
Abstract
Background and objectives:
The aim of this study was to evaluate the protective effects of quercetin on cisplatin-induced hair cell damage in transgenic zebrafish embryos.
Materials and methods:
Five days postfertilization zebrafish embryos were exposed to 1 mM cisplatin and quercetin at 10, 50, 100, or 200 μM for 4 h. Hair cells within neuromasts of the supraorbital, otic, and occipital lateral lines were analyzed by fluorescent microscopy (n = 10). Survival of hair cells was calculated as the average number of hair cells in the control group that were not exposed to cisplatin. Ultrastructural changes were evaluated using scanning electron microscopy.
Results:
Hair cell damage in neuromasts was decreased by co-treatment of quercetin and cisplatin (quercetin 100 μM: 8.6 ± 1.1 cells; 1 mM cisplatin only: 5.0 ± 0.5 cells; n = 10, p < 0.05); apoptosis of hair cells examined by special stain was also decreased by quercetin. The ultrastructure of hair cells within neuromasts was preserved in zebrafish by the combination of quercetin (100 μM) and cisplatin (1 mM).
Conclusion:
In conclusion, quercetin showed protective effects against cisplatin-induced toxicity in a zebrafish model. The results of this study suggest the possibility of a protective role of quercetin against cisplatin-induced apoptotic cell death in zebrafish.
Introduction
Cisplatin is a widely used chemotherapeutic agent for the treatment of various soft tissue malignant neoplasms. The platinum atom of cisplatin forms covalent bonds with DNA at the N7 positions of purine bases to form intrastrand and interstrand DNA cross-links in tumor cells. 1 Cell death induced by cisplatin occurs primarily through apoptosis. 2 Although cisplatin is an effective and useful chemotherapeutic agent, its use is limited by well-known side effects of nephrotoxicity, neurotoxicity, and ototoxicity. 1,3 The ototoxic manifestations of cisplatin include tinnitus and sensorineural hearing loss. Hearing loss induced by cisplatin appears to be dose dependent, cumulative, bilateral, occurs initially at higher frequencies, and is usually permanent. 4 Cisplatin ototoxicity appears to involve damage of not only hair cells, especially outer hair cells, but also degeneration of the stria vascularis, spiral ganglion cells, and supporting cells in the cochlea. 5–8
Cisplatin-induced ototoxicity is related to reactive oxygen species (ROS), including superoxide anions. ROS can deplete cochlear tissues of antioxidant protective molecules, including glutathione (GSH) and antioxidant enzymes. Free radical production with resulting GSH depletion and subsequent lipid peroxidation is a proposed pathway for cisplatin ototoxicity. 1 Specifically, free radical increases can increase calcium influx and induce apoptosis in cells of the cochlea. 1
Flavonoids are phytophenolic compounds with strong antioxidant effects that function as free radical scavengers. Flavonoids are present in fruits, vegetables, nuts, and plant-derived beverages, such as tea and wine. In addition, numerous antioxidant supplements and herb-containing medicaments contain high doses of flavonoids. Quercetin (3,5,7,3′,4′-pentahydroxy flavone) 9 is an important member of the flavonoid family in the human diet and is commonly present in most edible fruits and vegetables. 9–11 Quercetin has been reported to inhibit thrombocyte aggregation, to have an antihypertensive effect, and to reduce both oxidative stress in streptozotocin-induced diabetic rats and cisplatin-induced nephrotoxicity. 11–14 However, the protective effects of quercetin against cisplatin-induced ototoxicity have not yet been investigated.
Recently, zebrafish have been used to study ototoxicity. Zebrafish have several benefits. First, zebrafish have clusters of 5–10 hair cells, which are similar to the structure of hair cells in mammals. Second, the location of hair cells of zebrafish is superficial on the surfaces of their bodies. Therefore, the assessment of hair cells is very simple and rapid. Third, zebrafish are easily available for repeated and multiple tests because of high fecundity. The transgenic zebrafish (Brn3C:EGFP: enhanced green fluorescent protein) have distinguishing hair cells, which are visible under a fluorescent microscope without staining. 15–18
Hair cells of neuromasts in the lateral line of transgenic zebrafish (Brn3C: EGFP) have been demonstrated to be a useful system for studying hair cell loss after exposure to ototoxic agents as well as for studying the protective effect of antiototoxic agents. 18–20 The purpose of this study was to investigate the protective effects of quercetin on cisplatin-induced hair cell damage in transgenic zebrafish embryos.
Materials and methods
Chemicals and zebrafish housing
Cisplatin was purchased from Sigma Chemical Co. (St Louis, Missouri, USA). Quercetin was purchased from Cayman Chemical Company (Ann Arbor, Michigan, USA). Transgenic zebrafish (Brn3C: EGFP) embryos were provided by paired matings of adult fish maintained at 28.5 ± 1°C in a zebrafish facility at the Korea University Ansan Hospital, Ansan, Republic of Korea. Embryos were maintained in embryo medium (15 mM sodium chloride, 0.5 mM potassium chloride, 1 mM calcium chloride, 1 mM magnesium sulfate, 0.15 mM monopotassium phosphate, 0.05 mM ammonium phosphate, and 0.7 mM sodium bicarbonate) 21 and staged according to days postfertilization (dpf) and morphological criteria. 22 This study protocol was approved by the Korea University Institutional Animal Care and Use Committee (approval no. KUIACUC-2012-116). All experiments were performed in accordance with the guidelines of the Animal Care Ethics Committee of Korea University Medical Center and National Institutes of Health guidelines.
Drug treatment and evaluation of hair cells of zebrafish
Cisplatin solutions were prepared by adding the cisplatin powder to embryo medium. Cisplatin decreased the viability of neuromasts in a dose-dependent manner. We previously reported that neuromasts treated with 1 mM cisplatin for 4 h exhibited a viability of 50%, 23 and thus a dose of 1 mM cisplatin was chosen as an adequate experimental concentration for this study. Five dpf zebrafish were simultaneously exposed to 1 mM cisplatin and quercetin at concentrations of 10, 50, 100, or 200 μM for 4 h. After they were rinsed three times in embryo media, zebrafish were anesthetized with Tricane (3-aminobenzoic acid, 0.4 g/ethyl ester, 100 mL, pH 7, adjustments by Tris buffer) for 5 min. It has been known that zebrafish have a rapid regeneration activity. 3 However, we identified that regeneration of hair cells of zebrafish did not show significant within 2 h after combination exposure in this study (data not shown). Therefore, all experiments were performed within 2 h after chemicals exposure because hair cells of zebrafish were less influenced by regeneration itself. The zebrafish embryos were then mounted with methylcellulose on a depression slide for evaluation under a fluorescence microscope. The survival hair cells within the neuromasts of the supraorbital (SO1 and SO2), otic (O1), and occipital (OC1) lateral lines were determined using a fluorescence microscope. 3,15,23,24 The average numbers of hair cells of the SO1, SO2, O1, and OC1 neuromasts on one side of each fish were investigated in each zebrafish for all experimental and control conditions (n = 10) with a fluorescence microscope (AxioCam MRc5; Carl Zeiss, Jena, Germany). Previous study has determined that there is no significant differential sensitivity between neuromasts. 3,25 The total number of hair cells was counted by adding the number of hair cells from the four neuromasts. The average number of hair cells was calculated by dividing the total number of hair cells of each fish in the experimental group by the total number of fish in a similarly treated control group.
Examination of mitochondrial damage and apoptosis of hair cell of zebrafish
The fluorescent dye 2-[4-(dimethylamino) styryl]-N-ethylpyridinium iodide (DASPEI; Invitrogen, Carlsbad, California, USA) was applied to evaluate mitochondria within hair cells. The zebrafish embryos were exposed to medium containing 1 mM cisplatin and 100 μM quercetin for 4 h. The embryos were then washed with phosphate-buffered saline (PBS) and fixed in 4% paraformaldehyde. Next, the embryos were incubated in embryo medium containing 0.005% DASPEI for 15 min and analyzed as described above. 25
Apoptosis in neuromasts was estimated by the terminal deoxynucleotidyl transferase (TdT)-mediated 2′-deoxyuridine 5′-triphosphate (dUTP) biotin nick end-labeling (TUNEL) assay using an in situ cell detection kit (Roche Molecular Biochemicals, Mannheim, Germany) according to the manufacturer’s protocol. The embryos were then washed with PBS and fixed in 4% paraformaldehyde. The embryos were then incubated with 50 μL of the TUNEL reaction mixture (TdT and fluorescein-dUTP) at 37°C for 1 h in a humid atmosphere, after which they were evaluated under a fluorescence microscope. 23
Ultrastructural analysis by SEM
For scanning electron microscopy (SEM), five dpf zebrafish embryos exposed to 1 mM cisplatin and 100 μM quercetin for 4 h were prefixed by immersion in 2% glutaraldehyde in 0.1 M phosphate buffer and postfixed for 2 h in 1% osmic acid dissolved in PBS. The zebrafish embryos were treated in a graded series of ethanol and t-butyl alcohol solutions, freeze-dried (ES-2030; Hitachi, Tokyo, Japan), coated with platinum using an ion coater (IB-5; Eiko, Katsuda, Japan), and evaluated by a field-emission SEM (S-4700; Hitachi). 23
Statistical analysis
All values are presented as mean ± SD. For statistical comparisons, one-way analysis of variance was used for multiple comparisons. A post hoc analysis was performed using Tukey’s honestly significant difference test. The values of p < 0.05 were considered statistically significant. Statistical analysis was performed with Statistical Package for the Social Sciences Version 20.0 (IBM, Aramark, New York, USA).
Results
Quantitative analysis of hair cells in transgenic zebrafish embryos after treatment with cisplatin and quercetin
Hair cells from the four neuromasts (SO1, SO2, O2, and OC1) were evaluated by fluorescent microscopy to investigate the changes induced after treatment with cisplatin and quercetin. While 1 mM cisplatin significantly decreased, the numbers of hair cells in neuromasts, quercetin-treated zebrafish exhibited a significantly increased average number of hair cells (control: 13.1 ± 0.9 cells, 1 mM cisplatin only: 5.0 ± 0.5 cells, quercetin 10 μM: 5.3 ± 0.7 cells, 50 μM: 6.1 ± 0.6 cells, 100 μM: 8.6 ± 1.1 cells, 200 μM: 4.8 ± 0.4 cells; n = 10, p < 0.05; Figures 1 and 2).
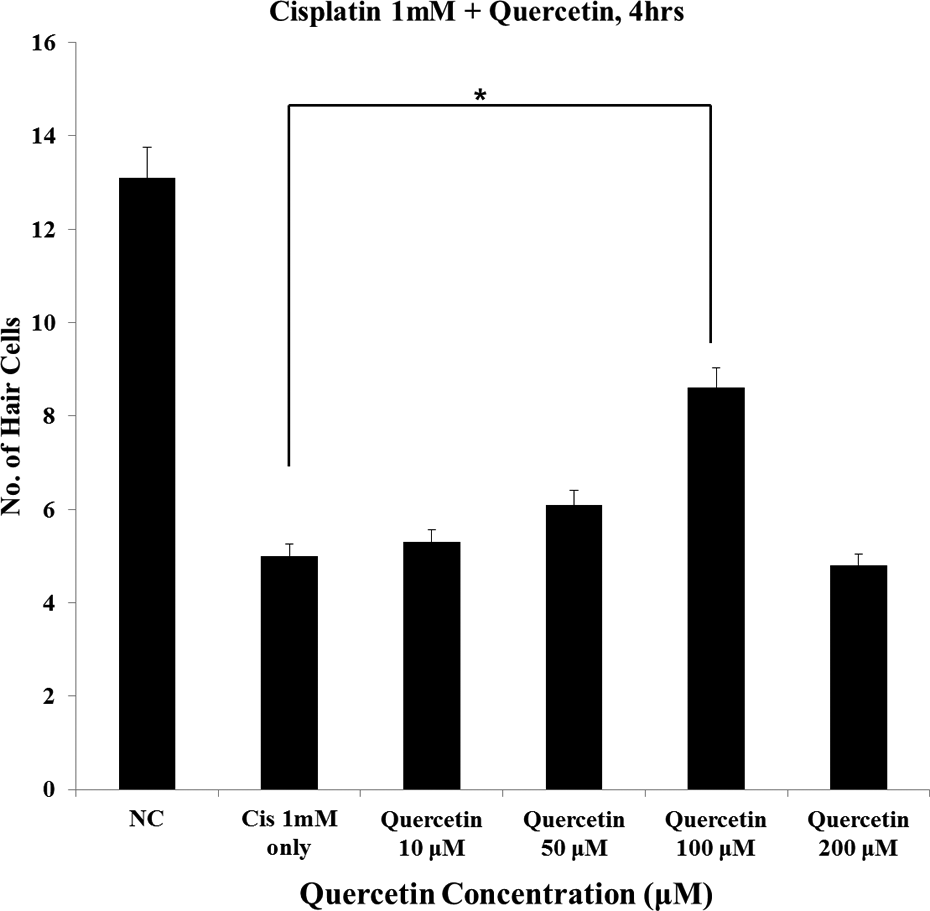
Quantitative analysis of hair cells within neuromasts of transgenic zebrafish embryos. Hair cells from four neuromasts (SO1, SO2, O1, and OC1) were counted and viable hair cells were calculated as average number of hair cells in the control group. The embryos co-treated with 100 μM quercetin and cisplatin exhibited most significantly increased levels of fluorescence compared with zebrafish treated with cisplatin only (control: 13.1 ± 0.9 cells, 1 mM cisplatin only: 5.0 ± 0.5 cells, quercetin 10 μM: 5.3 ± 0.7 cells, 50 μM: 6.1 ± 0.6 cells, 100 μM: 8.6 ± 1.1 cells, 200 μM: 4.8 ± 0.4 cells; n = 10, *p < 0.05). NC: negative control; Cis: cisplatin; SO: supraorbital; O: otic; OC: occipital.

Fluorescent microscopic findings (×40) of hair cells in neuromasts. Treatment with 1 mM cisplatin resulted in a significant decrease in the fluorescence of neuromasts. Zebrafish treated with quercetin showed higher fluorescence compared with zebrafish treated with cisplatin alone. Zebrafish treated with 100 μM quercetin exhibited the highest fluorescence. Bar = 10 µm. NC: negative control; Cis: cisplatin.
Evaluation of the effects of quercetin on mitochondrial damage and apoptosis in hair cells of transgenic zebrafish embryos
Treatment with 100 μM quercetin resulted in the largest number of preserved hair cells under cisplatin-induced toxicity (data as above), and thus this concentration was used for DASPEI and TUNEL assays. In cisplatin-treated zebrafish, fluorescence after DASPEI stain was decreased compared with the negative control. In addition, zebrafish co-treated with 100 μM quercetin and cisplatin exhibited increased levels of fluorescence compared with zebrafish treated with cisplatin alone (Figure 3). Apoptotic cells were marked as light red dots by the TUNEL assay, and comparison of the color intensity between the cisplatin-treated zebrafish and the quercetin co-treated zebrafish under a fluorescent microscope revealed that quercetin co-treated zebrafish exhibited a significantly decreased number of TUNEL-positive cells (Figure 4).

Evaluation of mitochondrial damage with DASPEI stain (O1, ×40). Co-treatment with 100 μM quercetin resulted in a significant decrease in mitochondrial staining with DASPEI compared with zebrafish treated with cisplatin alone. Bar = 10 µm. O: otic; NC: negative control; Cis: cisplatin; DASPEI: 2-[4-(dimethylamino) styryl]-N-ethylpyridinium iodide.
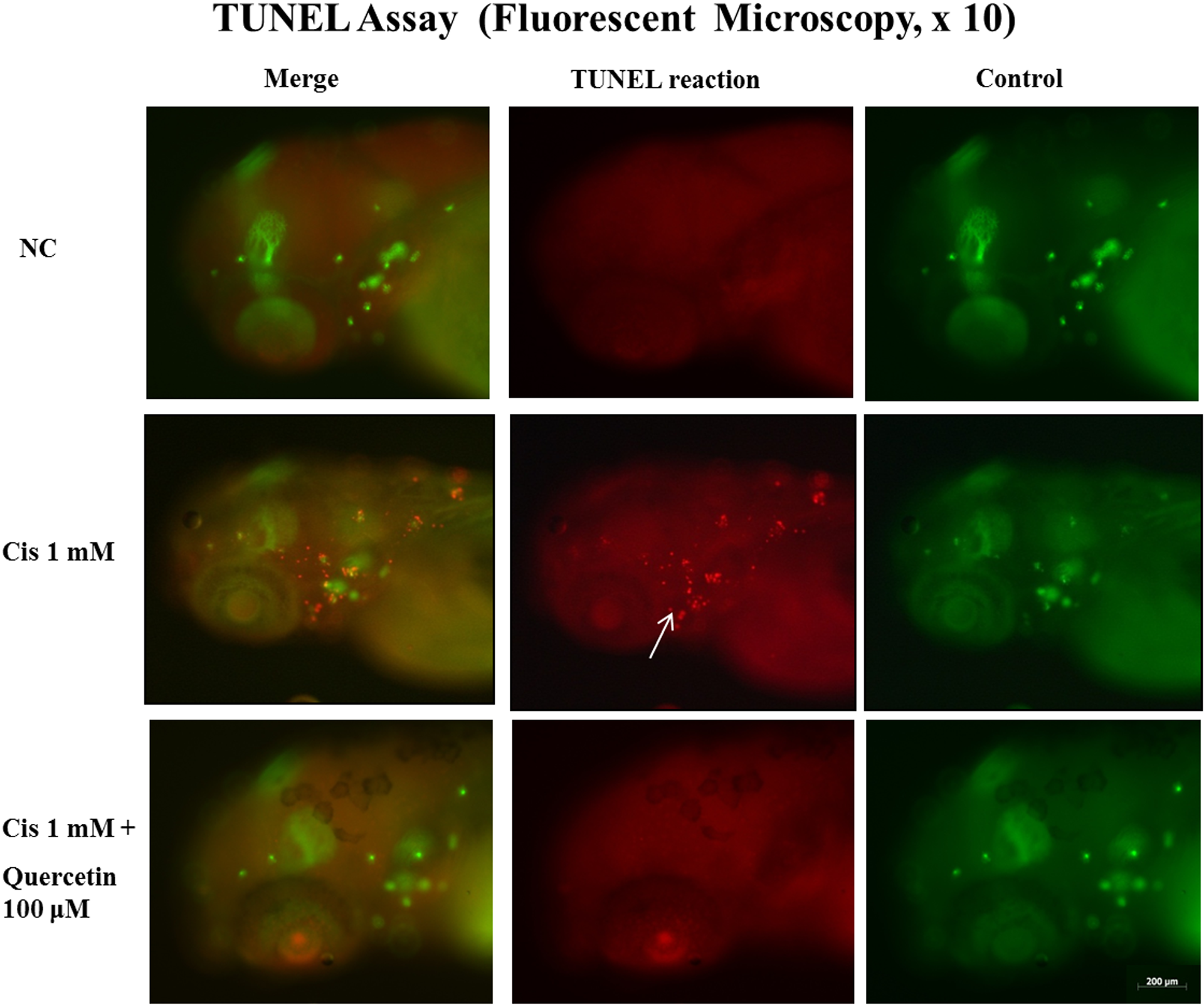
Evaluation of apoptosis with TUNEL assay. Apoptotic cells were marked as light red dots in red-colored fish after TUNEL staining by fluorescent microscopy (arrow indicates TUNEL-positive cells). A comparison of the color intensity between the groups treated with 1 mM cisplatin with 100 μM quercetin for 4 h showed that quercetin significantly decreased the number of TUNEL-positive cells (green-colored fish of right column: control for TUNEL reaction; red-colored fish of middle column: TUNEL reaction; green- and red-colored fish of left column: merge for control and TUNEL reaction). TUNEL Bar = 200 µm. TUNEL: terminal deoxynucleotidyl transferase-mediated 2′-deoxyuridine 5′-triphosphate biotin nick end-labeling.
Protective effect of quercetin on the ultrastructure of transgenic zebrafish treated with cisplatin
The kinocilia and stereocilia of zebrafish embryos were evaluated using SEM analysis. Compared with the control, exposure to 1 mM cisplatin for 4 h resulted in severe morphological changes of hair cells including loss or fusion of stereocilia and kinocilia. Conversely, zebrafish treated with 100 μM quercetin showed minimal changes of the stereocilia and kinocilia compared with the control (Figure 5).

SEM (SO1, ×3k) findings of hair cells in neuromasts. Compared with control zebrafish, the neuromasts in zebrafish treated with 1 mM cisplatin for 4 h exhibited severe morphological change to kinocilia. This type of damage was decreased in zebrafish co-treated with quercetin. Bar (at the bottom of each figure, one space) = 10 µm. SEM: scanning electron microscopy; SO: supraorbital.
Discussion
Ototoxic injury to hair cells is a well-known side effect of certain chemotherapeutic agents, especially cisplatin. However, cisplatin and other platinum derivatives are common components of many chemotherapeutic regimens, with no true alternatives available. Indeed, while it is known to be effective against a variety of solid malignancies, the use of cisplatin is still limited by its well-known ototoxicity and nephrotoxicity.
Cisplatin generates ROS, including superoxide anions in explants of cochlear tissues. 26 Interestingly, ROS mimics the effects of cisplatin on outer hair cells in vitro. Specifically, ROS can deplete cochlear tissues of antioxidant protective molecules, including GSH and antioxidant enzymes, which may allow lipid peroxidation, as evidenced by increased concentrations of malondialdehyde and toxic lipid peroxides and aldehydes, such as 4-hydroxynonenal and peroxynitrite. 1,27 In turn, these agents can increase calcium influx and apoptosis in cells of the cochlea. 26–28 ROS also activate Bcl-2-associated X protein in the cytosol, which translocates to mitochondria, causing mitochondrial damage and release of cytochrome c into the cytosol, 1 which in turn can activate caspases 9 and 3, leading to a breakdown of DNA by caspase-activated deoxyribonuclease and cleavage of fodrin within the cuticular plate of damaged outer hair cells by caspase 3, leading to apoptosis. 29,30 Cells exposed to cisplatin exhibit increased phosphorylation of extracellular regulated kinase 1/2, which can function as a main effector of cell death, causing both nuclear fragmentation and actin cytoskeleton rearrangement. 1
Zebrafish have hair cells on the surface of their bodies as part of a lateral line sensory system. These hair cells are structurally and functionally similar to mammalian inner hair cells and are organized into clusters of 5–20 hair cells called neuromasts. 3 The hair cells of neuromasts selectively take up fluorescent vital dyes such as YO-PRO1 and FM1-43, and this selective uptake combined with the optical clarity of the body allows for rapid in vivo assessment of hair cell death. 3 Ototoxic and potentially protective compounds can be added directly to zebrafish water, allowing direct observation of hair cell death as well as construction of precise dose–response curves. 3
A recent study established that cisplatin causes repeatable and predictable damage to zebrafish lateral line hair cells, defined the time course of hair cell loss, and demonstrated that the kinetics of hair cell loss in response to treatment with cisplatin at both low and high doses follow a mathematically predictable relationship. 3 This study provided additional support for the validity and utility of using the zebrafish lateral line as an accessible model for studying mechanosensory hair cell death from various ototoxic agents as well as the possibility of evaluating the effects of anti-ototoxic agents on lateral hair cells of zebrafish. 3 Indeed, several agents applied to hair cells of the lateral line of zebrafish in combination with ototoxic agents have been reported to have protective effects. 23
Flavonoids are a class of naturally occurring polyphenolic compounds, ubiquitously present in photosynthesizing cells. 31 Most flavonoids share a common three-ring structure, of which rings A and B are aromatic and ring C is heterocyclic. Variation in the heterocyclic C ring forms the basis of the division of the flavonoids into various subclasses including flavones, isoflavone, flavonols, flavanals, flavanones, anthocyanidins, and chalcones. 32 Quercetin is a dietary flavonol that is by far the most abundant of the flavonoids present in edible plants and beverages. 33 Various epidemiological studies have been performed to provide support for the proposed beneficial health effects of a high intake of flavonoids such as quercetin. 31 A protective effect of flavonol and flavone intake has been found with respect to (1) risk of fatal or nonfatal coronary artery diseases, 34 (2) risk of lung cancer, 35 (3) incidence of asthma, 35 and (4) impairment of pulmonary function in chronic obstructive pulmonary disease. 36 The beneficial effects of flavonoids are attributed primarily to their antioxidant effects as free radical scavengers, hydrogen-donating compounds, singlet oxygen quenchers, and metal ion chelators. 37
Quercetin has been shown to have excellent antioxidant properties in vitro. Within the flavonoid family, quercetin is the most potent scavenger of ROS, including superoxide
To determine whether cisplatin-induced damage of the lateral line hair cells was averted by quercetin, transgenic zebrafish embryos were treated with various concentrations of quercetin along with 1 mM cisplatin for 4 h. While 1 mM cisplatin alone significantly decreased the numbers of neuromast hair cells, quercetin protected against cisplatin-induced hair cell damage in transgenic zebrafish, with a dose of 100 μM preserving the largest number of hair cells in the neuromast (Figures 1 and 2). Thus, a dose of quercetin at 100 µM was determined to be the most effective dose.
Mitochondrial damage has an important role in ototoxicity. Release of cytochrome P450 from the mitochondria to cytoplasm activates caspases 9 and 3, which in turn activate caspase-activated deoxyribonucleases, resulting in DNA fragmentation and cell death. 1 Mitochondrial damage was investigated with DASPEI staining, which revealed that mitochondria within hair cells were protected by quercetin treatment (Figure 3). In addition, apoptosis in neuromasts was evaluated by TUNEL assay (Figure 4). Quercetin treatment decreased the number of TUNEL-positive cells in neuromasts, suggesting that apoptotic cell death induced by cisplatin was prevented by treatment with quercetin in zebrafish.
Cisplatin ototoxicity affects especially hair cells in humans and induces severe morphological changes in hair cells; clubbing of the distal end of cilia as well as fusion and/or loss of stereocilia or kinocilia are typical changes associated with cisplatin-induced ototoxicity. According to SEM analysis, cisplatin-treated zebrafish exhibited severe morphological change of hair cells, whereas zebrafish co-treated with quercetin exhibited minimal changes of the stereocilia (Figure 5). Thus, these results suggested that quercetin protects against cisplatin-induced hair cell damage compared with treatment with cisplatin alone.
Quercetin exerts antioxidant effect by reducing ROS and increasing endogenous antioxidants. Indeed, the numerous proposed beneficial effects of quercetin are primarily associated with its antioxidant effects. Quercetin is present in fruits, vegetable, and beverages as well as a dietary supplement. Thus, it is very easy to take quercetin compared with other medicines. Two studies have been conducted to evaluate the in vivo effect of quercetin in patients suffering from a hypertension and sarcoidosis. 40,41 Together with the results of this study, such in vivo investigations of quercetin may be helpful for reducing the time and cost for validation of its use in humans.
Despite its advantages, use of quercetin has some limitations. Specifically, quercetin does not produce a definite increase in antioxidant levels compared with normal basal damage level; the beneficial health effect of quercetin is primarily expected to be useful when basal levels of damage are increased. 31 Quercetin has also been reported to have genotoxic effects in vitro through the formation of quinone in bacteria. 42 In addition, the toxicity of long-term use of quercetin has not been established. An additional complication is that it may be difficult to reach high serum concentrations through typical diet or dietary supplement. However, these limitations can be overcome by adopting strategies such as short-term use of quercetin during treatment of cisplatin for a protective purpose, for example, giving intratympanic injections to reach high concentration in the cochlea could be a possible solution to limit quercetin use. Nevertheless, more research will be required to evaluate the potential genotoxicity and other side effects of quercetin in the cochlea.
Conclusion
To the best of our knowledge, this is the first study to investigate the protective effects of flavonoids, specifically quercetin, against cisplatin-induced toxicity in zebrafish, which is an excellent experimental model for ototoxicity. In this study, we showed that concomitant administration of quercetin attenuated cisplatin-induced cell death, mitochondrial damages, apoptosis of hair cells, and ultrastructural changes in hair cells of zebrafish. However, limitations of this study included the lack of specific information about measuring biomarkers such as ROS and GSH levels in the transgenic zebrafish. Functional effects and genotoxicity of quercetin in vivo, especially in mammalian cells, should be analyzed to establish the role of quercetin in cisplatin-induced ototoxicity.
Footnotes
Conflict of interest
The authors declared no conflicts of interest.
Funding
This research was supported by a Korea University Grant, the Soo ENT Clinic, and the Communication Disorders Center of Korea University.