
Abstract
The presence of 5-HT7r’s in both human and rat cardiovascular and immune tissues and their contribution to inflammatory conditions prompted us to hypothesize that these receptors contribute in acute myocardial infarction (MI) with underlying chronic endothelial dysfunction. We investigated the role of 5-HT7 receptors on heart tissue that damaged by isoproterenol (ISO)-induced MI in rats with high-fat diet (HFD). In vitro and in vivo effects of 5-HT7r agonist (LP44) and antagonist (SB269970) have been investigated on the H9C2 cell line and rats, respectively. For in vivo analyses, rats were fed with HFD for 8 weeks and after this period ISO-induced MI model has been applied to rat. To investigate the role of 5-HT7r’s, two different doses of LP44 and SB269970 were evaluated and compared with standard hypolipidemic agent, atorvastatin. In vitro studies showed that LP44 has protective and proliferative effects on rat cardiomyocytes. Also in in vivo studies stimulating 5-HT7r’s by LP44 improved blood lipid profile (decreased total cholesterol, low-density lipoprotein-C, and triglyceride, increased high-density lipoprotein), decreased cardiac damage markers (creatine kinase and troponin-I), and corrected inflammatory status (tumor necrosis factor-α, interleukin-6). Our results showed significant improvement in LP44 administered rats in terms of histopathologic analyses. In damaged tissues, 5-HT7 mRNA expression increased and agonist administration decreased this elevation significantly. We determined for the first time that 5-HT7r’s are overexpressed in ISO-induced MI of rats with underlying HFD-induced endothelial dysfunction. Restoration of this overexpression by LP44, a 5-HT7r agonist, ameliorated heart tissue in physiopathologic, enzymatic, and molecular level, showing the cardiac role of these receptors and suggesting them as future potential therapeutic targets.
Introduction
Atherosclerosis, the principal contributor to the pathogenesis of myocardial infarction, is known to be one of the main causes of morbidity and mortality worldwide. 1 High lipid levels, including triglycerides (TGs), non-esterified fatty acids, and low-density lipoprotein (LDL-C), damage vascular tissues and produce endothelial dysfunction. 2 Cardiovascular research suggests that endothelial dysfunction is the initial step in the development of atherosclerosis. 3 Dysfunction of the arterial vasculature is a main contributor to atherosclerotic cardiovascular disease and the initiation and progression of atherosclerosis. 4,5 Endothelial dysfunction caused by lipotoxicity is mediated through several mechanisms that include increased oxidative stress and pro-inflammatory responses. 6 Endothelial dysfunction also increases the risk of myocardial infarction (MI) that is also associated with high levels of pro-inflammatory response. 7
Serotonin (5-HT) is released during thrombocyte activation. It mediates thrombocyte-related vasoconstriction and activation of other thrombocytes that contribute to thrombotic complications and coronary artery disease progression. 8 LDL has also been shown to increase 5-HT release from thrombocytes and stimulate 5-HT-induced thrombocyte aggregation in human plasma. 9 Namely, 5-HT contributes to atherosclerotic lesions. Besides, 5-HT has both vasoconstrictive and vasodilator effects on coronary arteries 10,11 ; injecting low doses of 5-HT into normal human coronary arteries induces vascular dilation, but at high doses 5-HT injections cause vasoconstriction. 10 The vasoconstrictive effect of 5-HT on coronary arteries is mostly mediated by 5-HT2A receptor, and to a lesser extent by the activation of 5-HT1B receptor. 12,13 5-HT7 receptor is the latest identified subtype of 5-HT receptors. 14 It couples to Gs protein and induces cAMP accumulation when activated, 15 and in rat glomerulosa cells the activation of 5-HT7receptor increases calcium influx via T-type Ca2+ channels by raising adenylyl cyclase activity. 16 5-HT7 receptors have also been shown in arteries such as coronary arteries, aorta, and pulmonary. 13,17 Activation of 5-HT7 receptor has been reported to mediate 5-HT-induced relaxation in isolated dog coronary arteries 18 and 5-HT-induced coronary flow increase has been blocked by the selective 5-HT7 receptor antagonist SB269970. 19 Previous studies showed the presence of 5-HT7 receptor mRNA in both human and rat immune tissues such as thymus, spleen, peripheral lymphocytes, and mitogen-activated spleen cells. 20 The presence of 5-HT7 receptors in both human 21 and rat 22 cardiovascular 23 and immune tissues and its contribution to inflammatory conditions 24 –27 prompted us to hypothesize that these receptors contribute in acute MI with underlying chronic endothelial dysfunction as a rebound mechanism to increase coronary outflow and decrease inflammatory response.
In the light of above information, this study aimed to investigate the role of 5-HT7receptors, whose role have been shown in both endothelium and vascular smooth muscle, on heart tissue that damaged by isoproterenol (ISO)-induced MI in rats with high-fat diet (HFD) exacerbated coronary endothelial dysfunction
Materials and methods
In vitro cell proliferation and viability analysis
The embryonic cardiomyocyte cell line H9C2 was provided from American Type Culture Collection (ATCC, USA). The cell lines in Cryotube in the liquid nitrogen were removed from the tank and allowed to dissolve in the water bath at 37°C for a short time. Soluble cells were transferred to flasks with T75 cm2. After 48 h, H9C2 cells were counted at 2 × 105 cells/well in DMEM containing 10% FBS and seeded in a 96-well plate and incubated at 37°C in a humid atmosphere containing 5% CO2. The experimental groups are control, ISO, ISO + AGO1, ISO + AGO2, ISO + ANTA1, ISO + ANTA2, ISO + AGO2 + ANTA2, and ISO + ATOR. After 24 h, cells were exposed to LP44 (5-HT7 receptor agonist) and SB269970 (5-HT7 receptor antagonist) at different concentrations (10−6 and 10−4 M), atorvastatin (10 μmol/l) for 30 min before ISO (10 µM) administration. After the MTT method was applied to the cells at 24 h, the absorbance values of 570 nm were measured three times with microplate reader spectrophotometer (Epoch Microplate Spectrophotometer, BioTek Instruments, Winooski, USA). Inhibition of cell growth was calculated as the percentage survival of treated cells over control cells × 100 (T/C %).

Animals
One hundred male Albino Wistar rats weighing 200–220 g was used in the study. The experiments were conducted according to the ethical norms approved by the Ethics Committee of the Experimental Animal Teaching and Research Center (No: 36643897-100-E.1600080347). The rats were obtained from the Medicinal and Experimental Application and Research Center, Erzurum, Turkey (ATADEM). They were kept in standard laboratory conditions under a natural cycle of light and dark.
Experimental protocols of ISO-induced MI
The rats were divided into 10 groups (each consisting of ten rats) as follows: Healthy Healthy + ISO HFD HFD + ISO (100 mg/kg) HFD + ISO (100 mg/kg) + LP44 1 (1 mg/kg) HFD + ISO (100 mg/kg) + LP44 2 (2 mg/kg) HFD + ISO (100 mg/kg) + SB269970 1 (1 mg/kg) HFD + ISO (100 mg/kg) + SB269970 2 (2 mg/kg) HFD + ISO (100 mg/kg) + LP44 2 + SB269970 2 (2 mg/kg) HFD + ISO (100 mg/kg) + atorvastatin (10 mg/kg)
Except for groups 1 and 2, all groups were fed with HFD (code: C1061-Altromin Spezialfutter GmbH & Co. KG, Lage, Germany) (Table 1) for 8 weeks for induction of hypercholesterolemia. On the last day of the 8-week period, LP44 (Sigma Chemical Company, Germany) and SB269970 (Sigma Chemical Company, Germany) were injected into selected groups (i.p.). The calculated dose (100 mg/kg) 28 of ISO (Sigma Chemical Company, Germany) was dissolved in 1 ml of distilled water (injectable water) and injected (s.c.) 2 h after LP44 (5-HT7 receptor agonist) and SB269970 (5-HT7 receptor antagonist) administration. 29 Agonist and antagonist were administered at 1 and 2 mg/kg doses. 30 Atorvastatin was administered at 10 mg/kg dose. 31 We used atorvastatin in our study to compare effects of 5-HT7 receptors with an anti-atherogenic agent in MI induced damage in rats with underlying atherosclerosis. All control rats received distilled injectable water as vehicle for all applications. Twelve hours after ISO injection, all the rats were anesthetized with i.m. ketamine (30 mg/kg) and xylazine (6 mg/kg). Blood samples were collected and serum separated after centrifugation. Serum was used for various biochemical estimations. The heart was dissected out and washed immediately in ice-chilled saline. The heart tissue was homogenized. The homogenate was centrifuged at 3000 r/min for 5 min. The supernatant was used for the estimation of various biochemical parameters.
Consumption of HFD (C1061-atherogenic diet, 1% cholesterol, and 1% Na-cholate, Altromin Spezialfutter GmbH & Co. KG, Lage, Germany, info@altromin.de—www.altromin.com).

a Detailed content is available as supplementary table.
Biochemical estimations
The separated serums were used for the determination of aspartate aminotransferase (AST), glucose, total cholesterol (TC), TGs, HDL-C (high-density lipoprotein), LDL-C, and creatine kinase (CK) levels with an auto-analyzer (AU 5800, BECHMAN-COULTER Diagnostic System, West Sacramento, California, USA) and for determination of troponin-I (Tn-I) levels with another auto analyzer (Access2).
Sodium–potassium-dependent adenosine triphosphatase (Na+/K+ ATPase) and calcium-dependent adenosine triphosphatase (Ca2+ ATPase)
The protein concentration in the supernatant was determined using the Bradford method. Levels of Na+-K+-ATPase (cat. no: 201-11-2666) and Ca2+-ATPase (cat. no. 201-11-2665) activities in whole heart tissues were measured with an ELISA kit (SunRed Bio, Shanghai, China) in accordance with the manufacturers’ instructions.
Total RNA extraction and cDNA synthesis
The whole heath tissues (20 mg) were stabilized in RNA Stabilization Reagent (RNAlater, Qiagen, Hilden, Germany), and then disrupted using a TissueLyser II (Qiagen). The total RNA was purified using an RNeasy Mini Kit (Qiagen) in accordance with the manufacturer’s instructions in a QIAcube (Qiagen). The RNA samples were then reverse-transcribed into complementary DNA with a High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, California, USA). The cDNA concentration and quality were assessed and quantified using the Epoch Spectrophotometer System and Take3 Plate (BioTek).
Relative quantification of gene expression (real-time reverse transcriptase-polymerase chain reaction)
The relative interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), transforming growth factor beta1 (TGF-β1), and 5-HT7receptor expression analyses were performed with StepOnePlus Real-Time PCR System technology (Applied Biosystems) using cDNA synthesized from rat RNA, as previously described. 24 All primers and probes used were purchased as TaqMan Gene Expression Assays: IL-6 (F: TCC ATC CAG TTG CCT TCT TG; R: GGT CTG TTG GGA GTG GTA TC), TNF-α (F: GCT CCC TCT CAT CAG TTC CA; R: CTC CTC TGC TTG GTG GTT TG), TGF-β1 (F: CGCAACAACGCAATCTATGAC; R: GGA CAG CAA TGG GGG TTCT), and 5-HT7 receptor (F: AGACTGCTCAAACACGAAAGG; R: AAG GCT CCC ACG ATG ATC C) (Applied Biosystems). The expression data for β-actin (F: TGG TGG GTA TGG GTC AGA AG; R: GAC AAT GCC GTG TTC AAT GG) in each tissue was used as the endogenous control, 32 –34 and the primers and probes for the β-actin were designed by Primer Design.
Histological procedure
Heart tissue samples were obtained at necropsy and fixed in a 10% formaldehyde solution. Then, routine pathological processing was done and tissues were embedded in paraffin wax. From the paraffin wax block of each lung sample, 4–5 µm slices were cut. After hematoxylin and eosin staining (H&E), the ventricles were examined by a pathologist who is blinded to study groups and equipped with a digital camera (Olympus BX51, DP25 camera, Japan) using light microscopy. The following histopathological parameters were accepted for scoring according to the previous literature 35 : cell infiltration, necrosis, hemorrhage. A minimum of five fields for each heart section at ×200 magnification were evaluated and assigned for severity of changes using scores on a scale of grade 0: − (negative), grade 1: +1 (mild), grade 2: +2 (moderate), grade 3: +3 (severe), grade 4: +4 (most severe).
Statistical analysis
Values are expressed as the mean ± SD. The numerical data from biochemical analyses and real-time polymerase chain reaction (PCR) results was analyzed using the one-way analysis of variance post hoc Duncan test. SPSS-17 for Windows was used for all statistical analyses. p Values less than 0.05 were considered significant.
Results
Proliferative effect of LP44 in H9C2 cells
As shown in Figure 1, 10 μM ISO significantly decreased the number of H9C2 cells at 24 h time. When we administered LP44 with ISO, LP44 prevented the cardiomyocytes from ISO damage dose dependently. Also SB269970 had positive effects on cell proliferation, but this effect was not as strong as LP44 had. Even the effect of SB269970 decreased as its dose increased. Atorvastatin also prevented ISO-induced cell death as well as LP44 showing beneficial effects of both LP44 and atorvastatin on rat cardiomyocytes.

Effects of LP-44, SB269970, and ISO on the viability of H9C2 cells at 24 h. MTT shows significant differences between experimental groups compared to control. ISO (10 μM), LP44 1 (10-6 M), LP44 2 (10-4 M), SB269970 1 (10-6 M), SB269970 2 (10-4 M). Values not sharing a common superscript differ significantly at p < 0.05 DMRT. ISO: isoproterenol; DMRT: Duncan’s Multiple Range Test.
Biochemical results
As seen in Figure 2, serum glucose and AST levels were significantly increased in the HFD and HFD + ISO groups when compared to the healthy group. LP44 administration decreased both glucose and AST levels in a dose-dependent manner. Atorvastatin was also able to decrease serum glucose and AST levels. 2 mg/kg dose of SB269970 increased serum glucose level while 1 mg/kg dose of SB269970 decreased serum glucose level when compared to HFD and HFD + ISO groups. Any dose of SB269970 did not cause a significant difference in terms of AST levels. When we administered LP44 and SB269970 at 2 mg/kg doses together, they blocked each other’s effect.

Effect of HFD on the activities of serum: (a) glucose and (b) AST in normal and ISO-treated rats. HFD, ISO (100 mg/kg), LP44 1 (1 mg/kg), LP44 2 (2 mg/kg), SB269970 1 (1 mg/kg), SB269970 2 (2 mg/kg), ATOR. Values not sharing a common superscript differ significantly at p < 0.05 DMRT. ISO: isoproterenol; DMRT: Duncan’s Multiple Range Test; HFD: high-fat diet; ATOR: atorvastatin.
Figures 3 show that serum TC, LDL-C, and TG lipid levels were increased significantly in the sera of HFD groups. ISO administration to both healthy rats and HFD rats increased these parameters. As an opposite marker in terms of hyperlipidemia, HDL-C levels were decreased in rats which received HFD. Atorvastatin and LP44 administration ameliorated serum lipid profile significantly. SB269970 also decreased TC and LDL-C levels but did not affect and/or increased TG levels in rats with HFD + ISO. SB269970 administration also reversed the ameliorative effect of LP44 on serum lipid levels.

Effect of HFD on the activities of serum (a) TC, (b) LDL-C, (c) HDL-C, and (d) TG in normal and ISO-treated rats. HFD, ISO (100 mg/kg), LP44 1 (1 mg/kg), LP44 2 (2 mg/kg), SB269970 1 (1 mg/kg), SB269970 2 (2 mg/kg), ATOR. Values not sharing a common superscript differ significantly at p < 0.05 DMRT. ISO: isoproterenol; DMRT: Duncan’s Multiple Range Test; HFD: high-fat diet; ATOR: atorvastatin; TC: total cholesterol; LDL: low-density lipoprotein; TG: triglyceride; HDL: high-density lipoprotein.
On the other hand, both HFD and ISO administration increased cardiac markers (CK, Tn-I) significantly. When we applied HFD and ISO together, the increase of cardiac markers was much more evident. LP44 decreased both of the cardiac markers significantly (Figure 3(a) to (b)). In terms of cardiac markers, different from serum lipid profile, both doses of SB269970 increased CK and Tn-I significantly when compared to HFD+ISO group. Also, SB269970 blocked the ameliorative effects of LP44 on these markers (Figure 4(a) to (b)).

Effect of HFD on the activities of serum (a) CK, (b) Tn-I and activities of heart tissue ELISA, (c) Na+-K+-ATPase, and (d) CA+2 ATPase in normal and ISO-treated rats. HFD, ISO (100 mg/kg), LP44 1 (1 mg/kg), LP44 2 (2 mg/kg), SB269970 1 (1 mg/kg), SB269970 2 (2 mg/kg), ATOR. Values not sharing a common superscript differ significantly at p < 0.05 DMRT. ISO: isoproterenol; DMRT: Duncan’s Multiple Range Test; HFD: high-fat diet; ATOR: atorvastatin; Tn-I: Troponin-I; CK: creatine kinase.
In addition, the effects of 5-HT7 agonist and antagonist on the activities of Na+-K+-ATPase and Ca2+-ATPase (μmol of phosphorous liberated/h/mg protein) in the heart of normal and ISO-treated rats were evaluated. The activity of Na+-K+-ATPase significantly decreased, while the activity of Ca2+-ATPase significantly increased in the heart tissue of HFD, ISO, and HFD + ISO-treated rats (Figure 4(c) to(d)). LP44 was able to correct the changes in these parameters dose dependently. Especially high dose of SB269970 administration worsened the status of these enzymes in cardiac tissue. Atorvastatin administration corrected all cardiac markers when compared HFD + ISO group.
Real-time PCR results
The inflammatory cytokines TNF-α (Figure 5(a); p ≤ 0.05), IL-6 (Figure 5(b); p ≤ 0.05), and TGF-β1 (Figure 5(c); p ≤ 0.05) increased in the HFD + ISO-treated group when compared to the healthy control group. LP44 dose dependently reversed these elevations induced by HFD + ISO. For SB269970, the effects were a little bit confusing. While 1 mg/kg SB269970 decreased TNF-α mRNA expression, 2 mg/kg significantly increased this parameter when compared to HFD + ISO group. Also, SB269970 significantly decreased IL-6 and TGF-β1 mRNA expression levels while this effect was not as strong as LP44 did. Also combined administration of these two agents decreased levels of all cytokines; however, this effect was not still similar to that of LP44. SB269970 seems to be acting as a partial antagonist that exerts anti-inflammatory effect alone but blocks the effect of LP44 in combined administration. Atorvastatin also decreased the levels of these cytokines when compared to HFD + ISO group.

Effect of HFD on the activities of heart tissue mRNA levels: (a) TNF-α, (b) IL-6, (c) TGF-β1 and 5-HT7 receptor in normal and ISO-treated rats. HFD, ISO (100 mg/kg), LP44 1 (1 mg/kg), LP44 2 (2 mg/kg), SB269970 1 (1 mg/kg), SB269970 2 (2 mg/kg), ATOR. Values not sharing a common superscript differ significantly at p < 0.05 DMRT. ISO: isoproterenol; DMRT: Duncan’s Multiple Range Test; HFD: high-fat diet; ATOR: atorvastatin; IL-6: interleukin-6; TNF-α: tumor necrosis factor-α; TGF-β1: transforming growth factor beta1.
5-HT7 receptor mRNA expression was increased in both the HFD and ISO groups when compared to the healthy group (Figure 5(d)). Both doses of LP44 treatment decreased 5-HT7 receptor mRNA expression when compared to the HFD + ISO group. However, SB269970 administration did not change this expression at a low dose and even increased it at a high dose. Atorvastatin also decreased 5-HT7 receptor mRNA expression in line with its other beneficial effects.
Histopathological results
The heart tissue of the animals in the control group showed normal histological features (Figure 6(a)). Despite severe cell infiltration, necrosis, and hemorrhage in HFD and HFD + ISO heart tissue, this was observed to be more severe in the HFD + ISO group (Figure 6(b) and (c)). Healthy + ISO had histology similar to HFD + ISO, but less cell infiltration, necrosis, and hemorrhage were observed (Figure 6(d)). When we looked at the HFD + ISO + AGO1 and HFD + ISO + AGO2 groups (Figure 6(e) and (f)), cell infiltration, necrosis, and hemorrhage decreased significantly compared to the healthy + ISO group. Compared to the HFD + ISO group, it was observed that there was a much more meaningful correction. In particular, the improvement in the HFD + ISO + AGO2 group is more evident in terms of infarction, necrosis, and hemorrhage than in the HFD + ISO + AGO1 group. When we examined the HFD + ISO + atorvastatin group (Figure 6(j)), cell infiltration was significantly reduced compared to the HFD + ISO group, and hemorrhage was similar to that of the HFD + ISO + AGO1 group. When the HFD + ISO + ANTA1 and ANTA2 groups (Figure 6(g) and (h)) were compared with HFD + ISO, similar results were obtained. There was intense cell infiltration, necrotic areas in many slides, and severe hemorrhage. The histopathological findings of the HFD + ISO + AGO2 + ANTA2 group were similar to those of the HFD + ISO group, with a high rate of cell infiltration necrosis and hemorrhage observed (Figure 6(i)).
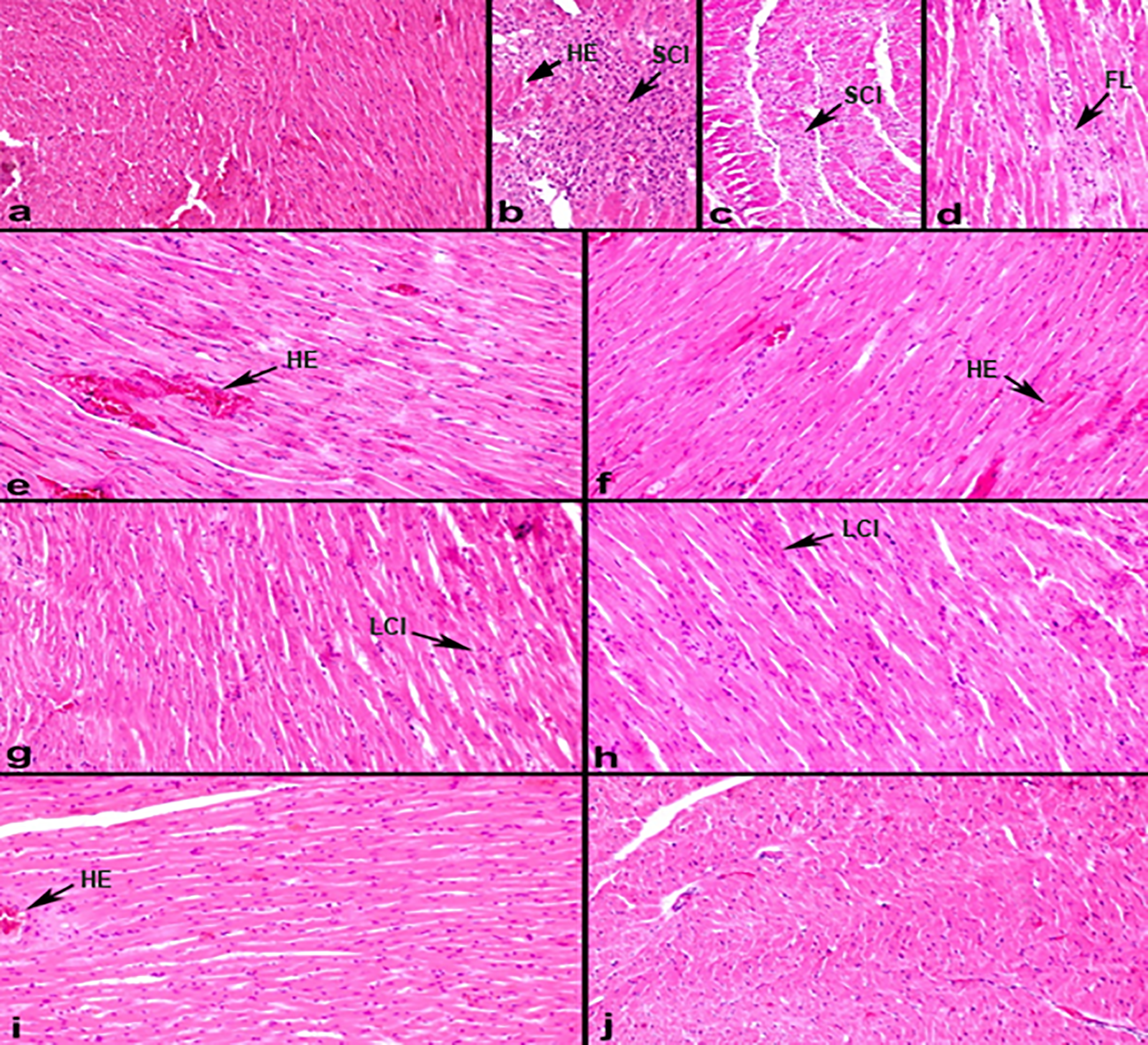
(a) Healthy, (b) HFD, (c) HFD + ISO, (d) healthy + ISO, (e) HFD + ISO + LP44 1(1 mg/kg), (f) HFD + ISO + LP44 2 (2 mg/kg), (g) HFD + ISO + SB269970 1 (1 mg/kg), (h) HFD + ISO + SB269970 2 (2 mg/kg), (i) HFD + ISO + LP44 2 (2 mg/kg) + SB269970 2 (2 mg/kg), and (j) HFD + ISO + ATOR. ISO: isoproterenol; HE: hemorrhage; SCI: severe cell infiltration; HFD: high-fat diet; FL: fewer lesions; LCI: less cell infiltration—Hematoxylin and eosin staining. (a) to (c) ×100 magnification, (e) to (j) ×200 magnification.
Discussion
In spite of numerous studies regarding MI, the high rate of mortality associated with MI still remains a major problem today. Hyperlipidemia-induced atherosclerosis is among the major causes of MI. In this study, the possible protective effects of 5-HT7 agonist (LP44) and antagonist (SB269970) on ISO-induced MI in hyperlipidemic rats were investigated biochemically, molecularly, and histopathologically. Also, the effects of LP44 and SB269970 on heart tissue and H9C2 rat cardiomyocyte cell line were shown for the first time. Our study was also the first to show that HFD increases 5-HT7 receptor mRNA levels in the heart. HFD is known to increase endothelial damage and cause atherosclerosis in long term. 2 In our study, we found increased 5-HT7 receptor expression in HFD groups with endothelial damage as an important finding. In addition to the HFD, MI induction and ischemic damage by ISO further exacerbates this increase. However, it is important to know whether overexpression of 5-HT7 receptors during HFD + ISO-induced cardiac damage is a defense mechanism against damage or harmful factor contributing to damage. To answer this point, we focused on the groups received 5-HT7 receptor agonist and/or antagonist. In the group received HFD + ISO + LP44 all pathologic findings and also inflammatory cytokines were decreased in parallel to decreased 5-HT7 receptor expression. When we checked SB269970-administered HFD + ISO groups, no pathological healing observed in the heart tissue in parallel to increasing 5-HT7 overexpression though some contradictory results obtained from biochemical and inflammatory analyses. This situation can be resulted from either antagonist dose was not adequate to bind overexpressed 5-HT7 receptors or any other endogenous mechanisms that stimulate 5-HT7 receptor expression contribution. 5-HT7 agonist, reduced tissue damage, oxidative stress, and inflammatory cytokines which are markers of heart damage at all doses administered. Protective effect of LP44 on heart tissue also confirmed by in vitro analyses that performed on rat cardiomyocytes.
5-HT mediates the activation of platelet-derived vasoconstriction and other platelets and may accelerate the progression of coronary artery disease by initiating thrombotic complications. 36 Also serum 5-HT levels were found to increase during ischemic heart conditions 37 and atherosclerosis. 38 While five of the defined 14 5-HT receptor subtypes are involved in central cardiovascular regulation, studies on the other nine subtypes continue to validate their potential role in this system. Our study examined the agonist and antagonist of 5-HT7 receptors in ISO-induced MI with underlying atherosclerotic endothelial dysfunction. In the study, a HFD was applied to rats for 8 weeks to induce hyperlipidemia. 29
Xu et al. have shown that serum glucose is increased due to hyperlipidemia 39 and that ISO application leads to an increase in blood glucose level both by direct participation of β1 and β2 adenoceptors and by adrenergic innervation. 40 In our study, increased blood glucose was reduced by the agonist, indicating that 5-HT7 receptors are involved in MI that occurs in hyperlipidemic animals. As another routine biochemical parameter, AST is a reliable constructive enzyme in animal research. In our study, the 5-HT7 receptor agonist significantly reduced the level of AST compared to the HFD + ISO group, which supports the possible protective effect of these receptor agonists.
Hyperlipidemia, clinically expressed by high levels of TC, TG, and LDL-C and low levels of HDL-C, plays a vital role in atherosclerosis and cardiovascular diseases attributed to defective lipid metabolism systems. 41 A HFD is the most effective cause of the elevation of TC, TG, and LDL-C and the reduction of HDL-C. 42 It has been reported that with ISO application, the levels of TG, TC, LDL-C, and VLDL-C are increased and the levels of HDL-C are decreased. 43,44 In parallel with these studies, we found that serum TG, TC, LDL-C, and VLDL-C levels of ISO and ISO + HFD groups were higher and HDL-C levels were lower than that of healthy control group. Both doses of agonist reversed the impaired lipid profile. In previous studies, the induction of hyperlipidemia and/or hyperglycemia by 5-HT receptor antagonists 45,46 also promoted the protective effect of the 5-HT7 receptor agonist LP44. 5-HT7 receptors are membrane bound G coupled receptors and can be expressed in all tissue types. Previously, Sjogren et al. demonstrated that cholesterol depletion reduces binding of SB269970 to 5-HT7 receptors and counteracts 5-HT7-mediated intracellular signaling. 47 This made us to suggest that 5-HT7 receptors can directly interact with cholesterol and membrane lipids in all tissues, resulting in direct depletion in serum lipid profile after agonist stimulation. However, future detailed studies are required for evaluation of this interaction and underlying mechanism.
Clinically, the enzymes and proteins released by cardiomyocytes into the membrane injury endocardium can be used to identify cardiac problems—the most commonly used are CK, CK-MB, and Tn-I. These biochemical markers are known as the gold standard in the diagnosis of patients with myocardial insufficiency in which the ECG is normal. 48 CK-MB, which is highly specific for the heart, may also increase due to iso-enzymatic skeletal injury. Several studies have shown that CK, CK-MB, and Tn-I are elevated in rats with ISO application. 49,50 In our study, it was also shown that CK, CK-MB, and Tn-I were increased by ISO application when compared with the healthy group, and the present situation in HFD groups was further exacerbated. While the agonist application reversed CK, CK-MB, and Tn-I levels as well as atorvastatin.
Changes in the properties of membrane-bound enzymes affect the functioning of the heart. Researchers pointed out that the failure of the cell membrane to maintain its normal transmembrane ionic distribution via ion pumps is an important event in the pathogenesis of ischemia and arrhythmias. 51 In our study, it was observed that Ca2+-ATPase activity was increased while Na+-K+-ATPase activity was decreased in ISO-treated rat hearts. Increased Ca2+-ATPase activity and decreased Na+-K+-ATPase activity in HFD- and ISO-treated rats may be due to increased lipid peroxidation of free radicals. 52 In our study, agonist treatment protected ATPase activities at normal and/or near-normal levels in HFD- and ISO-treated rats. The reason why 5-HT7 receptor stimulation change cardiac marker can also be related to the changes in blood pressure during MI. However, previously Anwar et al. reported that blockade of 5-HT7 receptors by SB269970 unaffected tryptamine-induced vasodilator responses. 53 So we suggested that the protective effects of 5-HT7 receptor stimulation is not directly related with blood pressure but can be related with improved cardiac enzymes.
Studies have shown that TGF-β1 may be an important molecule in cardiac embryogenesis, hypertrophy, atherogenesis, improvement of myocardial infarction, and development of coronary collaterals. 54,55 In our study, it was observed that TGF-β1 mRNA levels were increased by ISO application, and this condition was further exacerbated in HFD groups. The agonist administration in our treatment groups resulted in a significant decrease, especially in our HFD + ISO + AGO2 group. This decrease in TGF-β1 mRNA levels due to agonist administration suggests that 5-HT7 receptors may also play a role in the development of cardiac damage.
There is an increase in pro-inflammatory cytokines, the flow of inflammatory cells such as monocytes, macrophages, lymphocytes, and neutrophils, in the infarct region during MI. 56,57 Relatively, there are studies demonstration the relationship between 5-HT7 receptors and immune system. Previously, 5-HT7 receptors were found to be expressed in immature T cells and 5-HT7 receptor stimulation increase T cell function and activation. 58 Also 5-HT has been shown to prevent monocytic apoptosis in these cells via 5-HT7 receptors. 20 Stefulj et al. investigated 5-HT7 receptor expression in rat immune tissues and determined 5-HT7 receptor mRNA in thymus, peripheral leucocytes, spleen, and mitogen-activated immune system cells. 22 We also observed that the levels of TNF and IL-6 mRNA significantly increased in the HFD and HFD + ISO groups in our study and significantly decreased in the HFD + ISO + AGO2 group. This effect, which is observed in parallel with anti-inflammatory impact, is also supported by previous studies. Albayrak et al. found that 5-HT7 receptor agonist showed anti-inflammatory and anti-cytokine effects and that 5-HT7 receptors may be a novel therapeutic target in the prevention of inflammation and inflammatory diseases. 27 In addition, Cadirci et al. have supported the anti-inflammatory activity 5-HT7 receptor agonist during sepsis. 26 The induction of 5-HT7 receptors in our study may be due to the effects of anti-inflammatory and related anti-cytokines, which may produce beneficial effects in the post-HFD-aggravated MI model. The 5-HT7 receptors were mainly discovered in the central nervous system, 59 but subsequently, the peripheral effects were also frequently investigated. 60,61 However, no study thus far has investigated the pre- and post-injury levels of 5-HT7 receptors in cardiac tissue. In our study, we examined the presence of 5-HT7 receptors in heart tissue in rats that were fed with HFD, followed by ISO and MI models and the damage-dependent alteration of these receptors. Increasing levels of 5-HT7 receptors in both the HFD and HFD + ISO groups and with the LP44 administration, both damage reduction and reduction of receptor expression suggest that these receptors may have increased as a defense mechanism against injury in the heart tissue.
In conclusion, this study showed that HFD increases the 5-HT7 receptor mRNA expression in heart tissue for the first time. Combination of HFD with ISO-induced MI aggravated this increase of 5-HT7 receptors. We determined that 5-HT7 receptors are overexpressed in ISO-induced MI of rats with underlying HFD-induced endothelial dysfunction. Restoration of this overexpression by LP44, a 5-HT7 receptor agonist, ameliorated heart tissue in physiopathologic, enzymatic, and molecular level, showing the cardiac role of these receptors and suggesting them as future potential therapeutic targets.
Footnotes
Author's note
Z Halici is also affiliated with Clinical Research, Development and Design Application and Research Center (KARGETAM), Ataturk University, 25240 Erzurum-Turkey.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.