
Abstract
Although the endocrine-disrupting bioactivity of parabens is weakly estrogenic (parabens are xenoestrogens), their combined synergistic effect is unknown. The aim of this study was to investigate the effects of methyl paraben (MP), ethyl paraben (EP), propyl paraben (PP), isopropyl paraben (IPP), butyl paraben (BP), and isobutyl paraben (IBP), either alone or in combination (MP + EP + PP + BP; PP + IPP; and BP + IBP) on the induction of the estrogenic biomarker gene, calbindin-D9k (CaBP-9k), in rat pituitary lactosomatotrophic GH3 cells. The expression of CaBP-9k mRNA and protein was analyzed using real-time PCR and Western blot analysis, respectively. After 24 h of treatment, a significant increase in CaBP-9k expression was observed. This was dependent upon the length of the paraben alkyl chains (shortest in MP and longest in IBP). Interestingly, the synergistic effects of these paraben combinations were observed at a dose (10−5 M) of these parabens, which induced the highest expression of CaBP-9k mRNA and protein. To investigate the involvement of estrogen receptors (ERs) and progesterone receptors (PRs), through which parabens exert their effects, the expression levels of ERα and PR-B were also examined. The expression of ERα mRNA and protein fluctuated after paraben treatment in GH3 cells, which was not significant. However, the expression level of ERα gene was induced when cotreated with 17β-estradiol (E2) and ICI 182, 780 (estrogen receptor antagonist). The different combinations of parabens induced the expression of the PR-B gene, which was abolished by cotreatment with ICI 182,780. The expression patterns of CaBP-9k and PR-B genes appeared to be similar in response to paraben treatments. This implied that CaBP-9k expression in GH3 cells may be induced by parabens via a PR-mediated pathway. Taken together, these results suggest that exposure to multiple parabens at low concentrations may increase their synergistic estrogenic activities in GH3 cells through a PR-mediated pathway.
Introduction
Methyl paraben (MP), ethyl paraben (EP), propyl paraben (PP), isopropyl paraben (IPP), butyl paraben (BP), and isobutyl paraben (IBP) are widely used as antimicrobial agents in cosmetics, toiletries, and various drugs because they have relatively low toxicity and long history of safe use. 1 In general, parabens are considered safe, nontoxic, nonirritating, and nonsensitizing to both human and animals. 2 However, several recent studies show that exposure to parabens may disrupt the endocrine system and thus be harmful to animal and human health. 1,3,4 As with many other estrogenic compounds that have structures similar to natural and synthetic estrogens, parabens can bind to estrogen receptors (ERs), progesterone receptors (PRs), and pS2 and interfere with the actions of endogenous steroid hormones. 5-7 The previous studies using MCF-7 human breast cancer cells, ZR-75-1, indicated that parabens act primarily through nuclear hormone receptors. 5,6,8-10 A linear relationship between paraben levels and cell proliferation and ER-binding capacity has been reported. 11,12 The potent estrogenic activity of parabens is dependent upon the length of their alkyl side chains. 6,13 Routledge et al. reported that an increase in the size of the alkyl group may enhance paraben-induced transactivation of ERs in vitro. 14 Furthermore, parabens may also increase the risk of estrogen-related end points, including an increased incidence of female breast cancer, interference with male reproductive function, and the development of melanomas. 15
The previous studies showed that calbindin-D9k (CaBP-9k), a cytosolic calcium binding protein, is expressed in a variety of mammalian tissues, such as uterus, placenta, intestine, kidney, bone, and pituitary gland. 16–18 CaBP-9k is involved in interstitial calcium absorption and is regulated at the transcriptional and posttranscriptional levels by 1,25-dihydroxyvitamin (D3). In the rat uterus, CaBP-9k expression is strongly regulated by sex steroid hormones. The mechanisms underlying the regulation of CaBP-9k expression in uterine myometrium and stromal cells are closely associated with the direct stimulatory effects of 17β-estradiol (E2) and/or the indirect inhibitory effects of progesterone (P4). 18 The GH3 cell line, a well-established pituitary cell line, is sensitive to estrogenic stimulation. 19 Dang et al. showed that a single treatment with environmental compounds such as 4-tert-octylphenyl (OP), 4-nonylphenol (NP), or bisphenol A (BPA) upregulates CaBP-9k expression in GH3 cells. 20 Both estrogen-responsive and P4-responsive elements are present in the CaBP-9k promoter and mediate the transcriptional regulation of CaBP-9k in the rat uterus. 21 A recent study showed that BP enhanced GH3 cell proliferation by acting as a weak thyroid hormone receptor agonist. 22
Chemicals are usually present in the environment as mixtures, rather than as single entities, and studies of their effects are limited. One study showed that a combination of different pesticides enhanced the proliferation of MCF-7 cells in the E-Screen in what appeared to be a synergistic manner. 23 Also, the combination of some polychlorinated biphenyls and environmental estrogens induces sex reversal at concentrations that have no effect when the chemicals are used individually. 24 Interaction of estrogenic chemicals with each other leads to produce significant effects, even when combined at concentrations at which, when used alone, they have no observable effects. 25
The estrogenic activity of parabens has been evaluated in recent studies. 7 However, the interaction between these chemicals when present at low concentrations is still unknown. Therefore, the aim of this study was to investigate the effects of parabens, both alone and in combination, on GH3 cells by monitoring the expression of the estrogenic biomarker gene, CaBP-9k.
Materials and methods
Chemicals
MP, EP, PP, and E2 were obtained from Sigma Aldrich, Inc. (St Louis, MO, USA). BP was purchased from Sigma Aldrich GmbH (Steinheim, UK). IPP and IBP were obtained from Tokyo Kasei Kogyo Co. Ltd (Tokyo, Japan). The ER antagonist ICI 182,780 was obtained from Tocris (Ellisville, MO, USA). All chemicals were dissolved in dimethyl sulfoxide (DMSO). Rabbit CaBP-9k and the goat-anti-rabbit antibodies were provided by Swant (Bellinzona, Switzerland). Anti-ERα and PR-B antibodies and horseradish peroxidase (HRP) conjugated anti-mouse IgG and anti-rabbit antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Cell culture and treatment
GH3 cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Gibco BRL, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS; Gibco BRL, Grand Island, NY, USA), 100 U/ml penicillin and 100 μg/ml streptomycin (Gibco, Grand Island, NY, USA) at 37°C in a humidified atmosphere of 95% O2 and 5% CO2.
For chemical treatment, 2.1 × 106 cells were plated in 6-well plates (NUNC, Roskilde, Denmark) and grown until 70–80% confluent. To ensure depletion of steroid hormones and growth factors from the cells, the medium was replaced with starvation media containing phenol red-free DMEM supplemented with 5% charcoal–dextran-stripped FBS and 100 U/ml penicillin–streptomycin, as previously described. 20 The GH3 cells were maintained in starvation media for 7 days before exposure to parabens (either singly or combined) at a concentration 10−5 M. Each chemical was dissolved in DMSO and added to the starvation media at a final DMSO concentration of 0.1%. E2 (10−9 M) and DMSO alone (0.1%) were used as positive and negative controls, respectively. The cells were harvested after 24 h and mRNA and protein levels were measured. To identify the mechanism of CaBP-9k induction mediated by these endocrine-disrupting chemical combinations, the cells were treated with different paraben combinations (either two or four compounds mixed at a ratio of 1:1; final concentration 10−5 M) in the presence or absence of ICI 182,780 (10−7 M; added 1 h prior to parabens exposure). After 24 h, whole cells were harvested for mRNA and protein analysis. All experiments were performed in triplicate.
RNA extraction and quantitative real-time PCR
Total RNA was extracted using the Trizol reagent (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s protocol. The concentration of RNA was determined by absorbance at 260 nm. The quality of the RNA was confirmed using visual inspection following electrophoresis on 1% agarose gels. Briefly, total RNA (1 μg) was reverse transcribed into first-strand complementary DNA (cDNA) using moloney murine leukemia virus reverse transcriptase (Invitrogen) and a random primer (a 9-mer, Takara Bio, Otsu, Shiga, Japan). Each cDNA template (1 μl) was analyzed by the addition of 10 μl of 2× SYBR® Premix Ex TaqTM (TaKaRa Bio. Inc., Otsu, Shiga, Japan) and 10 pM of each primer. Amplification was performed in a 7300 Real-time PCR System (Applied Biosystems, Foster, CA, USA) using the following parameters: denaturation at 95°C for 5 min, 40 cycles of denaturation at 95°C for 30 sec, and annealing and extension at 60°C for 30 sec, and 72°C for 45 sec. The oligonucleotide primer sequences used to amplify CaBP-9k (product size, 314 bp) were 5-AAGAG CATTT TTCAA AAATA-3′ (sense) and 5-GTCTC AGAAT TTGCT TTATT-3′ (antisense). The oligonucleotide primer sequences used to amplify the cytochrome c oxidase subunit 1 (1A; product size, 293 bp) were 5-CCAGG GTTTG GAATT ATTTC-3′ (sense) and 5-GAAGA TAAAC CCTAA GGCTC-3′ (antisense). The relative expression levels of CaBP-9k (normalized to that of 1A) in each sample were determined using RQ software (Applied Biosystems). All real-time PCR experiments were repeated twice.
Western blot analysis
Protein samples were extracted using pro-prep protein extraction solution (iNtRON Biotechnology, Seoul, South Korea), according to the manufacturer’s instructions. Protein concentrations were determined using a BCA assay (Sigma, St Louis, CA, USA). Cytosolic proteins (50 μg per lane) were resolved in 10% SDS-PAGE gels and transferred onto polyvinylidene fluoride (PVDF) membranes using a trans-blot cell (Bio-Rad, Hercules, CA, USA). The membranes were then blocked for 2 h with phosphate-buffered saline containing 0.1% Tween-20 (PBS-T) and 5% skim milk (DifcoTM, Sparks, MD, USA). The membrane was then incubated with the indicated primary antibodies diluted in 1% BSA for 3 h (for glyceraldehyde 3-phosphate dehydrogenase [GAPDH] and CaBP-9k) or overnight (for PR-B and ERα) at room temperature. The primary antibodies for CaBP-9k (9 kDa), PR-B (116 kDa), and ERα (66 kDa) were diluted into 1:500 and the anti-GAPDH (38kDa) antibody was diluted into 1:2000. HRP-conjugated anti-mouse IgG (for CaBP-9k and PR-B) and anti-rabbit IgG (for GAPDH and ERα) were diluted 1:3000 in PBS-T/2.5% skim milk. Immunoreactive proteins were visualized by exposing the membranes to an X-ray film. The films were then scanned and the band intensities and optical densities were quantified using Image J software (version 1.52, Wayne Rasband, NIH, Baltimore, MD, USA) after correction by background subtraction and then normalized against GAPDH expression. 26
Data analysis
Data were expressed as the mean ± SD and analyzed using one-way analysis of variance (ANOVA) followed by Tukey’s test for multiple comparisons. Analysis was performed using Graph Pad Prism4 for Windows Edition (Graph Pad Software Inc., San Diego, CA, USA). A p value of <0.05 was considered statistically significant.
Results
Effects of paraben combinations on the induction of CaBP-9k expression
GH3 cells were treated with parabens, either individually or in combination, at a final concentration of 10−5 M. Samples were collected 24 h after treatment and mRNA and protein levels were analyzed. As shown in Figure 1A, exposure of GH3 cells to parabens, either in combination (MP + EP + PP + IPP, PP + IPP, or BP + IBP) or individually (MP, EP, PP, IPP, BP, and IBP) resulted in a significant increase in CaBP-9k transcription compared with vehicle alone. Interestingly, expression of CaBP-9k mRNA and protein was highest in cells treated with BP + IBP. E2 (positive control) led to significantly increased levels of CaBP-9k gene expression. Similarly, CaBP-9k protein levels were upregulated by both E2 and parabens, with the exception of MP and EP (Figure 1B). The difference between CaBP-9k transcription and translation induced by these parabens may be explained by mRNA stability and/or protein accumulation. In addition, the expression of both CaBP-9k mRNA and protein increased in parallel with the length and number of branches of the alkyl chains in the individual parabens. A significant increase in the levels of CaBP-9k mRNA and protein was observed after treatment with paraben combinations compared with single treatments, suggesting that, when combined, parabens interact in a synergistic manner.

Effects of parabens, either alone or in combination, at a concentration of 10−5 M on calbindin-D9k (CaBP-9k) mRNA and protein expression in GH3 cells. (A) CaBP-9k mRNA expression determined by real-time PCR. (B) CaBP-9k protein expression determined by Western blotting. CaBP-9k gene expression was normalized to that of internal control genes (1A for mRNA and glyceraldehyde 3-phosphate dehydrogenase [GAPDH] for protein). Data are expressed as the mean ± SD of triplicate experiments. a p < 0.05 versus vehicle; b p < 0.05 versus parabens (methyl- [MP], ethyl- [EP], propyl- [PP], and butyl paraben [BP]); c p < 0.05 versus parabens (propyl- [PP] and isopropyl paraben [IPP]); d p < 0.05 versus parabens (butyl- [BP] and isobutyl paraben [IBP]).
To further understand the mechanisms underlying CaBP-9k gene expression induced by the various paraben combinations, the expression of both ERα and PR-B was analyzed at the mRNA and protein level. As illustrated in Figure 2A and B, there were fluctuations in the expression levels of ERα mRNA and protein. These results highlight a difference between the expression of CaBP-9k and ERα, suggesting that CaBP-9k mRNA and protein expression may not be induced by parabens via an ERα-mediated pathway in GH3 cells. Interestingly, Figure 3A and B shows that the pattern of paraben-induced PR-B expression was similar to that of the CaBP-9k gene. PR-B was upregulated by exposure to both E2 and the various paraben combinations. This result indicates that CaBP-9k mRNA and protein expression may be induced via a PR-mediated pathway.

Effects of parabens, either alone or in combination, at a concentration of 10−5 M on the expression of estrogen receptor α (ERα) mRNA and protein in GH3 cells. (A) ERα mRNA expression determined by real-time PCR. (B) ERα protein expression determined by Western blotting. ERα gene expression was normalized to that of internal control genes (1A for mRNA and glyceraldehyde 3-phosphate dehydrogenase [GAPDH] for protein). Data are expressed as the mean ± SD of triplicate experiments.

Effects of parabens, either alone or in combination, at a concentration of 10−5 M on the expression of progesterone receptor B (PR-B) mRNA and protein in GH3 cells. (A) PR-B mRNA expression determined by real-time PCR. (B) PR-B protein expression determined by Western blotting. PR-B gene expression was normalized to that of internal control genes (1A for mRNA and glyceraldehyde 3-phosphate dehydrogenase [GAPDH] for protein). Data are expressed as the mean ± SD of triplicate experiments. a p < 0.05 versus vehicle; b p < 0.05 versus parabens (methyl- [MP], ethyl- [EP], propyl- [PP], and butyl paraben [BP]); c p < 0.05 versus parabens (propyl- [PP] and isopropyl paraben [IPP]); d p < 0.05 versus isobutyl paraben (IBP).
Effects of ICI 182,780 on paraben-induced CaBP-9k
To identify the steroid hormone receptor pathway involved in transcriptional and translational upregulation of CaBP-9k expression by the various paraben combinations, GH3 cells were pretreated with the ER antagonist, ICI 182,780. CaBP-9k mRNA and protein levels were then analyzed by real-time PCR and Western blotting, respectively. As seen in Figure 4A and B, the induction of CaBP-9k mRNA and protein by E2 and the different paraben combinations was completely abolished by cotreatment with ICI 182,780 at a concentration of 10−7 M.

Effects of the antiestrogen, ICI 182,780, on the calbindin-D9k (CaBP-9k) mRNA and protein expression after treatment with various paraben combinations at 10−5 M. GH3 cells were pretreated with ICI 182,780 (10−7 M) for 1 h prior to paraben exposure. (A) CaBP-9k mRNA expression determined by real-time PCR. (B) CaBP-9k protein expression determined by Western blotting. CaBP-9k gene expression was normalized to that of internal control genes (1A for mRNA and glyceraldehyde 3-phosphate dehydrogenase [GAPDH] for protein). Data are expressed as the mean ± SD of triplicate experiments. a p < 0.05 versus vehicle; b p < 0.05 versus chemical-only treated groups.
Next, the expression levels of ERα and PR-B mRNA and protein were analyzed. Although ERα mRNA and protein expression levels were not changed after treatments with E2 or paraben combinations, there was a significant increase in ERα mRNA and its protein levels, respectively, following treatment with ICI as shown in Figure 5A and B. However, the mRNA and protein expression levels of PR-B were induced by E2 or paraben combinations, which were completely blocked by ICI treatment as seen in Figure 6A and B. This expression pattern appears to be in parallel with that of CaBP-9k gene. This result provides further evidence that exposure of GH3 cells to estrogenic parabens specifically upregulates PR levels, suggesting that CaBP-9k gene expression may be induced, in part, through PRs in response to various combinations of estrogenic parabens.

Effects of treatment with various paraben combinations (10−5 M) with/without ICI 182,780 on estrogen receptor α (ERα) mRNA and protein expression. GH3 cells were pretreated with ICI 182,780 (10−7 M) for 1 h prior to paraben exposure. (A) ERα mRNA expression determined by real-time PCR. (B) ERα protein expression determined by Western blotting. ERα gene expression was normalized to that of internal control genes (1A for mRNA and glyceraldehyde 3-phosphate dehydrogenase [GAPDH] for protein). Data are expressed as the mean ± SD of triplicate experiments. a p < 0.05 versus vehicle; b p < 0.05 versus chemical-only treated groups.
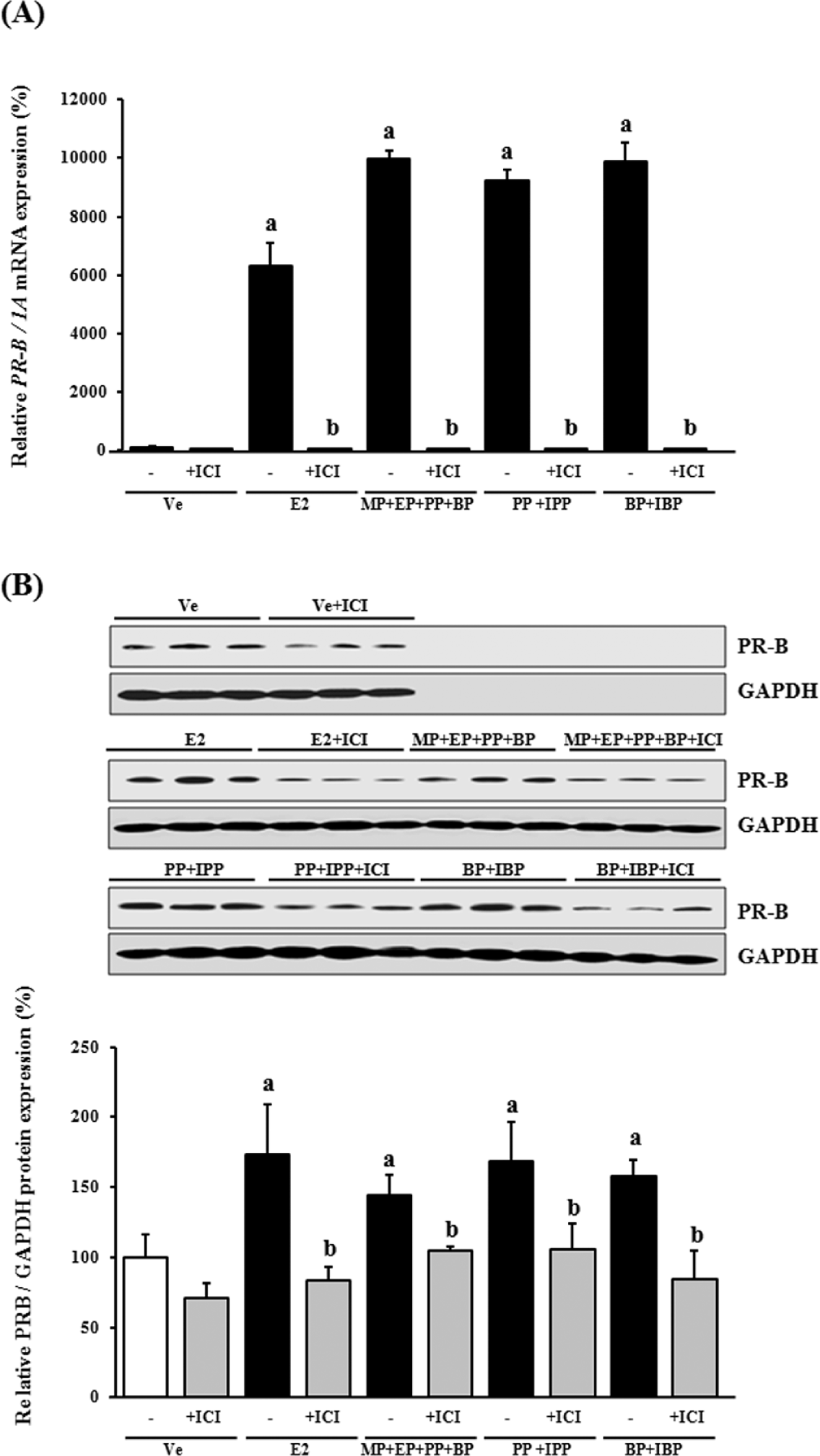
Effects of treatment with various paraben combinations (10−5 M) with/without ICI 182,780 on progesterone receptor B (PR-B) mRNA and protein expression. GH3 cells were pretreated with ICI 182,780 (10−7 M) for 1 h prior to paraben exposure. (A) PR-B mRNA expression determined by real-time PCR. (B) PR-B protein expression determined by Western blotting. PR-B gene expression was normalized to that of internal control genes (1A for mRNA and glyceraldehyde 3-phosphate dehydrogenase [GAPDH] for protein). Data are expressed as the mean ± SD of triplicate experiments. a p < 0.05 versus vehicle; b p < 0.05 versus chemical-only treated groups.
Discussion
It is well documented that the CaBP-9k gene is controlled by sex hormones. The expression of the CaBP-9k gene is upregulated by estrogen and downregulated by P4 during the estrous cycle and early pregnancy in the rat uterus. 27,28 Recently, the estrogenic activity of a panel of structurally related parabens, including MP, EP, PP, IPP, BP, and IBP, were evaluated in the GH3 cell line by monitoring the expression of the estrogenic biomarker gene, CaBP-9k. Indeed, several studies show that the CaBP-9k gene can be used as a potential biomarker for assessing the ‘estrogenicity’ of environmental compounds. 29,30 Expression of the CaBP-9k gene is regulated by some xenoestrogenic chemicals, such as OP, NP, BPA, genistein, and parabens in many cell types, including immature rat uterus, 7,21,31 rodent pituitary gland, 17,18 and GH3 cells. 20 This suggests that this gene is a useful biomarker for evaluating the ‘estrogenicity’ of environmental estrogen-like compounds.
Previous studies showed that parabens have structural similarities to alkyl phenols and they are, therefore, able to induce weak estrogenic effects both in vitro and in vivo. 32 Because of this bioactivity, parabens are thought to disrupt the development of the male and female reproductive tracts at critical phases during maturation. 14,33,34 Parabens are readily absorbed through the skin and gastrointestinal tract, after which they are metabolized via hydrolysis to p-hydroxybenzoic acid. 3 The bioactivity of ingested parabens has adverse reproductive effects due to their rapid absorption, metabolism and excretion as urinary conjugates. 3,35 Moreover, parabens might have achieved full agonist responses at lower concentrations if the assays had been longer in duration. 15 For instance, there were no estrogenic responses to MP, EP, and PP at a dose of 100 mg/kg per body weight/day in a murine uterotrophic assay; however, rat uterine weight increased after a high dose of 600 mg/kg per body weight/day. 10,32 The potential ‘estrogenicity’ of parabens has been demonstrated in various cell-based assays, including the MCF-7 10,36 and ZR-75-1 breast cancer cell lines. 8,37 Parabens bind to the ER, activate genes controlled by these receptors, stimulate cell growth, and increase the immunoreactivity of ERs. 38 A panel comprising MP, EP, n-PP, and n-BP increased the proliferation rate of MCF-7 breast cancer cells. 5
In the present study, we showed that CaBP-9k expression in GH3 cells is a useful ‘readout’ for evaluating the estrogenic potential of environmental chemicals such as parabens. The results showed a marked increase in CaBP-9k mRNA and protein expression (2.3–3.7-fold and 4.2–8.7-fold, respectively) after treatment with the doses (10−5 M and 10−4 M) of long alkyl chain parabens (PP, IPP, BP, and IBP). However, the induction of CaBP-9k mRNA and protein expression was weak compared with that induced by the positive control, E2 (10−9 M; 4.2-fold and 9.8-fold, respectively). Similarly, a weak relative binding affinity (in the order of 0.001% for parabens compared with estrogen) has been reported from in vitro assays using MCF-7, ZR-75-1, HELN, and HELNα and HELNβ (HeLa cells stably transfected with an ERE-driven luciferase plasmid) cell lines, in which E2 was shown to act maximally between 10−10 M and 10−8 M and parabens between 10−5 M and 10−4 M. 5,8,11,39 We also showed that increased alkyl chain length correlated with increased CaBP-9k expression, suggesting that CaBP-9k expression in GH3 cells is a sensitive and effective measure of the estrogenic activity of environmental chemicals and their structural relatives.
The present study showed that various combinations of parabens exert a synergistic effect on CaBP-9k expression at a dose of 10−5 M. It was also showed that increasing the length of the alkyl chain of the paraben side group correlates with increased CaBP-9k gene expression. Arnold et al. reported the E2 and 17α-estradiol (EE) act synergistically in yeast strains expressing human ERs; this synergistic activity being evident at submaximal concentrations (1 nM) of E2. 40
To identify the biological pathways involved in the induction of CaBP-9k gene expression in GH3 cells following exposure to various paraben combinations, an antiestrogen (ICI 182,780) was employed to examine the potential involvement of ER or PR in paraben-induced responses in vitro. CaBP-9k mRNA and protein expression were completely abolished in cells co-treated with ICI 182,780. However, ERα mRNA and protein levels increased after coexposure to ICI 182,780 compared with exposure to E2, parabens alone, or in combination. However, its mechanism is not known in regard to increased expression of ERα mRNA and protein levels by ICI 182,780. In other studies, a significant increase in ERα mRNA and protein expression (a minimum of 30% and 20%, respectively) compared with parabens alone was observed. ICI 182,780 binds, inhibits, and promotes the degradation of ERs 41,42 and decreases the expression of ERα, but not that of ERβ or the androgen receptor (AR), in rats. 43 Moreover, ICI 182,780 reduces ER steady-state levels by increasing the turnover of the protein. 44-46 In another study, measurement of uterine nuclear ER levels showed a 20% reduction in total ER levels within 0.5 h of ICI 182,780 treatment. ICI also affected ER expression in an in vitro model using mouse TM4 estrogen-responsive cells and dramatically reduced ER levels to 5% of the control value by 4 h. 45 However, ICI 182,780 may affect ER downregulation in a dose-dependent manner. An approximately 70% reduction in ER expression was observed with a single dose (250 mg) of ICI 182,780. 47 In other study, ICI 182,780 mimicked some E2 actions in gene expression in the liver of sea bream by upregulating ERα. 48 In this study, ICI 182,780 does not seem to play a role as an antiestrogen since it did not block the effects of an E2 challenge in GH3 cells, suggesting that the biological activity of parabens may not be involved in an ER-mediated signaling pathway in these cells.
To further demonstrate the potential impact of various paraben combinations on CaBP-9k gene expression, PR-B levels were analyzed. PR is an E2-responsive gene and an indicator of ERα-mediated transcription in the rat uterus. 49 PR expression is induced via an ER-activated protein-1 pathway. 50 In the present study, we found that the patterns of CaBP-9k and PR-B gene expression in response to paraben treatment were similar and that pretreatment with ICI 182,780 completely attenuated PR-B expression. In addition, we also observed downregulated ERα expression and upregulated PR levels following paraben exposure in vivo. 6,51 Previous studies show that ICI 182,780 blocks PR-mediated transcription in the immature rat uterus, 7 and in several cell lines, including HeLa, HepG2, and CV1 (which are considered the best cell lines for determining antiestrogenic activity). 52 In GH3 cells, the luciferase activity of ERE (estrogen response element) sequences was induced by E2 and paraben, which was similar to the enhanced expression of CaBP-9k and PR following paraben exposure. 53
Taken together, our results indicate that various paraben combinations can increase the expression of CaBP-9k mRNA and protein in GH3 cells, and its regulations may affect the steroid hormone receptors. In addition, an estrogenic activity of parabens was blocked by ICI 182,780, an ER antagonist, in GH3 cells. Nevertheless, further research is needed to elucidate the potential endocrine-disrupting activity of parabens and other exogenous combinations and their potential effects on human health.
Footnotes
H Yang and T-T Nguyen equally contributed as first authors.
This work was supported by a grant from the Next-Generation BioGreen 21 Program (No. PJ008323), Rural Development Administration, Republic of Korea.