
Abstract
Ipomoea carnea Jacq. ssp. fistulosa (Mart. Ex Choisy; Convolvulaceae; I. carnea) possesses a toxic component: an indolizidine alkaloid swainsonine (SW) that has immunomodulatory effects due to its inhibition of glycoprotein metabolism. It is also known that SW is excreted into both the amniotic fluid and milk of female rats exposed to I. carnea. Thus, the aim of this study was to determine whether SW exposure, either in utero or from the milk of dams treated with I. carnea, modulates offspring immune function into adulthood. In addition, adult (70 days old) and juvenile rats (21 days old) were exposed to I. carnea in order to evaluate several other immune parameters: lymphoid organs relative weight and cellularity, humoral and cellular immune responses. Offspring exposed to I. carnea during lactation developed rheumatoid arthritis (RA) in adulthood after an immunogenic challenge. In addition, both adult and juvenile rats exposed to I. carnea showed discrepancies in several immune parameters, but did not exhibit any decrease in humoral immune response, which was enhanced at both ages. These findings indicate that SW modulates immune function in adult rats exposed to SW during lactation and in juvenile and adult rats exposed to SW as juveniles and adults, respectively.
Introduction
Ipomoea carnea Jacq. ssp. fistulosa Choisy (Convolvulaceae; I. carnea) is a poisonous plant widely distributed in Brazil and in other tropical countries. 1 Two toxic components have been isolated from this plant: the nortropane alkaloid calystegines B1, B2, B3 and C1 and the indolizidine alkaloid swainsonine (SW). 2,3 SW’s toxic mechanism of action involves potent inhibition of two distinct intracellular enzymes: the acidic or lysosomal α-mannosidase and the Golgi mannosidase II; however, there is little information concerning the potential toxic effects of the nortropane alkaloids. It is known that calystegines B2 and C1 are glycosidase inhibitors affecting α-glucosidase and α- and β-galactosidase, respectively. 4 However, in a previous study conducted in our lab, all of the isolated calystegines from I. carnea did not produce toxic effect in rats. 5
The inhibition of α-mannosidase results in the lysosomal accumulation of incompletely processed oligosaccharides that are rich in α-mannosyl and β-N-acetyl glucosamine moieties inside vacuoles, which leads to a loss of cellular function and, ultimately, cell death. 6 Histologically, cellular vacuolization in Purkinje cells, thyroid follicles, exocrine pancreas, liver and kidney cells has been observed.
SW inhibition of the Golgi mannosidase II enzyme causes alterations in the N-linked glycoprotein process. 7 This, in turn, increases the production of high-mannose, hybrid or complex types of oligosaccharide structures that are components of hormones, cytokines, membrane receptors and adhesion molecules. 8 The modification resulting from glycoprotein metabolism affects immune system responses. Experimental studies conducted in vitro show that SW has potential immunomodulatory properties, such as increasing murine natural killer cell (NK) activity, 9 enhancing the generation of lymphokine-activated killer (LAK) cell activity, 10 increasing human large granular lymphocyte cytotoxicity against NK-resident colon carcinoma cells 11 and activation of resident tissue macrophages. 12 Also, the administration of I. carnea to rats enhances macrophage activity, such as phagocytosis and hydrogen peroxide release. 13
Previously, we administered I. carnea to female rats at two different stages of development: prenatally and during lactation. 14 We detected the presence of SW in both the amniotic fluid and in the milk, and this produced toxic effects in the offspring, such as reduction in the body weight, an increase in the spleen/body weight ratio, a decrease in the thymus/body weight ratio, and histological changes. Thus, the purpose of the present study was to evaluate whether SW exposure during gestation or lactation could produce immunotoxic effects that persist into adulthood in offspring of females exposed to I. carnea. Additional experiments were also performed in juvenile and adult rats to determine which immune system response (humoral or cellular) is affected by I. carnea and accounts for its immunomodulatory effects.
Materials and methods
Plant material
I. carnea leaves were collected in May 2004 from plants cultivated at the Research Center for Veterinary Toxicology (CEPTOX), University of São Paulo (USP), Pirassununga, Brazil. A voucher specimen was deposited in the Herbarium of the Instituto de Botânica de São Paulo, Brazil (SP-360911), and its authenticity was confirmed by the taxonomist Rosangela Simão Bianchini at the same institute.
The aqueous fraction (AF) resulting from the extraction of dry I. carnea leaves was obtained according to previously described methods. 13 Briefly, dry leaves were macerated in 96% ethanol. After total solvent evaporation under reduced pressure at 50°C, a dark green extract was obtained, which was suspended in water to remove the waxy residue and consecutively fractionated with n-butanol saturated with water. The remaining aqueous solution was lyophilized to give the AF, which, by a previous assay, revealed the presence of the active principles.
In our previous studies, a dose of 15 g/kg of AF administered by gavage was determined in our lab for the model of I. carnea toxicity in rats. 5,14 –16
Doses were delivered at a constant volume of 1 mL. All instillates were prepared fresh daily from a stock AF solution (15 g/mL in distilled water). Each instillate was generated by dilution of an aliquot of the stock AF with distillated water; the amount of diluents varied based upon the host body weight such that the final doses of 1, 3, 7, and 15 g AF/kg were achieved.
Animals
Male and pregnant and nursing female adult Wistar rats, 10 weeks of age, and juvenile male Wistar rats, 21 days of age, were obtained from our own colony in the Department of Pathology, School of Veterinary Medicine and Animal Science. All animals received food and water ad libitum and were maintained under controlled temperature (22–25 ° C), humidity (50–65%) and lighting (12/12 light/dark cycle) conditions. The experiments were carried out in accordance with the ethical principles for animal research adopted by the Bioethics Committee of the School of Veterinary Medicine and Animal Science, University of São Paulo.
Experimental design I—Perinatal treatment and offspring evaluation in adulthood
Twenty pregnant and 20 nursing rats were divided equally into eight groups: six experimental and two control groups. Pregnant rats in the experimental group were treated by gavage with 1, 3 and 7 g/kg of I. carnea AF from day 6 to day 20 of gestation. Nursing rats were treated with 3, 7 and 15 g/kg of I. carnea AF from postnatal day (PND) 1 to PND 21. Female rats in the control groups received only tap water by gavage. The dams’ total body weight gain and food consumption were measured during all the experimental periods. Where possible, after birth the litters were normalized to four females and four males. All offspring were individually weighed immediately after birth and daily until PND 21. On PND 3 and 21, three randomly-selected female pups from among the 20 (pooled from the 5 mothers in each treatment group) were euthanized and necropsied to obtain representative tissue samples for histopathological evaluation. The renal tissue was employed as a marker of I. carnea toxicity. The evaluation of immune parameters in adulthood (70 days old) was assessed only in males to avoid sexual hormonal interference.
Macrophage activation
On day 69 (PND), pooled rats (n = 5–8 from among the 20 male/20 females per group) from mothers treated with I. carnea AF during gestation or lactation were given an intraperitoneal (ip) injection of lipopolysaccharide (LPS; 1 mg/kg) to induce peritoneal inflammatory cell activation. The methods used to study macrophage spreading and phagocytosis were based on those described by Rabinovitch and DeStefano, 17,18 with modifications. Briefly, macrophages were obtained by lavage of the peritoneal cavity with 10 mL of cold PBS. The cells were centrifuged and resuspended in RPMI-1640 culture medium (RPMI) and adjusted to 2 × 106 cells/mL. Two hundred μL of each cell suspension was then placed in multi-well (six wells) tissue culture plates (at two wells/animal). To assess phagocytic activity, 1 mg of opsonized zymosan A solution (5.0 mg/mL) was added to each well and the cells then incubated for 60 min at 37 °C. After this period, the wells were rinsed with PBS to remove both un-ingested zymosan and any non-adherent cells. Thereafter, a total of 200 cells/well were analyzed and the macrophage spreading index (SI%) and phagocytic index (PI%) calculated as follows: SI or PI = number of spreading macrophages or phagocytic cells (PI, cells seen to have ingested ≥ 1 particle) × 100/200 adherent cells counted. Each well was examined twice; the mean of the four counted was obtained from the two wells/rat was then used to perform statistical analyses of SI and PI.
Spontaneous and phorbol myristate-acetate solution (PMA)-induced H2O2 release by macrophages was measured by the method of Russo et al. 19 H2O2 concentration was calculated from absorbance measurements as described by Pick and Mizel. 20
BCG-induced granulomatous inflammation
On day 70 (PND 70), granulomatous inflammation was induced by the injection of a constant volume (0.1 mL) of heat-inactivated oral onco-BCG (Instituto Butantan-Brazil, 4.0 × 107 bacillus/mL, Moreaux strain) intradermally into the left hind foot of offspring (n = 5–10) from dams treated with AF during gestation or lactation. The resulting edema was measured for 21 days by a water plethysmometer (Ugo Basile) at several time points: before BCG-injection, at 1, 2, 4, and 8 hours and then daily. Granulomatous inflammation is expressed as a volume of dislocated water and is the difference between the foot volumes of the first measurement and the subsequent measurements. Throughout the experimental period, some sick animals were euthanized and the left hind foot was stored in 10% buffered formalin, decalcified, embedded in paraffin, sectioned at a 5 μm thickness, and stained with HE.
Experimental design II—Comparison of the immunotoxic effects of I. carnea between young and adult rats
To evaluate the lymphoid organs’ relative weight, cellularity (immunopathology), and humoral and cellular immune responses, forty adult male rats (70 days old) and forty juvenile male rats (21 days old) were dosed with 0, 1, 3 or 7 g/kg of I. carnea AF by gavage for 14 consecutive days. Body weight gain and food consumption were measured every day. Instillate volume was maintained at 1 mL throughout. All instillates were prepared as described earlier to accommodate changes in host body weight over the 14-d period.
Immunopathology
At the end of the experimental period, the thymus and spleen were removed from euthanized rats in CO2 chamber and weighed. The spleen was broken up with two pieces of ground glass and the red blood cells were removed by lysing with 0.4% ammonium chloride sterile solution. This resulted in single splenocytes in suspension in cold RPMI-1640 culture medium. Bone marrow cell suspensions were achieved by flushing the marrow cavity of the left femur from each rat with ice-cold RPMI-1640 medium using a sterile syringe with a 26-gauge needle. Viability was assessed by means of a trypan blue dye exclusion test and cell numbers were determined by a hemocytometer.
Cellular immune response (delayed type hypersensitivity [DTH])
On day 15 (PND 15), rats were injected subcutaneously in the caudal tail fold with 0.1 mL of a 1 mg BSA/mL (Fraction V; Sigma) that was emulsified in Freund’s complete adjuvant (FCA; Sigma) at a ratio of 1:1. An immunological challenge was performed 7 days later with heat-aggregated BSA (hBSA), which was prepared by heating BSA (10 mg/mL) in saline to 75°C for 1 hour. After centrifugation and removing the excess saline, hBSA was injected intradermally into the left hind paw. Before and 24 hours after the hBSA injection, the footpad was measured with a caliper (Mytutoyo number 7309). Swelling was calculated by subtracting the value obtained after hBSA injection from value obtained before challenge.
Humoral immune response
After I. carnea AF treatment (i.e. on day 15), rats were immunized by ip injection with 2 × 109 sheep red blood cells (SRBC) in 0.9% saline. Seven days later, blood samples were harvested from euthanized rats, allowed to clot in vertical tubes at room temperature for 45 min and centrifuged at 4000 rpm for 10 min. The serum was collected and anti-SRBC antibody titers were estimated. Briefly, to each microtiter plate, 25 μL of diluted sera (2-fold dilutions with 0.9% saline) and 25 μL of SRBC suspension (1%, v/v) were added. The plates were then incubated at 37°C for 1 hour and then observed for hemagglutination. The highest dilution that still led to hemagglutination was taken as the “antibody titer”. All titers were expressed in a graded manner, with minimum dilution (1:2) ranked as “1”. The mean ranks of the different groups were then compared.
In addition, the plaque-forming cells assay (PFC) was performed to assess the humoral immune response in animals treated with the higher dose of I. carnea AF (7 g/kg). Briefly, splenocyte single-cell suspensions (1 × 107 cells/mL) were prepared in RPMI-1640 medium at 4°C. The proper amounts of splenocyte suspension, SRBC, and guinea pig complement were added to a 1 × 1 cm2 PFC well to achieve final concentrations of 2 × 106 splenocytes/mL, 7% SRBC, and 10% complement. The final volume in each PFC well was 50 μL. The PFC plate was covered with a glass slide (24 × 24 mm) and sealed with varnish. The PFC plate was incubated at 37°C for 3 hours. The number of lysate plaques produced by 1 × 105 splenocytes was counted under a light microscope (×40 magnification). The mean count obtained from two slides from each rat was used to express PFC/1 × 105 splenocytes.
Statistical analyses
Data were analyzed by one-way analysis of variance (ANOVA) followed by a post hoc analysis using Dunnett’s test. Differences between the control groups and the other groups treated with different doses of I. carnea were considered to be statistically significant when p < 0.05. Paired data were analyzed statistically by Student’s t-test, and the level of significance was set at p < 0.05. Data are reported as mean ± SEM.
Results
Maternal toxicity
Clinically, none of the dams from the experimental groups showed soft feces, depression, neurological signs or mortality during the study. However, two pregnant rats treated with 7 g/kg of I. carnea AF showed dystocia, and all of their litters died immediately after birth. There were no statistical differences in the total food consumption or total body weight gain between the pregnant rats and their respective controls. Nursing rats from the group treated with the highest dose of I. carnea AF (15 g/kg) consumed significantly less food when compared with untreated dams from the control group (p < 0.01); however, no changes in total body weight gain were noted between groups (Table 1).
Total food consumption and body weight gain of pregnant and nursing rats treated with Ipomoea carnea aqueous fraction
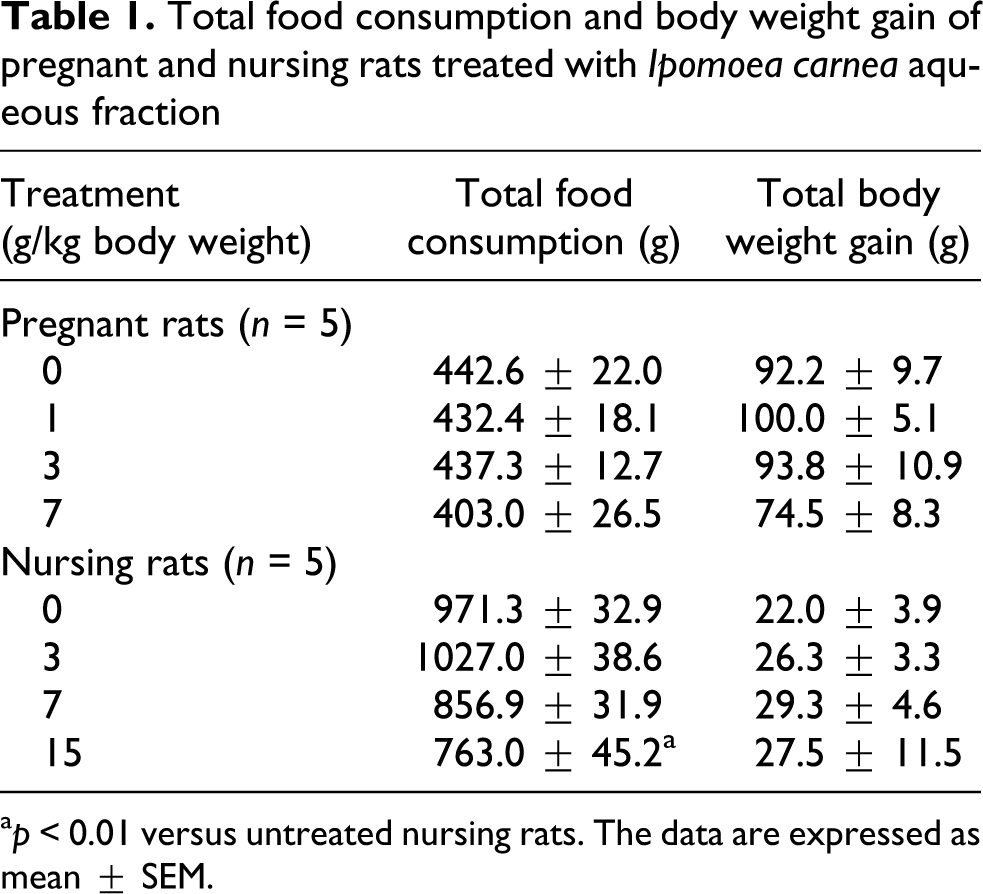
a p < 0.01 versus untreated nursing rats. The data are expressed as mean ± SEM.
Offspring toxicity
Table 2 shows pup body weights measured immediately after birth and the total body weight gain of these animals until weaning. These pups were from mothers treated with I. carnea during gestation or lactation. Litters from mothers treated with 7 g/kg of I. carnea AF during gestation showed a significant reduction in weight at parturition (p < 0.05) when compared to their respective control group; however, no differences in total body weight gain were observed between these litters after weaning. Litters from nursing dams treated with 7 or 15 g/kg of I. carnea AF showed decreases in total body weight gain during the lactation period when compared to litters from untreated nursing dams (p < 0.01).
Weight at parturition and total body weight gain of pups from dams treated with Ipomoea carnea aqueous fraction

a Number of pups immediately after birth.
b Number of litters per group—only males were employed.
c Two females lost their litters.
d p < 0.05 versus pups from untreated mothers. The data are expressed as mean ± SEM.
e p < 0.01 versus pups from untreated mothers. The data are expressed as mean ± SEM.
Histopathological evaluation of renal tissue revealed characteristic vacuolar lesions of I. carnea toxicity on PND 3 in pups from mothers treated with I. carnea during gestation (Figure 1); however, these lesions were absent when these pups were evaluated on PND 21. Histological analysis of pups from mothers treated with the extract of I. carnea during lactation did not reveal lesions in the renal tissue on PND 3; however, on PND 21 these pups showed marked cytoplasmic vacuolation in the convoluted tubular epithelium cells (Figure 1). In both histological studies, the vacuolar lesions occurred dose-dependently.

Tissue sections of pups from mothers treated with I. carnea during the perinatal period. (A) Kidney section of a pup on postnatal day (PND) 3 from dam treated with 7 g/kg of I. carnea aqueous fraction (AF) during gestation. (B) Kidney section of a pup on PND 21 from a mother treated with 15 g/kg of I. carnea AF during the lactation period. Note the marked vacuolar lesions on the convoluted epithelium cells in pups from both periods (HE, ×10)×.
Macrophage activity
In adult rats from dams treated with I. carnea during the gestation or lactation periods, the spreading and phagocytic properties and hydrogen peroxide production in the peritoneal macrophages were evaluated. Perinatal exposure to SW did not affect the activities of these macrophages in adulthood (data not shown).
BCG-induced granulomatous inflammation
The time course of paw edema formation was not affected by transplacental SW exposure or SW milk excretion following intradermal injection of BCG into rats from mothers treated during perinatal periods with I. carnea. The edema induced by BCG was evident three days after BCG injection and reached its maximum value after 13 days. This value remained stable until the end of the study with no statistical differences between the groups of rats from mothers treated with I. carnea during both perinatal periods (data not shown).
The clinical evaluation, however, revealed that many of the rats exposed to SW by the milk that were challenged with BCG in adulthood, independent of the dose administered, showed the following symptoms: pain when they were touched, locomotor difficulty, body weight loss, arthralgia and presence of edema throughout all joints. Following euthanasia of these animals, the histological evaluation of the edematous joints revealed the presence of joint damage resulting from inflammatory cell infiltration into the synovial cavity, cartilage and bone marrow degeneration (Figure 2).

Histological section of the left carpal joint of a rat from a mother treated with 3 g/kg of the I. carnea aqueous fraction during lactation and challenged with BCG in adulthood. (A) Note the presence of the hyperplastic synovial membrane comprising a layer of synovial fibroblasts and macrophages that invades adjacent cartilage and promotes marked joint damage. (B) The underlying bone marrow also exhibits an inflammatory infiltrate with necrosis and trabecular destruction (HE, ×10).
Ipomoea carnea toxicity and immunotoxicity in adult and juvenile rats
Table 3 displays the total food and body weight gain of the young and adult male rats treated with similar doses of I. carnea. These parameters did not change in juvenile rats at any dose. In contrast, adult rats treated with 3 and 7 g/kg of I. carnea AF consumed less food (p < 0.05) and gained less total body weight (p < 0.01) when compared to their respective controls.
Total food consumption and body weight gain of juvenile and adult male rats treated with Ipomoea carnea aqueous fraction

a,bNumber of 9 and 8 animals, respectively.
*p < 0.05
**p< 0.01 versus untreated rats. The data are expressed as mean ± SEM.
The effects of I. carnea on different lymphoid organ parameters in juvenile and adult rats were investigated. In young rats, the I. carnea AF (3 and 7 g/kg) promoted alterations in the thymus relative weight (p < 0.05); however, in adult rats, splenomegaly and a decrease in bone marrow cellularity were noted at these doses (Table 4).
Effects of Ipomoea carnea aqueous fraction on thymus and spleen relative weights and spleen and bone marrow cellularity of juvenile and adult male rats

a,bNumber of 9 and 8 animals, respectively.
*p < 0.05
**p < 0.01 versus untreated rats. The data are expressed as mean ± SEM.
Immune responses of juvenile and adult rats treated with I. carnea
Although no alterations were observed in the DTH response of juvenile and adult rats treated with different doses of I. carnea AF, an improvement in the humoral immune response after SRBC challenge was seen in both groups. As seen in Table 5, juvenile rats in all I. carnea AF-treated groups saw an increase in serum anti-SRBC titers when compared to their respective control group. However, only adult rats from the 3 and 7 g/kg groups showed the same increase in serum anti-SRBC titers. In the PFC assay, only juvenile rats (7 g/kg) showed an increase in in vitro SRBC lysis when compared to their control group.
Effect of Ipomoea carnea aqueous fraction on humoral and cellular immune response of juvenile and adult rats

a,bNumber of 8 and 9 animals, respectively.
*p < 0.05
**p < 0.01 versus untreated rats. The data are expressed as mean ± SEM.
Discussion
Implantation of the developing embryo into the maternal uterus wall and subsequent placental development is, immunologically, a critical stage in pregnancy. This is because the fetus has a hybrid-type genome with paternal antigens, which potentially could be detected as non-self by the maternal immune system. To solve this “immunological dilemma” in the peri-implantation period, many cytokines and hormones are released at the feto-maternal interface, so the maternal immune system can tolerate the trophoblastic invasion. 21 ,22 This tolerance process possesses a well-balanced cytokine network, and any immunological disturbances may cause implantation failure. 23 It is well known that SW produces immunomodulatory effects in different immune cells, such as NK cells 9 –11 and macrophages. 12,13,24 Therefore, we decided to begin the administration of I. carnea AF to pregnant rats on gestational day 6, the day of pregnancy when the embryo has already implanted into the maternal uterus. 25 This would prevent the possible immunomodulatory effects of SW on the inflammatory cells at the time of trophoblastic invasion.
Many studies, performed with different species of animals intoxicated with SW or plants that contain this alkaloid, revealed the occurrence of anorexia and, consequently, decreases in weight gain. 2,8,26 It is speculated that this anorexic effect could be due to CNS cell damage produced by the lysosomal vacuolar degeneration promoted by SW inhibition of α-mannosidase. 2 However, in previous experiments conducted in our laboratory, we administered up to 15 g/kg of I. carnea AF (containing approximately 2 mg/kg of SW) to rats and no morphological changes in neurons were noted. 14 Stegelmeier et al. 27 observed that only rats receiving high doses of SW (about 7 mg/kg/day) showed vacuolization in various neurons and glial cells.
Interestingly, in the present study, nursing rats treated with 15 g/kg of I. carnea AF showed a reduction in food consumption accompanied by an increase in body weight gain. Pups from nursing dams treated with I. carnea presented a decrease in body weight gain in a dose-dependent fashion, and these dams excrete SW into their milk during all period of lactation. 15 Therefore, it is possible that the increase in body weight gain in the dams was a result of a reduction in the suckling behavior of pups, which decreased the demand for milk production and, as a result, reduced their food consumption when compared to the control group. Indeed, in perinatal studies with I. carnea and other plants which have SW as toxic principle, it was verified that offspring of females (ewes and goats) exposed to these plants during gestation showed behavior abnormalities, including inability to suck.28–30
In this and a previous study, we confirmed that SW can be excreted into both the milk and amniotic fluid of females treated during lactation and gestation, respectively. 15 In the present study, we also determined that pups from nursing mothers treated with the AF lost weight during lactation, and histological changes (i.e. vacuolization) were noted on PND 21. Also, pups exposed to SW only in utero showed diminished body weight at birth but reached normal weights at weaning, and this suggests a recovery from SW toxicity, which agrees with the histological examination, when any alteration could be detected in the renal tissue of these pups on PND 21.
Many studies have been conducted on SW to better understand its different effects, such as protecting bone marrow against chemotherapeutic drug side effects,31–33 preventing metastasis, for use in cancer therapy9,34–36 and as an immune modulator in different cell lines,10,37,38 including macrophages that can be related to antineoplastic activity.12,24 A previous study conducted in our lab demonstrated that peritoneal macrophages from rats treated with I. carnea AF showed an increase in inflammatory activity and an improvement in phagocytosis and hydrogen peroxide production. 13
In the present study, we did not find an improvement in macrophage activity in the rats of mothers exposed to SW during gestation or lactation. Tissue macrophages have a maximum lifespan of several months, but they are not in a constant population because they are renewed regularly through the daily production of monocytes. 39 Moreover, mononuclear phagocytes show different kinetics during inflammatory processes, such as during acute inflammation where the number of circulating monocytes increases due to enhanced bone marrow production. 40 Thus, it is possible that we observed no improvement in macrophage activities because the normal macrophage turnover in the tissue and release from the bone marrow are not under SW influence.
It was also observed that, when offspring of female rats exposed to I. carnea during the perinatal period were challenged with BCG in adulthood, many animals showed arthralgia and edema in all joints. Histologically, the joint revealed a picture suggestive of rheumatoid arthritis (RA). Little is known about the pathogenesis of RA; however, it is believed that exposure of an individual that is genetically susceptible to an arthrogenic microbial antigen could lead to the development of the disease. 41 Indeed, it has been suggested that BCG-induced arthritis is related to a molecular mimicry mechanism in genetically susceptible individuals. 42 Nevertheless, it is known that the mechanism of SW action does not involve genetic disturbances; thus other mechanisms must be involved to predispose these animals to RA development.
In a pharmacokinetic study of SW in mice, Bowen et al. 43 observed that this alkaloid has a high affinity for the thymus and spleen. In our study, when I. carnea AF was administered to juvenile rats, an early thymic involution was observed. Thus, the presence of SW in the thymus may result in an alteration of the N-linked branched oligosaccharides on the glycoproteins due to Golgi mannosidase II inhibition. This would alter the molecular interactions between immature T cells and the thymic environment and interfere with the process of central tolerance. This interference could allow self-reactive T lymphocytes to escape and colonize secondary organs and facilitate their activation when an immunogenic challenge occurs. However, it should be noted that investigators have suggested that self-reactive T cell leakage to the periphery occurs more during fetal life 44 ; therefore, how does RA develop only in the rats exposed to SW during the lactation period? More studies should be conducted to better understand the mechanism of SW toxicity on the immune tolerance process.
The discrepancies in the results noted between juvenile and adults rats could be a consequence of the distinct physiology at each age. The decrease in thymus weight, in the young but not in the adult rat, could be because it did not possess its full functional activity and, therefore, was more prone to SW’s toxic effects. The decrease in bone marrow cellularity in the adult rats, but not in the younger rats, could be due to a low rate of proliferative activity in the adult bone marrow compared to the younger animals.
Despite differences in some parameters between the two age groups studied, both showed increases in the humoral immune response when treated with I. carnea AF. The antibody anti-SRBC production is a T cell-dependent antibody response. This means that the antibody production requires the interaction between T and B lymphocytes, and this is mainly through the co-stimulation of CD28 and CD80 molecules, respectively. This interaction is stronger in the presence of IL-2 produced by activated T cells. 45
To better understand the enhanced antitumor activities promoted by SW, in vitro studies have shown that this alkaloid can up-regulate the expression of the IL-2 receptor β-chain (IL-2Rb) in NK cells. 46 It is known that B cells express IL-2Rβ during their maturation process from naïve to memory B cell, 47 and IL-2 linkage can activate B cell proliferation and immunoglobulin synthesis. 45 However, no studies are available to verify the over expression of this IL-2Rβ on B cells exposed to SW. Moreover, the importance of CD28 and CD80 co-stimulatory signaling in antigen-specific T cell responses has been amply documented in studies of activation-induced T cell differentiation to memory, regulatory, or effector T cells. 48 Ma et al. 49 showed that treatment of CD28+ Jurkat cells (T cells) with inhibitors specific to N-glycosylation enzymes, such as SW, also enhanced binding with CD80 and resulted in increased IL-2 promoter activity. Taken together, we suggest that the SW contained in the I. carnea AF promotes an enhanced IL-2R expression on the B cells and an increased activation through the CD28/CD80 co-stimulatory interaction, which leads to an enhanced T cell-dependent antibody production in rats treated with I. carnea and high antibody titers.
In conclusion, administration of I. carnea AF to dams during the lactation period could promote development of self-reactivity in the offspring when an immunogenic challenge occurs in adulthood. Moreover, the juvenile and adult rats treated with I. carnea showed diverse toxic effects due to the physiological differences between the differently-aged rats; however, both age groups showed an enhanced humoral immune response, probably due to SW interference in glycoprotein synthesis.
Footnotes
This research was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP—proc. n.: 2001/11444-0), Brazil.