
Abstract
The wide use and wide-spectrum toxicity of synthetic pyrethroid (SP) insecticides make them an emerging ecotoxicological concern. The objective of the current study was aimed to investigate the involvement of oxidative stress in lambda-cyhalothrin (LTC)–induced cerebellum damages in adult rats and to evaluate the possible protective effect of vitamin C (vit C) as antioxidant. Exposure of rats to LTC during 3 weeks caused a significant (p < 0.05) increase in the levels of lipid peroxidation (LPO), nitric oxide (NO) and protein carbonyls (PCO) along with a significant (p < 0.05) decrease in the levels of reduced glutathione (GSH) and the activities of acetylcholinesterase, superoxide dismutase, catalase, glutathione peroxidase and gluthione-S-transferase (p < 0.05) when compared with the control group. The oral administration of vit C (200 mg/kg per d) to LTC-treated rats significantly (p < 0.05) diminished the levels of LPO, NO and PCO and significantly (p < 0.05) increased the activities of GSH and antioxidant enzymes. Our results showed that the administration of vit C could ameliorate some of the oxidative damage in the cerebellum induced by SPs exposure, suggesting that the ascorbic acid could exhibit a potential antioxidant activity against neurotoxicity induced by pesticides exposure.
Introduction
Pyrethroid insecticides represent an increasing proportion of pesticide sales in the world. 1 Pyrethroids’ popularity stems from their insecticidal potency, slow development of pest resistance and relatively low acute toxicity of most compounds in mammals. 2 Lambda-cyhalothrin (LTC), a synthetic pyrethroid (SP) insecticide, has been extensively used in the last two decades in many of the developing countries, including Tunisia, for combating agricultural pests and insects of veterinary as well as human concern. The SP insecticides are widely applied than organochlorines, organophosphates and carbamates, 3 in view of the fact that they have shown to possess a high insecticidal activity as well as a broad spectrum of high initial toxic action on several types of pests. Several studies focused on vertebrate toxicity, including cytotoxicity, 4 endocrine disruption, 5,6 genotoxicity 7,4 and nephrohepatotoxicity. 8,9 Although clark et al. 10 reported that the LTC is a neuropoison interfering with the ionic conductance of nerve membranes by prolonging the sodium current and increasing the spontaneous release of neurotransmitters such as GABA, dopamine or noradrenaline, the involvement of oxidative stress in LTC-induced cerebellum damages in adult rat has not been experimentally documented. The nervous system is especially vulnerable to toxic insults, which can perturb developmental processes like neuronal proliferation, migration and differentiation, synapse formation and myelination, leading to a permanent alteration of brain functions and pathologies later in life. 11 The cerebellum, which controls posture and voluntary movement and mediates motor functions like balance and coordination, is a known target of several toxic compounds. 12 Furthermore, it is highly vulnerable to lipid peroxidation (LPO) because of its high rate of oxygen utilization, an abundant supply of polyunsaturated fatty acids, a deficient antioxidant defense and a high content of transition metals like copper and iron in several regions. 13 Major types of neurotoxicity that can be produced by exposure to xenobiotics are the acute cholinergic syndrome. An accumulation of acetylcholine occurs at the nerve synapse, interfering with the normal nervous system when cholinesterase is inhibited. 14 This produces rapid twitching of voluntary muscles followed by paralysis. The main function of acetylcholinesterase (AChE) is the rapid hydrolysis of acetylcholine at cholinergic synapses. 15
Many insecticides are hydrophobic molecules that bind extensively to biological membranes, especially to the phospholipid bilayers. 16 Numerous studies showed that the antioxidant substances protect cells against deleterious effects of several environmental agents. 17 Vitamins, as antioxidants, may preserve cells membrane function including ion transport and membrane fluidity; they may also prevent the release of Fe2+ and Mg2+ from their binding protein, potentially decreasing the rate of LPO and protein oxidation. 18 The molecular and cellular effects of vitamin C (vit C) have been explained either by acting as an antioxidant preventing damage to membranes or proteins due to an antioxidant activity or by interacting and regulating specific proteins (receptors and transcription factors) capable of regulating signal transduction and gene expression. 19,20 Vit C may effectively minimize free radicals in biological systems, and it is considered the first line of defense against LPO in aqueous system, 21 protecting the cell from an early stage of free radical attack. Therefore, the present study was conducted to evaluate the protective effects of vit C with regard to its antioxidative potential in restoring the altered biochemical enzyme activities, LPO, nitrite formation and protein oxidation in the cerebellum of male rats exposed to SP (LTC), which may have implications in managing humans with accidental exposures to such compounds.
Material and methods
Chemicals
LTC is an SP insecticide (C23H19. ClF3NO3). CAS chemical name is a-cyano-3-phenoxybenzyl-3-(2-chloro-3,3,3-trifluoro-1-propenyl)-2,2-dimethylcyclo-propanecarboxylate, CAS registry number 91465-08-6. A commercial formulation of LTC, named ‘‘KARATE 5EC’’ (Syngenta agrochemicals, Greensboro, North Carolina, USA) was used in the experiments. All other chemical products used in this study were purchased from Sigma Chemical Co. (St Louis, France).
Animals
Male Wistar rats (aged 8–9 weeks; weighing 140–150 g) were obtained from the Central Pharmacy (SIPHAT, Tunis, Tunisia). The animals were housed at 22 ± 3°C temperature at 45 ± 5% humidity, in 12 h-light–dark cycle and left to acclimatize for 1 week before the experiments. They were fed standard laboratory chow 22 and provided with water ad libitum. The experimental protocol was approved by the Local Animal Care Committee, and all the experimental procedures were carried out in accordance with the international guidelines for the Care and use of laboratory animals.
Experimental procedure
Male Wistar rats (n = 28) were randomly divided into four groups of seven animals each. The first group served as the control, received distilled water ad libitum and 1 ml of saline solution was given intraperitoneally (IP). The second group (vit C group) was given a daily single dose of vit C in saline IP (200 mg/kg bw), while the third group (LTC group) received 668 ppm of LTC through drinking water. Animals in the fourth group (LTC + vit C) were given a single injection of vit C IP (200 mg/kg bw per d), 12 h after the administration of LTC. The dose of LTC used in this study represented 1/10 of the median lethal dose (612 mg/kg bw). This dose was used by previous investigations since it is toxic but not lethal to rats. 4,23 This dose of vit C (200 mg/ kg bw per d) gave good protection against neurotoxicity. 24,25 Lower doses of vit C gave less protection, while higher doses were not much more effective.
At the end of the experimental period, the animals in different groups were sacrificed by cervical decapitation to avoid stress conditions. Blood was collected into EDTA tubes and centrifuged (3000g for 15 min) for the separation of plasma. The cerebellum was dissected out, weighed and washed using saline solution. The tissue was minced and homogenized (10% w/v) in phosphate buffer (100 mM Na2HPO4/NaH2PO4; pH 7.4) and centrifuged at 10,000g for 15 min at 4°C. The resulting supernatant was used for various biochemical assays.
Biochemical assays
LPO assays
LPO in the renal tissue was estimated colorimetrically by measuring thiobarbituric acid reactive substances, which were expressed in terms of malondialdehyde (MDA) content, according to Draper and Hadley method. 26 The absorbance was measured at 532 nm. The MDA values were calculated using 1,1,3,3-tetraethoxypropane as the standard and expressed as nanomoles of MDA per gram of tissue.
PCO assays
Protein oxidation was determined based on the reaction of the carbonyl groups with 2,4-dinitrophenylhydrazine to form 2,4-dinitrophenylhydrazone, as described by Reznick and Packer method. 27 Samples were read at 370 nm and carbonyl content was calculated using the molar absorption coefficient for aliphatic hydrazones (22,000 M/cm) and expressed as nanomoles of carbonyl per milligram of protein.
NO production
Nitric oxide (NO) was determined by measuring the stable product nitrite through the colorimetric assay described by Archer method. 28 Samples were analyzed at 550 nm on a microplate spectrophotometer. Nitrite concentrations were calculated using a standard curve prepared with sodium nitrite (0–80 mM). Results were expressed as micromoles per milligram of protein.
Determination of GSH levels
Reduced glutathione (GSH) content of cerebellum was determined by Ellman’s method 29 modified by Jollow et al. 30 based on the development of a yellow color when 5,5-dithio-bis-2-nitrobenzoic acid is added to the compounds containing sulfhydryl groups. The absorbance was measured at 412 nm after 10 min. Total GSH content was expressed as nanomoles per milligram of protein.
Assay of antioxidant and glutathione metabolizing enzymes
Catalase (CAT) was assayed by the decomposition of hydrogen peroxide (H2O2) according to the Aebi method. 31 Decrease in absorbance due to H2O2 degradations was monitored at 240 nm for 1 min, and the enzyme activity was expressed as micromoles of H2O2 consumed per minute per milligram of protein.
Total suproxide dismutase (SOD) activity was evaluated by measuring the inhibition of pyrogallol activity as described by Marklund and Marklund. 32 One unit (U) corresponds to the enzyme activity required to inhibit half of the oxidation of pyrogallol. SOD activity was expressed as one unit per milligram of protein.
Glutathione peroxidase (GPx) activity was measured according to Flohe and Gunzler. 33 The enzyme activity was expressed as nanomoles of GSH oxidized per minute per milligram of protein.
Glutathione-S-transferase activity (GST) was determined following the procedure of Habig et al. 34 The change in color was monitored by recording absorbance (340 nm) at 30 s intervals for 3 min. The enzyme activity was calculated as nanomoles of C-DNB conjugates formed per minute per milligram of protein.
Determination of Na+/K+-ATPase activity in cerebellum tissue
Na+/K+-ATPase was assessed in cerebellum tissue according to the Quigley and Gotterer method. 35 The Na+/K+-ATPase activity was measured as the difference between total ATPase activities and ouabain-insensitive ATPase activities. The inorganic phosphate released by the action of ATPases was estimated by the Fiske and Subbarrow method. 36
Determination of AChE activity in the cerebellum tissue
AChE activity was determined in the cerebellum tissue according to the Ellman et al. method 37 and was expressed as micromole of thiocholine released per minute per milligram of protein.
Quantification of protein
The protein concentration in the homogenates was measured by the Lowry et al. method. 38
Statistical analysis
The data were analyzed using the statistical package program Stat view 5 Software for Windows (SAS Institute, Berkley, California, USA). Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Fisher’s protected least significant difference test as a post hoc test for the comparison between groups (treated groups (LTC, LTC + vit C, and vit C) vs. (controls)) and (LTC + vit C) versus (LTC and vit C). Student’s unpaired t test was also used when comparison between two groups was required. All values were expressed as mean ± standard error. Correlation coefficients were calculated using correlation matrix and regression analysis. Differences were considered significant, if p < 0.05.
Results
Concentration of LPO, PCO and NO
Statistically significant changes (F = 13.63, 8.51, 21.33, respectively; p < 0.01) in MDA, protein carbonyls (PCO) and nitrite formation were observed in the cerebellum of the LTC-treated group compared with the normal saline-treated controls (Table 1). Vit C cotreatment significantly (p < 0.05) decreases the cerebellum content of MDA, PCO and nitrite content by 28, 42 and 55%, respectively, when compared with LTC-treated group. Furthermore, there were no alterations in the LPO, nitrite and protein oxidation levels due to vit C treatment when compared with control group.
Effect of vit C (200 mg/kg bw) on LTC-induced alterations in the levels of lipid peroxidation (MDA), PCO levels and NO formation in the cerebellum of control and treated ratsa
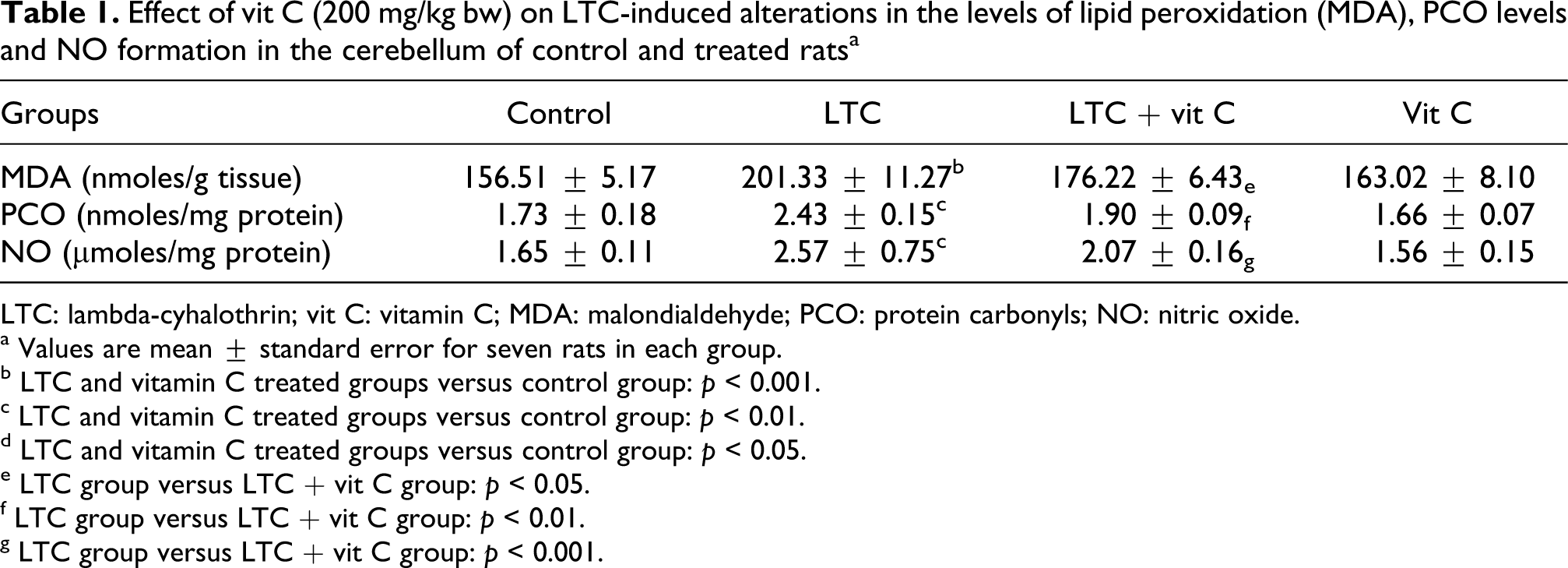
LTC: lambda-cyhalothrin; vit C: vitamin C; MDA: malondialdehyde; PCO: protein carbonyls; NO: nitric oxide.
a Values are mean ± standard error for seven rats in each group.
b LTC and vitamin C treated groups versus control group: p < 0.001.
c LTC and vitamin C treated groups versus control group: p < 0.01.
d LTC and vitamin C treated groups versus control group: p < 0.05.
e LTC group versus LTC + vit C group: p < 0.05.
f LTC group versus LTC + vit C group: p < 0.01.
g LTC group versus LTC + vit C group: p < 0.001.
Glutathione content and AChE activity
The contents of reduced GSH and AChE activity significantly (F = 23.39, 9.56, respectively; p < 0.05) decreased in the cerebellum of LTC-treated rats (Figure 1). A significant recovery (p < 0.05) in cerebellum GSH content and AChE activity was observed in LTC + vit C group compared with treated group (LTC). Vit C alone did not show any change in cerebellum GSH content and AChE activity when compared with controls.

Effect of vit C (200 mg/kg bw) on LTC-induced alterations in the levels of glutathione levels and AChE activity in the cerebellum of control and treated rats. Values are mean ± standard error for seven rats in each group. LTC + vit C treated groups versus control group: *p < 0.05, **p < 0.01, ***p < 0.001; LTC group versus LTC + vit C group: ##p < 0.05, ##p < 0.01, ###p < 0.001. GSH: reduced glutathione; AChE: acetylcholinesterase; LTC: lambda-cyhalothrin; vit C: vitamin C.
Antioxidant enzyme activities
The effects of LTC treatment on the activities of SOD, CAT and GPx in cerebellum are shown in Figure 2. Activities of SOD, CAT and GPx in cerebellum were significantly decreased (F = 9.22, 13.54, 17.13, respectively; p < 0.05) upon LTC treatment. Vit C administrated at a concentration of 200 mg/kg in LTC-treated rats improved significantly the activities of CAT, SOD, GPx and GST in the cerebellum of LTC + vit C group when compared with their respective control values. One-way ANOVA showed marked differences between the groups. Vit C alone did not produce any significant changes in antioxidant enzyme activities.
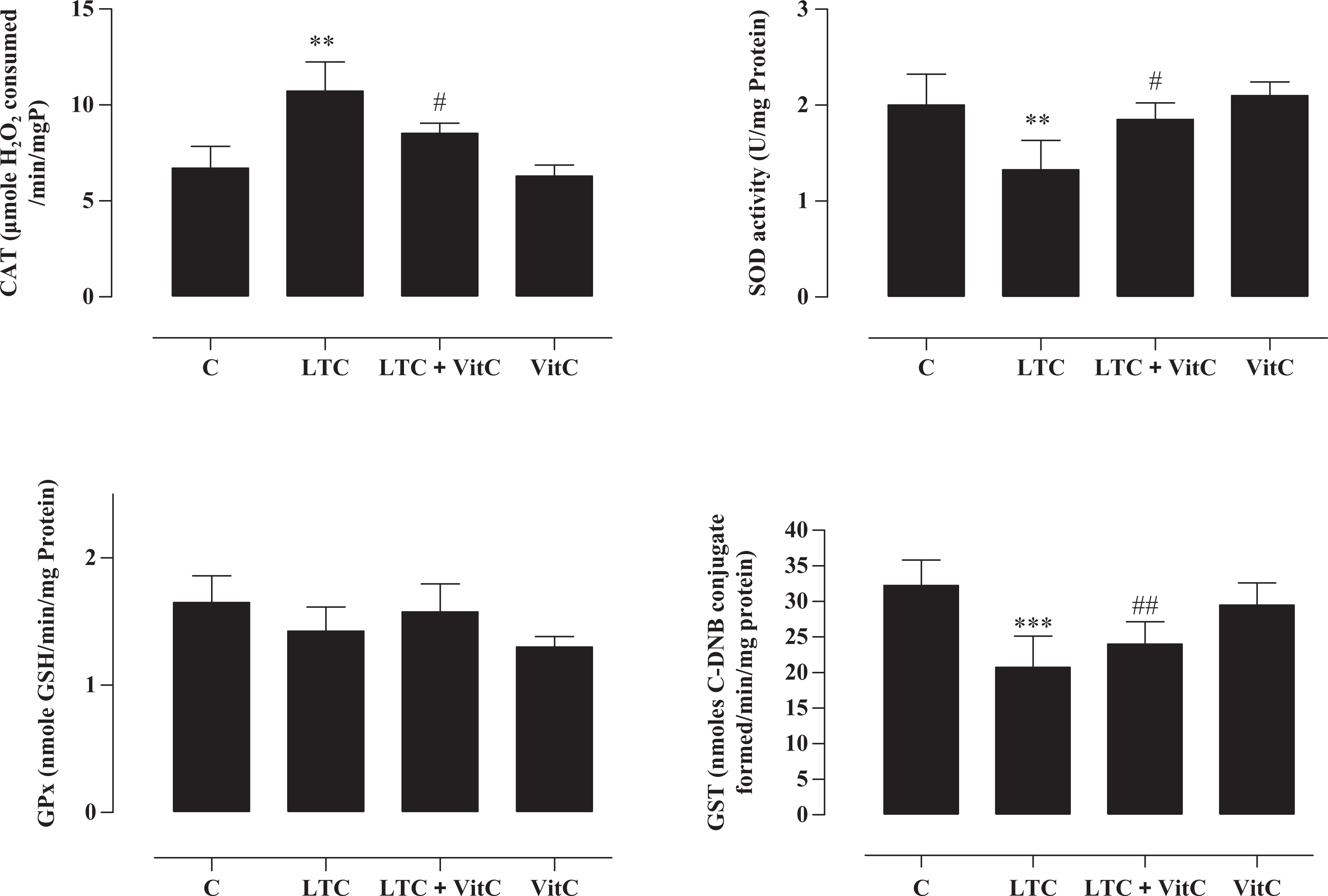
Effect of vit C (200 mg/kg bw) on LTC-induced alterations in the antioxidant enzyme activities in the cerebellum of control and treated rats. Values are mean ± standard error for seven rats in each group. LTC + vit C treated groups versus control group: *p < 0.05, **p < 0.01, ***p < 0.001; LTC group versus LTC + vit C group: ##p < 0.05, ##p < 0.01, ###p < 0.001. LTC: lambda-cyhalothrin; vit C: vitamin C.
Determination of Na+/K+-ATPase activity in cerebellum tissue
Figure 3 illustrates the activity of the membrane-bound Na+/K+-ATPase in the cerebellum tissue of rats. LTC treatment caused a significant decrease in the activity of Na+/K+-ATPase when compared with control. Vit C cotreatment significantly prevented the decrease in the enzyme activity induced by LTC treatment. Vit C administration alone did not produce any significant changes and the activity was similar to that of the control.

Effect of vit C (200 mg/kg bw) on LTC-induced alterations in the Na+/K+-ATPase activity in the cerebellum of control and treated rats. Values are mean ± standard error for seven rats in each group. LTC + vit C treated groups versus control group: *p < 0.05, **p < 0.01, ***p < 0.001; LTC group versus LTC + vit C group: ##p < 0.05, ##p < 0.01, ###p < 0.001. LTC: lambda-cyhalothrin; vit C: vitamin C.
Histopathology of the cerebellum
Under light microscopy, Figure 4 shows the histological aspect of cerebellum in all groups of rats. For control group, a typical uniform structure of Purkinje cell (PC) monolayer was located between the molecular layer and granular layer Figure 4A. However, in LTC-treated rats, the alignment of PCs was disturbed and the PC layer showed scattered, irregular and densely stained somas in Figure 4B. In rats treated with vit C (200 mg/kg) in combination with LTC, improvement in some cells was observed, showing normal PC morphology and normal PC layer alignment in Figure 4C. In vit C group alone, PCs structure is similar to that of the control group in Figure 4D.

Photomicrographs of rat cerebellum (hematoxylin and eosin, ×400) from (a) control group with normal appearance of PCs; (c) LTC-treated group showing necrotic PCs; (d) vit C + LTC-treated group displaying remarkable improvement in the histological appearance of PCs; (b) vit C alone treated group showing normal PCs similar to that observed in controls (normal PCs and necrotic PCs). LTC: lambda-cyhalothrin; vit C: vitamin C; PCs: Purkinje cells.
Discussion
Numerous studies demonstrate that the changes in the antioxidant system and impairment of sodium and potassium transport are essential factors involved in the brain tissue disturbance in various kinds of oxidative injury in the central nervous system. 39 –41 The potency of SPs as a neurotoxin has been demonstrated in both in vitro and in vivo studies. 42 –44 Similar results were also reported in the epidemiological study which was observed in farmers who used pyrethroids for crops, a damage in their central and peripheral nervous systems resulting in headaches, fatigue, lack of concentration, muscle spasms as well as impaired vision, hearing and taste. 45 It seems that once absorbed, pyrethroids rapidly distribute throughout the body. Their high lipophilicity and lack of exclusion by the multidrug transporter glycoprotein ensure ready entry into the brain. 46 Accordingly, the study of brain enzyme activities, such as AChE, is essential in detecting the neurotoxic effects of pesticides. Besides, several studies reported that the free radical production could at least in part be associated with the decrease in the activity of AChE in the brain, 47 leading to brain damages. Also, it has been shown that the decreased AChE activity leads to the accumulation of acetylcholine, which causes cholinergic hyperactivity, convulsion and status epilepticus. 48 Since brain AChE activity is an important regulator of the behavioral processes, the decreased level of AChE in plasma and brain might be one of the indicators for pesticide-induced hurdle in the brain. The administration of vit C at a dose of 200 mg/kg restored the activities of AChE in LTC-exposed rats, which indicates the preservation of cerebellum function from LTC-induced toxic effects. The effectiveness of vit C may be attributed to its scavenging property and capability to reduce free radicals generated by LTC exposure. 7,8,49,50 According to World Health Organization, 51 LTC toxicity might be due to the release of cyanohydrins, which are unstable under physiological conditions and further decomposed to cyanides and aldehydes which in turn could act as a source of free radicals. In addition, our results corroborate a previous study, which demonstrated that the by-products of LPO had been shown to cause profound alterations in the structural organization and functions of the cell membrane including decreased membrane fluidity, increased membrane permeability, inactivation of membrane-bound enzymes like ATPases and loss of essential fatty acids. 52 In our study, the elevation in MDA and increased formation of PCO with reduced levels of GSH in LTC-intoxicated rat brain might be due to the overproduction of free radicals and LPO end products, which leads to oxidative modifications of proteins.
Thiols (SH) are recognized to play a fundamental antioxidant role by protecting cellular and extracellular functions against oxidative stress. In this context, we observed that the LTC markedly reduced the sulfhydryl groups, indicating that the SP compound reduced the nonenzymatic antioxidant defenses in the cerebellum of the rats. It is described in the literature that the synthetic pesticides decreases the cellular content of reduced SH with a consequent increase in the production of reactive oxygen species (ROS). 7,8 Mammalian cells from different tissues, including the brain, possess a system that regulates the redox status of cellular SH and protects SH-containing proteins from excessive oxidation. It includes low-molecular-weight donors of SH groups and enzymes, which can catalyze the reduction in SH groups in proteins and detoxify pro-oxidants by conjugation with GSH. 53 We propose that the decrease in SH groups observed in our in vivo model could provide evidence for increased oxidative damage of proteins in the cerebellum of adult rats. Moreover, it is known that the Na+/K+-ATPase activity is involved in the activity-dependent synaptic plasticity as well as in the memory and learning mechanisms in the nervous system. 54 In our study, the decrease in Na+/K+-ATPase activity in the cerebellum tissue after LTC exposure indicates its efficacy to alter membrane integrity and fluidity in mammalian cells. Similar results are confirmed by Kakko et al. 55 who demonstrated that the exposure to pyrethroids caused a decrease in ATPase activity in a dose-dependent manner and that the changes in the neuronal sodium homeostasis is one of the mechanisms of neurotoxicity. Coadministration of vit C with LTC exposure restored the activities of ATPases, which reveals that vit C preserves the integrity of cellular membrane and the normal physiological functions of the cerebellum. The inhibition of radical generation with the preservation of mitochondrial function might contribute to the enhancement of ATPase activity in LTC-intoxicated rats treated with vit C. We also demonstrated that LTC modified the activity of the antioxidant enzymes by reducing their activities. We next investigated whether NO synthesis could be induced by this xenobiotic, since SOD activity can be altered by peroxinitrite, the most harmful NO derivative. 56 The increase in reactive nitrogen species (RNS) production result from excess or deregulated NO reacting with ROS. NO is a signaling molecule that reacts with superoxide radicals to generate RNS, including peroxynitrite, which is a very reactive molecule that can modify biomolecules, including DNA, lipids and proteins. 57,58 The administration of vit C at a dose of 200 mg/kg significantly restored the activities of antioxidants with a depletion of PCO formation in LTC-intoxicated rats, which could be due to the attenuation of MDA by the free radical scavenging activity of vit C. These biochemical alterations were correlated with the histological aspect of cerebellum, which demonstrated that the alignment of PCs observed in control animals was disturbed and the PC layer showed scattered, irregular and densely stained somas in LTC-treated group. The administration of vit C in combination with LTC, an improvement of some cells was observed, showing normal PC morphology and normal PC layer alignment in Figure 4C.
In conclusion, our data indicate that vit C has beneficial actions against LTC-mediated toxic effects in the brain via inhibiting LPO and subsequently restoring the AChE and membrane-bound enzymes. Therefore, vit C may be considered a therapeutic agent for enzyme structure. Thus, the decreased activities of cerebellum antioxidant systems indicate the accumulation of free radicals and increased level of MDA, which increase the oxidative threat in tissue.
Footnotes
Funding
The present work was supported by the grants of Direction Générale de la Recherche Scientifique en Tunisie (DGRST) (Appui à la Recherche Universitaire de base, UR11ES70) Tunisia.