
Abstract
Modulation of myometrial spontaneity by cadmium (Cd) and its regulatory pathways was studied in rat uterus in the absence and presence of blockers of different signaling pathways. Isometric tension in myometrial strips, under a resting tension of 1 g, mounted in organ bath containing Ringer–Locke solution (RLS) continuously aerated with carbogen, was measured using data acquisition system-based physiograph and Lab Chart Pro V7.3.7 software. Mean integral tension was measured for 8 min. Cd (1 nM–0.1 mM) not only produced concentration-dependent inhibitory effect on rat myometrium but it (10 µM) also significantly (p < 0.05) inhibited calcium chloride and BAY K-8644-induced myometrial contraction. Glybenclamide (10 µM), 4-aminopyridine (1 mM), and propranolol (10 µM) failed to significantly attenuate Cd-induced inhibitory responses, while L-NAME (0.1 mM), 1H-[1,2,4]Oxadiazolo[4,3-a]quinoxalin-1-one (ODQ; 25 µM), and 9-(tetrahydro-2-furanyl)-9H-purin-6-amine (SQ 22536; 1 µM) significantly (p < 0.05) produced inhibitory effects on Cd-induced myometrial relaxation. Phenylephrine (1 nM–10 µM) and salbutamol (0.01 nM–0.1 µM)-induced relaxant effects on rat myometrium were significantly potentiated by 10 µM Cd. Thus based on the results of present functional study, it may be inferred that inhibitory effects of Cd on rat myometrium are mediated through blockade of L-type calcium channels and activation of NOS-NO-sGC and/or AC-cAMP pathways.
Introduction
Cadmium (Cd) is one of the most abundant and potent hazardous metallic environmental pollutant and the geometric blood level of Cd in general population of ≥1 year of age is 0.315 µg/L. 1 Exposure to Cd occurs primarily through ingestion of contaminated water, food, and to a significant extent through inhalation and cigarette smoking. 2 Apart from cancer, immune, respiratory, and cardiovascular disorders, 2 Cd results in reproductive defects like subfertility or abortions in humans 3 and animals. 4 Cd accumulates in human endometrial tissue and its high levels have been reported in females with the history of smoking. 5 Smoking during pregnancy increases the contractile sensitivity and activity of the myometrium to oxytocin by upregulating oxytocin receptor mRNA and increases the risk of premature delivery in cigarette-smoke-exposed Wistar rats. 6 Cd inhibits angiogenesis in endometrial cells which results in endometrial dysfunctions and increased risk for fertility problems. 7
Cd produces inhibitory effect on human myometrium spontaneity while increases and decreases the response of Ca2+ and oxytocin at lower (1 nM–10 nM) and higher concentrations, respectively, 8 and inhibitory effect on bovine myometrium. 9 On guinea pig ileum, it has been reported to produce both the inhibitory and excitatory effects. 10,11 But its dose-dependent differential effects, that is, stimulatory at lower concentration and inhibitory at higher concentration have been observed on rat thoracic aorta. 12 Cd contracts ileal longitudinal muscle through release of cholinergic transmitter from parasympathetic-nerve terminals, which depends on external Ca2+ apart from smaller hyoscine-resistant contractile effect, presumably due to direct action on smooth muscle cells. 11 Chronic Cd intoxication at 5 mg/kg body weight resulted in an increase in norepinephrine response in rat mesenteric arteries 13 while endothelin, noradrenaline, and angiotensin II achieved lower maximal contraction in Cd-induced hypertensive rat aorta as compared to normotensive rats. 14 But in vitro Cd (10 µM) treatment for 24 h augmented α1-adrenoceptor-mediated contractile response in rat aorta even though endothelial nitric oxide synthase (eNOS) and inducible nitric oxide synthase (iNOS) were upregulated by Cd. 15 Takahashi et al. 15 also observed increase of nitric oxide (NO)-induced and β-adrenoceptor-mediated relaxing responses by Cd treatment in rat aorta. Thus, the regulatory mechanism(s) of Cd seem to vary not only with concentration but also the type of smooth muscles (myometrium, ileum, aorta, mesenteric artery) and between species, namely human, bovine, guinea pigs, and rats.
Ion transport and signal transduction, which include intracellular mobilization of second messengers such as inositol triphosphate, 16 inhibition of plasma membrane calcium channels, 17 and inhibition of plasma membrane Ca2+-ATPase, 18 have been identified as the cellular targets of Cd in different smooth muscles. Apparently, there is paucity of detailed information on effect of Cd on myometrial activity, its cellular targets and mechanistic pathways, including cyclic nucleotide-dependent pathways. Hence the present study was undertaken to study the direct effect of Cd and its mechanistic pathways on isolated myometrial strips of rats.
Materials and methods
Chemicals
Cadmium chloride (CdCl2) monohydrate and prostaglandin F2 alpha (PGF2α, lutalyse) were purchased from Himedia (Mumbai, Maharashtra, India) and Pfizer (Mumbai, Maharashtra, India), respectively. Glybenclamide—a specific KATP channel blocker, 4-aminopyridine -a specific Kv channel blocker, 1,4-dihydro-2,6-dimethyl-5-nitro-4-(2-[trifluoromethyl] phenyl) pyridine-3-carboxylic acid methyl ester (BAY K-8644)—a specific L-type calcium channel agonist, Nω-nitro-
Experimental animals
Study was conducted on female albino rats (Rattus norvegicus; strain Wistar) weighing 150–180 g procured from Laboratory Animal Resource Section, Indian Veterinary Research Institute (Izatnagar, Uttar Pradesh, India), and maintained under standard managemental conditions with 12-h dark and 12-h light cycle. An acclimatization period of 15 days was ensured before start of the experiment, to acclimatize the animals to the new environment and also switch over to new diet. Experimental studies were undertaken after approval of the Institutional Animal Ethics Committee, DUVASU, Mathura (approval no. IAEC/DUVASU/20/12 dated 1 December 2012) as per Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA) guidelines.
Selection of animals
Di-estrus stage cyclic rats were selected based on vaginal smear examination 19 and used for experimentation as rat myometrium in di-estrus stage exhibited regular and consistent amplitude, frequency, and mean integral tension (MIT) along with longevity of isolated tissue apart from short stabilization time compared to the myometrium from estrogenized rats. For each experimental protocol, eight to ten animals were used.
Collection of tissue
Cyclic adult female rats during di-estrus stage were ethically sacrificed as per the method described by Ghosh
20
after anesthetizing with pentobarbitone sodium (40 mg/kg body wt i.p.). After decapitation, mid-ventral incision was made and both the uterine horns were dissected out and carefully placed in a petri dish containing aerated RLS having NaCl-154 mM/L; KCl-5.6 mM/L; calcium chloride (CaCl2), 2H2O-2.2 mM/L; NaHCO3-6.0 mM/L;
Mounting of tissue and recording of isometric tension
Myometrial strips were mounted in thermostatically controlled (37.0 ± 0.5°C) organ bath (Ugo Basile, Italy) of 10 ml capacity containing RLS, continuously aerated with carbogen (95% O2 + 5% CO2) under a resting tension of 1 g. During the equilibration period of 30 min, physiological salt solution in tissue bath was changed almost every 10 min. Isometric tension in tissue was measured by force transducer (AD Instruments, Australia) and recorded with the help of data acquisition system-based physiograph using Lab Chart Pro V7.3.7 software (Powerlab, AD Instruments; Australia). Contact period of Cd with isolated myometrial strips in the tissue bath was 8 min. Tissues were exposed to different concentrations of Cd (10−9 to 10−4 M). Myometrial strips were incubated with different antagonists/blockers for a period of 30 min.
Data analysis
Concentration–response curves were constructed by measuring the MIT for 8 min using Labchart pro V 7.3.7 software as per the formula given by Aaronson et al. 21 Maximum contraction (Emax)/maximum relaxation (Rmax) and median effective dose (EC50) values were determined by nonlinear regression analysis using Graph Pad Prism 4.0 (Graph Pad, La Jolla, San Diego, USA) and pD2 value (potency) was calculated as −log of EC50. Results are expressed as mean ± SEM. Multiple mean values were analyzed using two-way ANOVA followed by Bonferroni post-hoc test or one-way ANOVA followed by Tukey’s post-hoc test, while two mean values were analyzed by student’s t test to compare the difference between two different treatments.
Results
Selection of di-estrus stage rat myometrium
Compared to the initial stabilization time, longevity of tissue responsiveness and spontaneity features (amplitude, frequency, and MIT) in isolated myometrial strips mounted in RLS from estrogenized rats, myometrial strips from normally cyclic rats during di-estrus stage were found to possess significantly (p < 0.05) higher amplitude, MIT and longevity, and significantly lower stabilization period as evident from the data summarized in Table 1 and representative physiographic recordings (Figure 1). Therefore, the study was undertaken on isolated myometrial strips of di-estrus stage cyclic rats.
Comparative amplitude, frequency, mean integral tension, stabilization time, and longevity of myometrial strips from estrogenized and cyclic rats during di-estrus stage.

aVertical bars represent SEM. Data were analyzed by student’s t test followed by unpaired t tests.
bp < 0.05 vs. estrus.

Representative physiograph recordings showing normal spontaneity pattern of isolated rat myometrial strips from di-estrus (a) and estrus (b) stage rats, mounted in RLS. RLS: Ringer–Locke solution.
Effect of cadmium on myometrial spontaneity
Following equilibration, rat myometrial strips (n = 16) exhibited consistent and regular pattern of myogenic spontaneity with an average amplitude, frequency, and MIT of 1.24 ± 0.10 g; 3.08 ± 0.43 beats per minute (bpm) and 0.48 ± 0.03 g, respectively. CdCl2 (1 nM–0.1 mM, at 1 log dose unit) produced concentration-dependent inhibitory effect (Figure 2(a)) on rat myometrium. Figure 2(b) illustrates the cumulative dose–response curve (DRC) of CdCl2 on rat myometrium. The pD2 and Rmax values of CdCl2 were calculated to be 5.15 ± 0.15 (n = 8) and 113.30 ± 5.68% (n = 8), respectively.
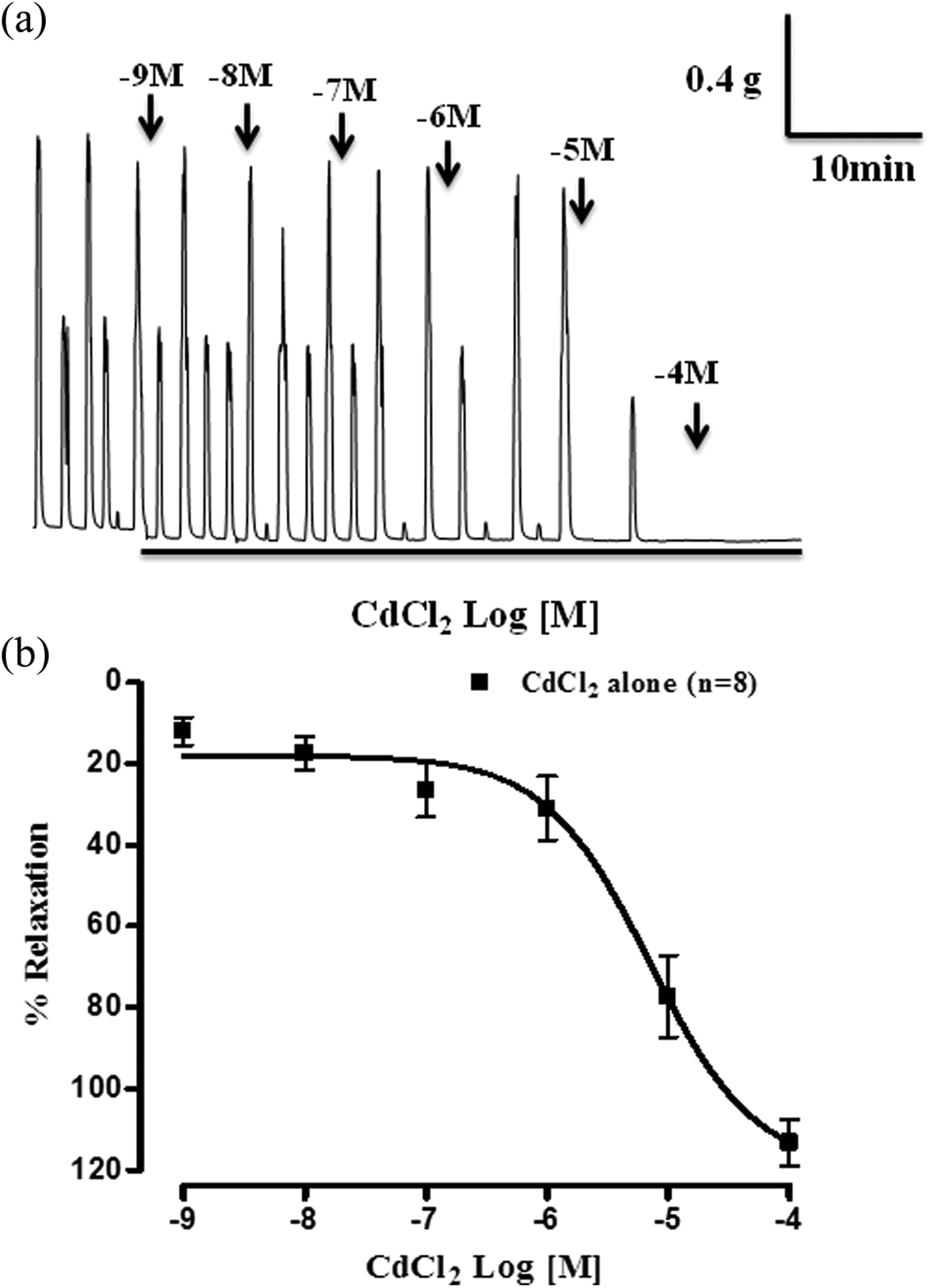
Representative physiograph recording (a) and cumulative concentration response curve (b) of cadmium chloride (1 nM to 0.1 mM) on rat myometrium. Vertical bars represent SEM (n = 8).
Effect of cadmium chloride on CaCl2-induced myometrial contraction in Ca2+-free high K+ (80 mM) depolarizing solution
To assess whether Cd directly reduces calcium entry through L-type of calcium channels, effect of cumulative doses of CaCl2 (1 µM–10 mM) was recorded in Ca2+-free high K+ (80 mM) depolarizing solution alone and in the presence of 10 μM Cd as this dose produced submaximal (∼80%) relaxation in rat myometrium. In this protocol, myometrial strips were initially exposed to Ca2+-free RLS containing 3 mM ethylene glycol tetraacetic acid and after two successive washings with this solution at 4-min interval, tissues were incubated with Ca2+-free high K+ depolarizing solution (to open the L-type Ca2+ channels) before recording the dose response of CaCl2 alone and in the presence of Cd (10 µM). In the presence of CdCl2, the DRC of CaCl2 was significantly (p < 0.05) shifted towards right (Figure 3(a)) with significant (p < 0.05) reduction of potency of CaCl2 (2.47 ± 0.39, n = 7 vs 3.89 ± 0.24, n = 7) without significant decrease in efficacy (0.70 ± 0.05 g, n = 7 vs 0.79 ± 0.10 g, n = 7).

Cumulative dose–response curve of CaCl2 in Ca2+-free high K+ (80 mM) depolarizing solution (a; n = 7) and that of BAY K-8644 (b; n = 6) on rat myometruim in the presence of CdCl2 (10 µM). Vertical bars represent SEM. Data were analyzed by two-way ANOVA followed by Bonferroni post-hoc tests. *p < 0.05. CaCl2: calcium chloride; ANOVA: analysis of variance.
Effect of cadmium on BAY K-8644-induced myometrial contraction
To further strengthen the finding of reduced calcium entry through L-type calcium channel by Cd in rat myometrium, cumulative dose response of BAY K-8644 (0.1 nM–10 µM) alone and in the presence of Cd (10 µM) were recorded. As shown in Figure 3(b), BAY K-8644 produced dose-dependent contractile effect on rat myometrium which was significantly (p < 0.05) attenuated by Cd (−0.30 ± 0.06 g, n = 6) compared to that of BAY K-8644 alone (0.51 ± 0.08 g, n = 6) but without any significant change in potency (6.61 ± 0.29, n = 6 vs 5.62 ± 0.45, n = 6).
Effect of L-NAME on cadmium-induced myometrium relaxation
To investigate the possible involvement of NO in CdCl2 (1 nM–0.1 mM)-induced relaxation, effect of CdCl2 was studied on rat myometrium in the presence of L-NAME. In the presence of L-NAME (0.1 mM), the DRC of CdCl2 was significantly (p < 0.05) shifted toward right (Figure 4(a)) but without any significant decrease in potency and efficacy of CdCl2. The pD2 and Rmax values of CdCl2 alone and in the presence of L-NAME were found to be 5.13 ± 0.24 (n = 7) vs 4.70 ± 0.54 (n = 7) and 116.73 ± 5.23% (n = 7) vs 100.52 ± 6.44% (n = 7), respectively.

Cumulative concentration response curves of cadmium chloride alone and in the presence of 0.1 mM L-NAME (a) and 25 µM ODQ (b) on isolated rat myometrial strips (n = 7). Vertical bars represent SEM. Data were analyzed by two-way ANOVA followed by Bonferroni post-hoc tests. *p < 0.05. L-NAME: Nω-nitro-
Effect of ODQ on cadmium-induced myometrium relaxation
To study the involvement of NOS-NO-sGC pathway in mediating CdCl2-induced myometrial relaxation, cumulative dose response of CdCl2 alone and CdCl2 in the presence of ODQ (25 µM) was recorded. In the presence of ODQ, the DRC of CdCl2 was significantly (p < 0.05) shifted toward right (Figure 4(b)) with significant (p < 0.05) inhibition in efficacy but nonsignificant reduction of potency. The pD2 and Rmax values of CdCl2 alone and in the presence of ODQ were found to be 5.18 ± 0.16 (n = 7) vs 4.66 ± 0.19 (n = 7) and 116.73 ± 5.23% (n = 7) vs 95.69 ± 7.30% (n = 7), respectively.
Effect of glybenclamide on cadmium-induced myometrium relaxation
To elucidate the possible involvement of adenosine triphosphate-sensitive potassium channels (KATP) in CdCl2-induced rat myometrial relaxation, effect of CdCl2 was studied in the presence of glybenclamide (10 µM). Neither the DRC of CdCl2 was shifted either way by glybenclamide as shown in Figure 5 nor was there any significant decrease in potency and efficacy of CdCl2. The pD2 value of CdCl2 alone and in the presence of glybenclamide were almost comparable and found to be 4.99 ± 0.17 (n = 6) vs 4.89 ± 0.19 (n = 6) but Rmax value reduced by almost 15% as the Rmax values were found to be 96.53 ± 3.26% (n = 6) and 113.71 ± 7.15% (n = 6) in the presence and absence of glybenclamide, respectively.
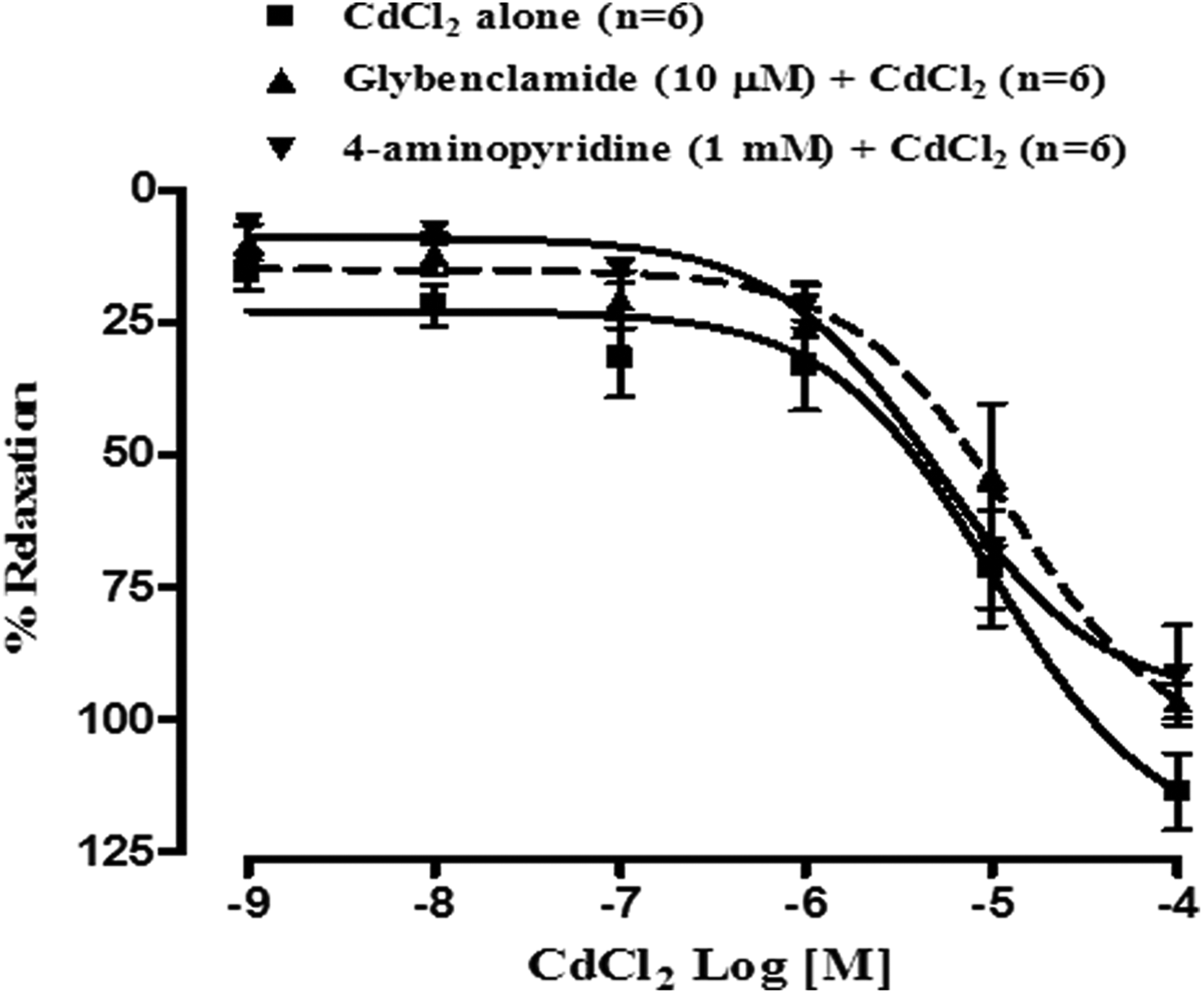
Effect of glybenclamide (10 µM) and 4-aminopyridine (1 mM) on cumulative concentration response curves of cadmium chloride on isolated rat myometrial strips (n = 6). Vertical bars represent SEM. Data were analyzed by two-way ANOVA followed by Bonferroni post-hoc tests. ANOVA: analysis of variance.
Effect of 4-aminopyridine on cadmium-induced myometrium relaxation
To study the possible involvement of voltage-gated potassium channels (Kv) in CdCl2-induced rat myometrial relaxation, effect of CdCl2 alone and in the presence of 4-amonopyridine (1 mM) was studied. In the presence of 4-amonopyridine, the DRC of CdCl2 was nonsignificantly shifted toward right (Figure 5) and there was no significant (p < 0.05) alteration in potency as the pD2 values of CdCl2 alone and in the presence of 4-amonopyridine were found to be 4.99 ± 0.17 (n = 6) vs 5.30 ± 0.17 (n = 6). But Rmax value was found to be reduced by almost 19% as Rmax values were found to be 113.71 ± 7.15% (n = 6) and 91.63 ± 9.46% (n = 6), respectively, in the absence and presence of 4-aminopyridine.
Effect of cadmium on phenylephrine-induced myometrial relaxation
Effect of cumulative-doses of phenylephrine alone (1 nM–10 µM) and in the presence 10 µM CdCl2 was studied in PGF2α (10 µM)-precontracted myometrial strips as this concentration of PGF2α (10 µM) was found to produce maximal tension on rat myometrium. Phenylephrine produced dose-dependent relaxant effect on rat myometrium and the DRC of phenylephrine was significantly (p < 0.05) shifted toward left (Figure 6(a)) in the presence of CdCl2 (10 µM) with significant (p < 0.05) increase in potency and nonsignificant increase in efficacy (Rmax). The pD2 values of phenylephrine were found to be 6.22 ± 0.11 (n = 9) and 6.65 ± 0.17 (n = 9) and Rmax values were 140.02 ± 14.33% (n = 9) and 189.52 ± 22.03% (n = 9), respectively, in the absence and presence of CdCl2 (10 µM).
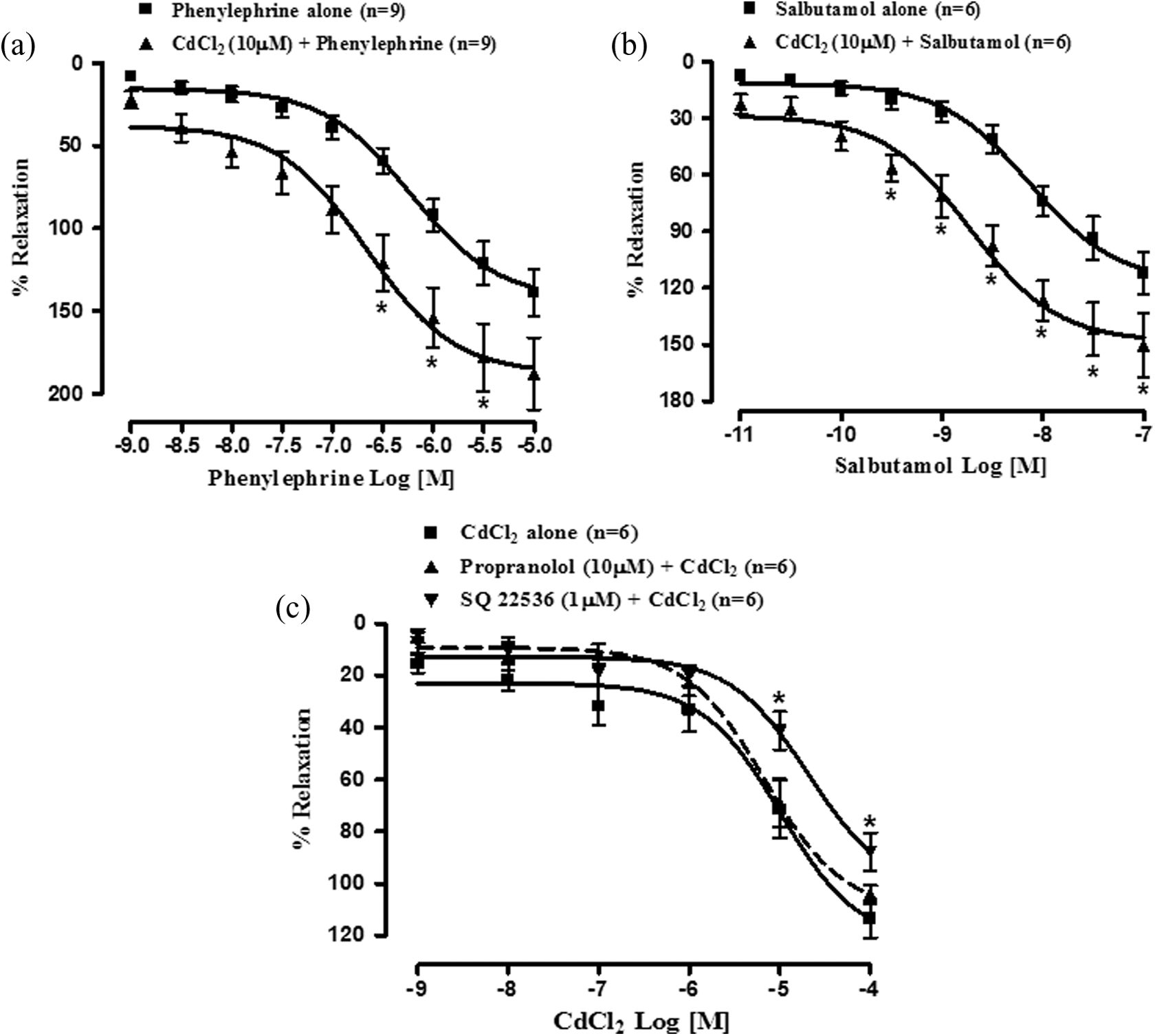
Cumulative concentration response curves of phenylephrine (a, n = 9) and salbutamol (b, n = 6) alone and in the presence of 10 µM CdCl2. (c) depicts the cumulative concentration response curves of CdCl2 alone and in the presence of propranolol (10 µM) or SQ 22536 (1 µM) on isolated rat myometrial strips (n = 6).Vertical bars represent SEM. Data were analyzed by two-way ANOVA followed by Bonferroni post-hoc tests.*p < 0.05. ANOVA: analysis of variance; CdCl2: cadmium chloride.
Effect of cadmium on salbutamol-induced myometrial relaxation
In the presence of CdCl2 in PGF2α (10 µM)-precontracted tissue, salbutamol produced dose-dependent relaxant effect similar to that by phenylephrine and the DRC of salbutamol was significantly (p < 0.05) shifted toward left (Figure 6(b)) with significant (p < 0.05) increase in potency but nonsignificant increase (p < 0.05) in efficacy (Rmax). The pD2 values of salbutamol were found to be 8.13 ± 0.12 (n = 6) and 8.72 ± 0.14 (n = 6), and Rmax values were 112.11 ± 11.25% (n = 6) and 150.36 ± 17.06% (n = 6), respectively, in the absence and presence of CdCl2 (10 µM).
Effect of propranolol on cadmium-induced relaxation of rat myometrium
In order to elucidate possible involvement of adrenergic receptors, especially β-adrenergic receptors, in mediating CdCl2-induced myometrial relaxation, effect of CdCl2 alone and in the presence of propranolol (10 µM) was studied. Although the DRC of CdCl2 was shifted toward right in the presence of propranolol (10 µM) (Figure 6(c)), neither the shift nor the decrease in potency and efficacy of CdCl2 were statistically significant. The pD2 values of CdCl2 alone and in the presence of propranolol were found to be 4.99 ± 0.17 (n = 6) vs 5.16 ± 0.12 (n = 6) while Rmax values were 113.71 ± 7.15% (n = 6) vs 104.23 ± 3.52% (n = 6).
Effect of SQ 22536 on cadmium-induced relaxation of rat myometrium
In view of the significant potentiation of phenylephrine- and salbutamol-induced myometrial relaxation by CdCl2 and failure of propranolol (10 µM) to attenuate CdCl2-induced relaxation, possible involvement of adrenoceptor-independent adenylyl cyclase–cAMP pathway was studied by observing the effect of cadmium alone and in the presence of SQ 22536 (1 µM). In the presence of SQ 22536, the DRC of CdCl2 was significantly (p < 0.05) shifted towards right (Figure 6(c)) with nonsignificant (p < 0.05) decrease in both the potency and efficacy of CdCl2. The pD2 values of CdCl2 were found to be 4.99 ± 0.17 (n = 6) and 4.67 ± 0.18 (n = 6) and Rmax values were 113.71 ± 7.15% (n = 6) and 87.82 ± 7.43% (n = 6), respectively, in the absence and presence of SQ 22536.
Discussion
Studies at cellular level under in vitro conditions have suggested that Cd exhibits multifarious actions but the precise mechanism(s) are yet to be comprehensively explained. 2 Our observation of inhibitory effect of Cd on rat myometrium is in agreement with similar findings on both human and bovine myometrium. 8,9 Cd has been reported to produce dose-dependent differential effects on rat thoracic aorta, that is, stimulatory at lower concentration (1 nM–0.1 µM) and inhibitory at higher concentration (1 µM–0.1 mM), 12 but no such effect was observed on rat myometrium. Thus, possible involvement of different mechanistic pathways in Cd-induced actions on different smooth muscles from different species cannot be ruled out.
Calcium is the major regulatory ion in reactivity of most smooth muscles including myometrium. Results of our study suggest that Cd decreases influx of Ca2+ through L-type of calcium channels by blocking voltage-dependent calcium channel (VDCC) as it significantly (p < 0.05) shifted the DRC of CaCl2 toward right with significant reduction in potency (2.47 ± 0.39) in Ca2+-free high K+ (80 mM) depolarizing solution. Blockade of VDCC is further substantiated by our observation of the significant (p < 0.05) reduction in BAY K-8644-induced myometrial contractions in the presence of submaximal concentration of Cd (10 µM). Similar findings have been reported in human uterine vessels where Cd blocked calcium channel. 22 Cd inhibits Ca2+ availability in rat aorta 12 and produced dose-dependent inhibitory effect on Ca2+-induced contractions in isolated cerebral and peripheral arteries of dog at higher concentration. 23 Cd enters through L-type calcium channel in pituitary cell line (GH4C1) 24 and neurons 25 and acts as a competitive inhibitor of calcium or decreases Ca2+ influx; therefore, similar effect of Cd on rat myometrium cannot be ruled out.
NO is known to be involved in a variety of physiological functions, including neurotransmission, vasodilation, and immunity. 26,27 Several reports indicate that nitric oxide synhase III (eNOS), but not NOS II (iNOS), is expressed in nonpregnant rat uterus, whereas both NOS II and III are expressed in pregnant rat uterus. 28,29 In our study, L-NAME, a NOS inhibitor, significantly inhibited Cd-induced inhibitory effect on myometrium which suggests possible stimulation of NOS by Cd. Significant shift of the DRC of Cd towards right with significant decrease in Rmax value of Cd in the presence of ODQ (25 µM), a specific inhibitor of guanylyl cyclase, added credence to involvement of NOS-NO-sGC pathway in mediating Cd-induced inhibitory effect on rat myometrium, similar to that reported in rat aorta 15 with enhanced expression of eNOS and iNOS in Cd-treated rats. But on the contrary, Cd has been reported to inhibit NO production in endothelial cells by blocking eNOS phosphorylation 30 and increase vasoconstrictor activity of phenylephrine by reducing NO bioavailability by increasing oxidative stress following Cd exposure. 31
To date, several types of K+ channels have been identified in myometrium which include ATP-sensitive K+ channel (KATP), shaker-like voltage-gated potassium channels (KV), calcium-activated large conductance potassium channels (BKCa), and small conductance calcium-sensitive potassium channels (SKCa). 32 Voltage-gated K+ (Kv) channels control excitability in nonpregnant and pregnant rat myometrium. 21 In the present study, 4-aminopyridine (1 mM), a specific blocker of Kv channel, 33 and glybenclamide (10 µM), a selective KATP channel blocker, 34 failed to attenuate inhibitory effect of Cd on rat myometrium, thus suggesting that possibly activation of voltage-dependent and ATP-dependent K+ channels do not have any role in CdCl2-induced myometrial relaxation. Our findings are in confirmation with the observations in bovine mesenteric artery where Cd has been reported not to affect the Ca2+-insensitive K+ channels. 35
Isolated rat uterus throughout natural estrous cycle possesses predominantly beta-adrenergic receptors. 36 High-density α1 and β2 adrenergic receptors are expressed in the circular and longitudinal layers of rat myometrium, respectively. 37 Phenylephrine and salbutamol, respectively selective α1 and β2 adrenoceptor agonists, produced concentration-dependent relaxant effect on rat myometrium. Mhaouty-Kodja et al. 38 have also reported relaxant effect of phenylephrine on myometrial strips of late pregnant and parturient mouse due to stimulation of β-adrenoceptors at higher concentration of phenylephrine. Significant leftward shift of the DRCs of phenylephrine and salbutamol in the presence of CdCl2 with significant increase in potency (pD2) without change in efficacy (Rmax) suggests that CdCl2 possibly enhances relaxant effect of β-adrenoceptor stimulants by affecting some downstream signaling mechanism rather than adrenergic receptors. This hypothesis is supported by our observation that SQ 22536, an inhibitor of adenylyl cyclase, significantly attenuated the CdCl2-induced tocolytic effect on rat myometrium, whereas propranolol, an antagonist of β- adrenoceptors, failed to produce any significant inhibitory effect on CdCl2-induced myometrial relaxation.
Summing up our findings, it may be inferred that Cd-produced relaxant effect on rat myometrium and its direct effect and potentiation of adrenoceptors agonists-induced relaxation in rat myometrium is mediated through blockade of calcium entry through L-type calcium channel and/or stimulation of NOS-NO-sGC and AC-cAMP pathways.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Indian Council of Agricultural Research (ICAR), New Delhi, India under Niche Area of Excellence programme to Department of Veterinary Pharmacology and Toxicology, DUVASU, Mathura, India (Grant No.:10(10)/2012-EPD, dated: 23rd March, 2012).