk.8,50–52 Consistent with this, Kostka et al. 53 found an increase of DNA methylation in the promoter regions of the p53 and p16 genes in rats treated with DDT. Additionally, DNA methylation changes have been observed in the promoter regions of the CDKN2B and CDKN2A genes in populations exposed to different pollutants such as arsenic, benzene, nickel, tobacco, and coke oven emissions in comparison with non-exposed groups.27,54–56 Nevertheless, information related to epigenetic modifications and pesticide exposure in genetically conserved populations is limited.
Our data did not show differences in DAP urinary metabolites between the study groups except for the DETP (Table 5S), probably because the whole study population lived near agricultural or farm areas, which left them vulnerable to potential pesticide exposure from the air and other sources. Accordingly, studies have previously reported the influence on the internal doses of pesticides by non-direct exposure in individuals residing in areas near agricultural fields.57–59 Additionally, the participants manifested the use of pesticides at home and through the consumption of crops.
Regarding the DNA methylation profile, our results showed an increase in %5mC in the 5′UTR region of the CDKN2B gene in indigenous farmers compared to the non-farmers; conversely, there were not significant differences between study groups for CDKN2A gene methylation. Contrary to our results, Herrera-Moreno et al. 10 reported a decrease in %5mC in the CDKN2B gene for groups occupationally exposed to pesticides (urban sprayers) in comparison to a reference group. In contrast with our results, Herrera-Moreno et al. 10 found higher methylation in the CDKN2A gene in urban sprayers with moderate exposure to pesticides with respect to a reference group. Based on these differences in the effect of xenobiotics on gene-specific DNA methylation profiles, studies have reported that these variations may occur depending on the compound or mixture of compounds to which the population is exposed; time and intensity of exposure, as well as other factors such as race and ethnicity.30,60,61
The increased methylation percentage in the 5′UTR region of the CDKN2B gene that we have observed in our population of farmers could be involved in the development of several types of cancer, as has been reported in previous studies.26,28,62 However, further investigations are necessary to provide evidence of the relationship between pesticide exposure and an increase of specific gene methylation in this type of population. Previous studies that support a relationship between alterations in DNA methylation and pesticide exposure in humans include a study by van der Plaat et al., 60 the authors found associations between occupational exposures to pesticides and changes in 31 CpG sites related to 29 genes. Likewise, Howard et al. 63 observed alterations in the methylation of 36 CpG sites in or near 72 genes in farmers exposed to pesticides compared with non-farmers; those genes could be affected by different metabolic pathways related to immune response, growth hormone signaling, DNA damage response, among others.
In this study, we observed a positive correlation in the farmer group between %5mC in CpG site 3 of CDKN2B and time spent working on at farm. We also found a correlation in the non-farmer group between age and %5mC in the CDKN2B gene overall. Several studies related to DNA methylation have reported age-associated changes; about one hundred CpG sites have been reported to show age-associated methylation modifications in different human tissues including whole blood. 64
Analysis of specific gene regions reveal that methylation in promoter regions associated with CpG islands tends to increase with age due to the influence of external factors and that, in some cases, these epigenetic markers can be heritable through cell division. The accumulation of these markers could contribute to epigenetic changes with age. Changes in the epigenetic machinery due to exposure to environmental factors have been demonstrated to increase epigenetic variability, and possibly contribute to the development of age-related illnesses such as neurological diseases.65–69
Studies have evidenced the influence of environmental exposure and genetic ancestry on DNA methylation in multiple specific locations of the human genome. Galanter et al. 30 studied DNA methylation patterns in 573 individuals from some Latino ethnic sub-groups and found that genetic ancestry and ethnicity were associated with DNA methylation levels at 194 and 916 CpG sites, respectively; in addition, the differential methylation of CpG sites can be influenced by social and environmental exposures between ethnic sub-groups. Thus, these authors concluded that the methylation of CpG sites might be influenced by a combination of environmental exposures, social characteristics, and genetic ancestry.
In this study, we found a relationship between pesticide exposure and the methylation patterns of the CDKN2B and CDKN2A genes in a genetically conserved population. To date, some researches have been performed to study the epigenetic modifications by pesticide exposure and their implications and, although some of the limitations of our study included that we did not evaluate the mRNA levels of CDKN2B and CDKN2A genes in the study population, to our knowledge, this is the first study to evaluate the gene-specific methylation patterns of CDKN2B and CDKN2A genes in a genetically conserved population exposed to pesticides. This study opens the way to additional questions related to methylation patterns in these types of genetically conserved populations, and to questioning focused on the possible interactions among ethnicity, pesticide exposure, diet, sociocultural context, and others factors. These findings provide a bases for the impact of pesticide exposure on epigenetic modifications in genetically conserved populations, as well as the important perspectives that are emerging with regard to identifying potential differentially methylated CpG sites that could be associated with increased vulnerability to disease development in these populations. Additionally, it is important to conduct future longitudinal studies on methylation profiles and the expression of specific genes in populations exposed to pesticides with an emphasis on ethnicity and ancestry.
Supplemental Material
sj-pdf-1-het-10.1177_09603271211063161 – Supplemental Material for Methylation patterns of the CDKN2B and CDKN2A genes in an indigenous population exposed to pesticides
Supplemental Material, sj-pdf-1-het-10.1177_09603271211063161 for Methylation patterns of the CDKN2B and CDKN2A genes in an indigenous population exposed to pesticides by Diana M Paredes-Céspedes, Yael Yvette Bernal-Hernández, José Francisco Herrera-Moreno, Aurora Elizabeth Rojas-García, Irma Martha Medina-Díaz, Cyndia A González-Arias and Briscia Socorro Barrón-Vivanco in Human & Experimental Toxicology
Footnotes
Acknowledgments
We thank all the workers that participated in this study. The authors also thank Leticia Yáñez-Estrada, PhD and her team at the Autonomous University of San Luis Potosí, Mexico for providing the support for DAP determination. Also, we thank Isabel Alvarado-Cruz, PhD and Betzabet Quintanilla-Vega, PhD of the Toxicology Department (CINVESTAV-IPN) for their support during the determination of methylation data.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by CONACyT grants [262284] and Red Temática de Toxicología de Plaguicidas [294303].
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.