
Abstract
Progesterone (a neurosteroid) is an important modulator of the nervous system functioning. Organophosphorus pesticides like phosphamidon have been shown to adversely affect memory and induce oxidative stress on both acute and chronic exposure. The present study was therefore designed to investigate the effects of progesterone (PROG) on phosphamidon-induced modulation of cognitive function and oxidative stress in rats. Cognitive function was assessed using step-down latency (SDL) on a passive avoidance apparatus and transfer latency (TL) on an elevated plus maze. Oxidative stress was assessed by examining the levels of thiobarbituric acid reactive species (TBARS) and non-protein thiols (NP-SH) in isolated homogenized whole brain samples. The results showed a significant reduction in SDL and prolongation of TL in the phosphamidon (1.74 mg/kg/d; p.o.) treated group at weeks 6 and 8 as compared to the control group. Two weeks treatment with PROG (15 mg/kg/d; i.p.) antagonized the effect of phosphamidon on SDL as well as TL. Phosphamidon alone produced a significant increase in the brain TBARS levels and decrease in the brain NP-SH levels. Treatment with PROG (15 mg/kg/d; i.p.) attenuated the effect of phosphamidon on oxidative stress. Together, the results showed that progesterone attenuated the cognitive dysfunction and increased oxidative stress induced by phosphamidon in the brain.
Introduction
Neurosteroids (NS) are the steroids that are newly synthesized from cholesterol or another early precursor in the nervous system. They are present in nervous system even after removal of peripheral steroidogenic glands.1–3 NS occur in the nervous system as unconjugated steroids or sulfated or fatty acid esters of steroids. 4 These are involved in the control of metabolic and neurological processes including cognition, stress, anxiety and sleep, etc. Progesterone (PROG), pregnenolone (PREG), allopregnanolone (AP), pregnenolone sulfate (PREG-S), dehydroepiandrosterone (DHEA), dehydroepiandrosterone sulfate (DHEA-S), tetrahydrodeoxycorticosterone (THDOC) are some of the important NS.4–7 Besides their effect at transcriptional level through steroid receptors, NS may act through membrane receptors like gamma-aminobutyric acid (GABAA), nicotinic, muscarinic, N-methyl-D-aspartate (NMDA), sigma, kainate, glycine, serotonergic and neuropeptide receptors. They are also involved in neuroprotection, neurite outgrowth and synaptogenesis.8–15
Recently, a lot of emphasis has been laid on the role of NS in the functioning of the nervous system including cognition. NS have been implicated to have a role in neuroprotection and reinforcement of long-term memory in active avoidance behavior and learning. It has been seen that the levels of PREG-S, DHEA-S, PROG and AP are reduced in the brains of patients with Alzheimer’s disease. 16 In mice, immediate post-training intracerebroventricular administration of PREG, PREG-S, DHEA or DHEA-S has been correlated with improvement of memory retention in foot-shock active avoidance training. 17 PREG-S has been reported to facilitate memory and AP is involved in rewarding properties. Post-training memory processes are significantly enhanced by intra-amygdala injection of as little as 150 molecules of PREG-S in rats.18,19 Neuroprotective role of PROG has been demonstrated on learning and memory impairment and hippocampal damage in an earlier study 20 while AP, PREG-S, PROG and 4-CD have been found to reduce hypoxic stress in rats. 21 DHEA-S, DHEA, PREG, PREG-S and AP are also involved in inhibiting the neuronal apoptosis in hydrogen peroxide- and staurosporine-induced damage of human neuroblastoma, SH-SY5Y cells. 22 Further, neuroprotection after injury has been demonstrated in contusion of prefrontal cortex and spinal trauma with PROG and AP, respectively. 23 Recently, NS have also been found to reduce the exacerbations of seizures in catamenial epilepsy. 24
NS are also involved in the antioxidant defense mechanisms of the nervous system.25,26 The results of an earlier study has shown that PROG inhibits the lipid peroxidation as evidenced by the reduced levels of 8-isoprostaglandin F2 alpha in brains of PROG-treated rats in a cerebral contusion model. 27 In another study, DHEA pretreatment was found to reduce DNA damage induced by oxidative insult. 25
Phosphamidon, an organophosphate insecticide, has been shown to produce a number of adverse effects, of which the most prominent is neurotoxicity. It has been associated with a decrease in memory and hand-eye co-ordination. The damage caused to the nervous system has been linked with inhibition of the activities of acetylcholinesterase and GABA.28,29 Phosphamidon has also been found to result in increased oxidative stress and suppression of humoral immune response in rats in some previous studies.30,31 It causes lipid peroxidation by generating oxygen free radicals and superoxide ions. Superoxide dismutase, catalase (anti-oxidant enzymes) and glutathione levels are also altered following exposure to phosphamidon.30,31
The present study was therefore designed to investigate the effects of PROG on phosphamidon-induced modulation of cognitive function and oxidative stress in rats.
Materials and methods
Animals
Male Wistar rats weighing between 150 and 220 g, obtained from the Central Animal House of the University College of Medical Sciences and GTB Hospital, were used. The animals were housed in polypropylene cages (30 × 15 × 15 cm) in groups of five rats per cage with free access to pellet diet and water, and kept under controlled environmental condition (temperature: 22 ± 2°C, humidity: 50%−55%, natural light/day cycle). All the experiments were performed during the light phase between 0930 and 1530 hours. The animals were taken care of in accordance with the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), India. The study was duly approved by the Institutional Animal Ethics Committee, University College of Medical Sciences, Delhi.
Drugs and dosing schedule
Phosphamidon and PROG were obtained from Sigma chemicals (Kansas, Missouri, USA). All other chemicals used were of laboratory grade. Phosphamidon was given in a dose of 1.74 mg/kg/d per orally (p.o.) with distilled water as a vehicle. PROG was administered intraperitoneally (i.p.) in a dose of 15 mg/kg/d after dissolving in distilled water with two drops of Tween-80 added per 10 mL of suspension. Each animal received 0.5 mL of suspension per 100 g of weight per day. Animals were randomly divided into four groups having 10 rats per group. All animals received phosphamidon or vehicle for phosphamidon (p.o.) for 6 weeks. PROG and its vehicle were administered i.p. for 2 weeks after treatment with phosphamidon (Table 1). The different groups were evaluated for cognitive function one day before the start of treatment, at six weeks and on the day of completion of treatment schedule. Animals were trained on each day prior to assessment of cognition. Animals were tested after 30 min for acquisition (training) and after 24 hours for retention (testing for retention of the learned task) of memory. Finally, at the end of study period, the animals were sacrificed using deep ether anaesthesia and brain taken out to assess oxidative stress.
Animal groups and their respective treatments

Abbreviations: PROG: progesterone, i.p.: intraperitoneal.
Assessment of cognition
Cognition was assessed on the basis of two separate experiments:– Step down latency (SDL) in passive avoidance apparatus. The apparatus consisted of a grid floor, on the center of which a wooden block of height 4 cm was placed. The block served as a shock-free zone (SFZ). The rat was placed on the SFZ and on stepping down was given electric shock (20V, 50 Hz, 1 mA, 1 sec) through the grid floor. The experiment was repeated after half hour and the time taken by the rat to step down was observed (SDL; acquisition of memory). The procedure was repeated after 24 hour without shock (retention of memory). A cut-off time of 180 sec was taken and for the animal which did not step down during this period, SDL was taken as 180 sec.32,33
Transfer latency (TL) on elevated plus maze. The elevated plus maze consisting of two open arms (50 × 10 cm) and two closed arms (50 × 10 × 40 cm) with an open roof was used. The maze was elevated to a height of 50 cm from the floor. The animals were placed individually at either ends of the open arms and allowed to enter the closed arms. During training, if the animal did not enter the closed arm within 180 sec, it was gently pushed in the closed arm. To become acquainted with the maze, the animals were allowed to explore the maze for 20 sec after reaching the closed arm and then returned to their home cage. The learning was tested 30 min later on the same day and the animals were retested 24 hours after the first day training to test the retention of memory. The time which the animal takes to move from the open arm to the closed arm was taken as TL. A time of 180 sec was taken as cut-off and animals which did not enter the closed arm during this period were assigned the TL of 180 sec.34,35
Assessment of oxidative stress
At the end of study period, animals were sacrificed by deep ether anesthesia. Whole brain was quickly dissected out, washed with ice-cold sodium phosphate buffer, weighed and stored over ice. The brains were further processed within half an hour of dissection and the estimation of oxidative stress was done on the same working day. Brain tissue was homogenized with 10% sodium phosphate buffer (7.4 pH, ice cold, mixture of KH2PO4 and Na2HPO4). The homogenate was centrifuged at 9170 g for 15 min and the supernatant was used for estimation of thiobarbituric acid reactive species (TBARS) and non-protein thiol (NP-SH) levels. The TBARS, an indicator of lipid peroxidation was estimated as described by Okhawa et al. 36 NP-SHs were estimated by the method as described by Ellman. 37 This assay estimates the activity of glutathione reductase that reduces glutathione disulfide to the sulfhydryl form GSH, which is an important cellular antioxidant.
Statistical analysis
All the values are expressed as mean ± standard error of the mean (SEM). Data were analyzed using one way ANOVA with post-hoc Tukey test (α = 0.05).
Results
Cognitive assessment parameters
Step-down latency
At the beginning of the experiment (day 0), no significant differences were found among the SDL values of all the experimental groups. A significant reduction in both acquisition as well as retention in SDL paradigm was observed for the phosphamidon-treated group at week 6 and 8 as compared to SDL values of both control (p < 0.001) and day 0 of phosphamidon-treated groups (p < 0.001; Figure 1A and B). A significant reduction in SDL values was also noted for phosphamidon + PROG group at week 6 (p < 0.001). After two weeks treatment (i.e. at the end of 8th week), PROG was observed to reverse the effect of phosphamidon on SDL in both acquisition as well as retention paradigms (Figure 1A and B).

Effect of phosphamidon and progesterone (PROG) on (A) acquisition on step-down latency (SDL); (B) retention on SDL. All animals received phosphamidon or vehicle for phosphamidon (1.74 mg/kg/d, p.o.) for 6 weeks. PROG or its vehicle was administered i.p. for 2 weeks after treatment with phosphamidon in a dose of 15 mg/kg/d. The different groups were evaluated for cognitive function 1 day before the start of treatment, at 6 weeks and on the day of completion of treatment schedule. Animals were trained on each day prior to assessment of cognition. Animals were tested after 30 min for acquisition and after 24 h for retention of memory. Values are expressed as mean ± SEM, for 10 animals in each group. a p < 0.001 as compared to normal control group; b p < 0.001 as compared to phosphamidon-treated group.
Transfer latency At day 0, no significant difference was found among the transfer latency values of all the groups. In the phosphamidon-treated group, a significant prolongation in both acquisition and retention was seen as compared to the control group (p < 0.001). Same effect was also noted in the group given phosphamidon + PROG, at 6th week as compared to the control and PROG groups, respectively. There was marked reduction in the TL of phosphamidon + PROG group (p < 0.001) as compared to the phosphamidon only group at 8th week (Figure 2A and B).
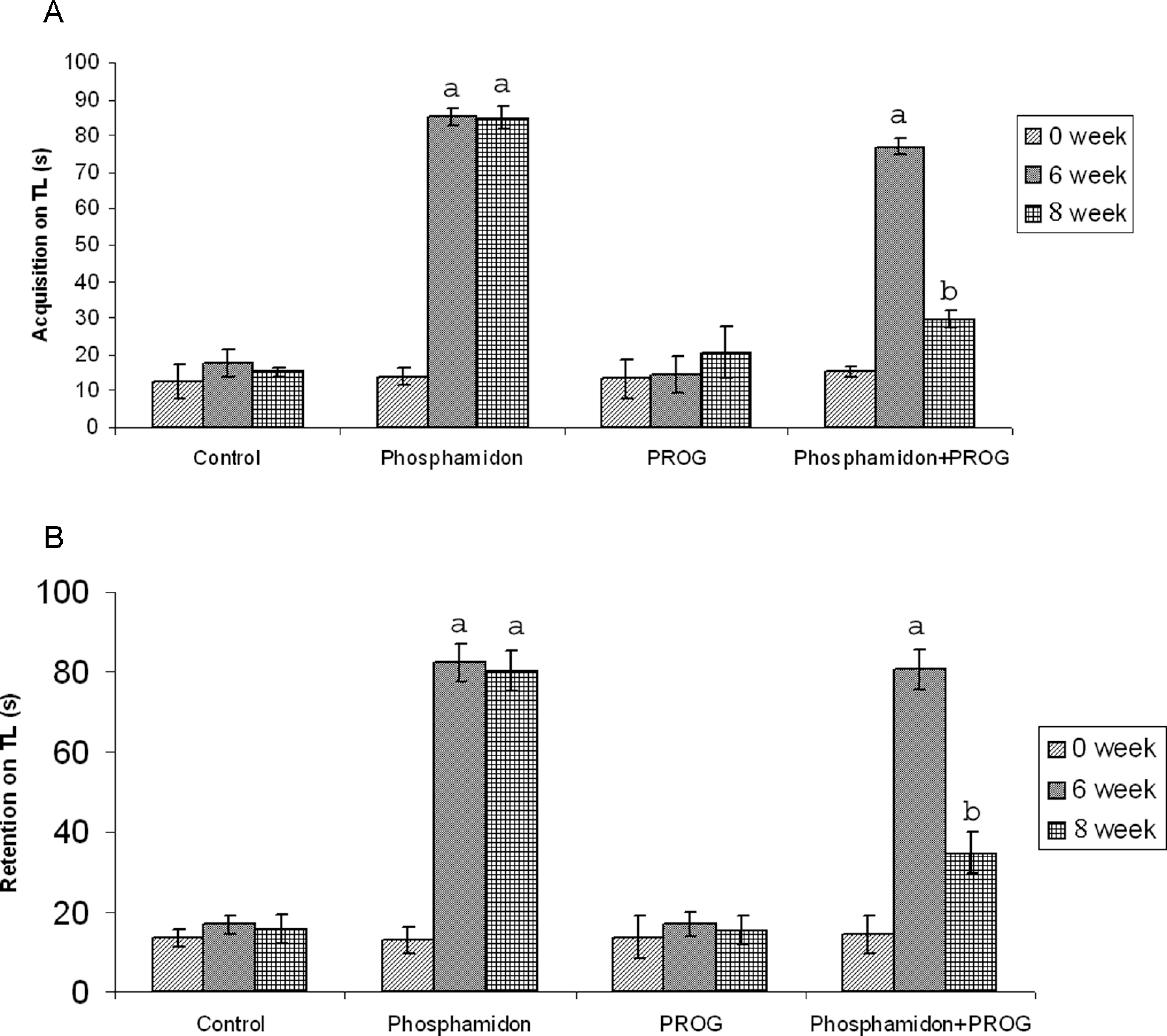
Effect of phosphamidon and progesterone (PROG) on (A) acquisition on transfer latency (TL); (B) retention on TL. All animals received phosphamidon or vehicle for phosphamidon (1.74 mg/kg/d, p.o.) for 6 weeks. PROG or its vehicle was administered i.p. for 2 weeks after treatment with phosphamidon in a dose of 15 mg/kg/d. The different groups were evaluated for cognitive function 1 day before the start of treatment, at 6 weeks and on the day of completion of treatment schedule. Animals were trained on each day prior to assessment of cognition. Animals were tested after 30 min for acquisition and after 24 hours for retention of memory. Values are expressed as mean ± SEM, for 10 animals in each group. a p < 0.001 as compared to normal control group; b p < 0.001 as compared to phosphamidon-treated group.
Assessment of oxidative stress
TBARS estimation
There was a significant increase in the brain TBARS levels of group treated with phosphamidon only (p < 0.001). Treatment with PROG attenuated the effect of phosphamidon on TBARS level with the difference between phosphamidon group and phosphamidon + PROG group found to be significant (p < 0.001). No significant difference was observed in the brain TBARS levels of PROG group only as compared to the control group (Table 2).
Effect of phosphamidon and progesterone (PROG) on parameters of thiobarbituric acid reactive species(TBARS) and non-protein thiols (NP-SH) in rats (n = 8) a

Abbreviation: i.p.: intraperitoneal, p.o.: per oral.
a The data has been expressed as mean ± SEM.
b p < 0.001 as compared to control group.
c p < 0.001 as compared to phosphamidon-treated group.
Non protein thiols
A significant decrease was found in the brain NP-SH levels of phosphamidon-treated group as compared to the control group (p < 0.001). A significant increase was noted for PROG only (p < 0.001), and phosphamidon + PROG (p < 0.001)-treated groups, respectively, as compared to the control group. The difference between phosphamidon alone and phosphamidon + PROG groups (p < 0.001) was found to be statistically significant (p < 0.001; Table 2).
Discussion
The present study was designed to explore the effect of PROG on phosphamidon-induced cognitive dysfunction by measuring the SDL in passive avoidance paradigm and TL in the elevated plus-maze apparatus. Since oxidative stress has been found to affect memory and both NS and phosphamidon have influence on the oxidative stress, parameters of oxidative stress viz. TBARS and NP-SH were also estimated in the brain at the end of the study period.
In the present study, PROG showed attenuating effect on phosphamidon-induced cognitive dysfunction. There are reports in literature regarding the influence of PROG on cognitive function. Progesterone has been shown to have a beneficial effect on episodic memory and verbal learning in post-menopausal women on hormone replacement therapy.38,39 Other workers have demonstrated a decrease in cell death and cognitive deficits after experimental contusion to rat pre-frontal cortex by AP and PROG when used 1 day after injury. 40 In contrast to above observations, in a recent study, PROG reversed the beneficial cognitive effects of estradiol over spatial performance in ovarectomized rats. 41 Some studies failed to demonstrate the improvement of memory in experimental animals 42 and post-menopausal women by PROG. 43 It is now well established that gonadal hormones like PROG can affect memory. Further, the performance in cognitive parameters can be influenced by sex as well as the phase of estrous cycle in female animals.44,45 PROG is an intermediate substance in the biosynthesis of other NS. We chose PROG over AP and DHEA on the basis of our hypothesis that besides its direct effect on cognition, PROG should also enhance the biosynthesis of other NS leading to a more convincing effect. Further studies are required to confirm these findings.
Phosphamidon resulted in induction of oxidative stress in brain as evidenced by reduction in brain NP-SH activity and increase in brain TBARS levels. Lipid peroxidation has been postulated as the primary event mediating the toxicity of a broad spectrum of pesticides. Other works have shown that in vitro activities of delta-amino levulinic acid dehydratase and Cu-Zn SOD in human red blood cells were inhibited after methamidophos exposure. 46 In pesticide poisoning cases, tissue glutathione reductase (GR), glutathione peroxidase, SOD and CAT activities, as well as malondialdehyde production are increased but GSH levels are decreased suggesting adaptive measure to overcome any insecticide accumulation. 47 These enzymes efficiently scavenge toxic free radicals and partly protect against lipid peroxidation from acute / chronic pesticide exposure. The generation of oxidative stress by phosphamidon in the present study is in strong accordance with some previous studies. 48
NP-SH is the most prevalent and important intracellular antioxidant. This compound is able to scavenge both singlet oxygen and hydroxyl radicals. Levels of NP-SH were observed to decrease in the PHOS-treated group as compared to the control group. PROG significantly reversed this effect as evidenced from an increase in the levels of NP-SH.
Besides their effect on cognition, NS like PROG have been previously demonstrated to have effect on the metabolic profile, behavior and other body systems.49,50 These possible effects were not parameters of this study, hence were not explored.
Oxidative stress has been closely related to cognitive dysfunction. Oxidative stress has been implicated in the pathogenesis of Alzheimer’s disease in humans as well as in several animal models of Alzheimer’s disease.51,52 At the same time, the mechanism of influence of phosphamidon over cognition and behavior is not solely limited to oxidative stress. It has been postulated that phosphamidon achieves its behavioral effects and neurotoxicity through several mechanisms like inhibiting acetylcholinesterase, interfering with GABA, affecting energy supply, glucose utilization and SH-groups in neurons. 53 In the present study, phosphamidon adversely affected the parameters of memory and oxidative stress. PROG demonstrated favorable effects on these parameters. The results of the present study provided evidence that SDL was reduced and TL prolonged on long-term (6 week) administration of phosphamidon. PROG was able to reverse the impairment in memory caused by phosphamidon when administered for 2 weeks following pretreatment with phosphamidon for 6 weeks. Furthermore, phosphamidon alone increased the oxidative stress while PROG reversed the phosphamidon-induced increase in the TBARS levels. This study reveals a possible correlation between memory impairment and oxidative stress in brain, signifying yet another potential role of NS in the functioning of nervous system and possible use of this group of drugs in reversing the damage induced by pesticides like phosphamidon in the brain.
Footnotes
This research received no specific grant from any funding agency in the public, commercial, or not for-profit sectors.