
Abstract
Loxosceles venoms can promote severe local and systemic damages. We have previously reported that Loxosceles gaucho spider venom causes a severe early thrombocytopenia in rabbits. Herein, we investigated the in vitro effects of this venom and its sphingomyelinase fraction on the main functions of platelets. Whole venom and its fraction induced aggregation of both human and rabbit platelets. Aggregation was dependent of plasma component(s) but independent of venom-induced lysophosphatidic acid generation. There was no increase in the levels of lactate dehydrogenase during platelet aggregation, ruling out the possibility of platelet lysis. The increased expression of ligand-induced binding site 1 (LIBS1) induced by L. gaucho venom and its sphingomyelinase fraction, as well as of P-selectin by the whole venom, evidenced the activation state of both human and rabbit platelets. Adhesion assays showed an irregular response when platelets were exposed to the whole venom, whereas the sphingomyelinase fraction induced a dose-dependent increase in the platelet adhesion to collagen. These findings evidence that L. gaucho venom and its sphingomyelinase fraction trigger adhesion, activation, and aggregation of both human and rabbit platelets. Thus, this work justifies the use of rabbits to investigate Loxosceles venom-induced platelet disturbances, and it also supports research on the role of platelets in the pathogenesis of loxoscelism.
Introduction
Spiders belonging to the genus Loxosceles, popularly known as brown spiders, are widespread from equatorial to subtemperate regions of the world. 1 Envenomation by Loxosceles sp may result in a necrotizing-hemolytic syndrome, loxoscelism, which is considered the most important form of araneism in South America, where the main species of medical importance are L. laeta, L. intermedia and L. gaucho. 2
The hallmark of the loxoscelism is the development of a dermonecrotic wound at the bite site, which is histologically characterized by edema, extensive intradermal and subcutaneous hemorrhage, polymorphonuclear infiltration, thrombus and extravascular fibrin deposition that progress to an ulterior aspect of fibrinoid necrosis.1,2,3 Although less frequent, human accidents may evoke severe systemic disturbances, such as intravascular hemolysis and disseminated intravascular coagulation, which may result in renal failure and death.4,5 We have previously verified that rabbits injected intradermally with L. gaucho whole venom develop severe early thrombocytopenia, but this phenomenon is not related to systemic loxoscelism since intravascular hemolysis and disseminated intravascular coagulation could not be verified in this experimental model. 6
The sphingomyelinase activity of 30-35 kDa proteins from Loxosceles sp venom has been described as the main mechanism in the induction of dermonecrosis, and the mediation of complex and multifactorial processes, such as complement activation and interactions between venom components and the cell membrane and/or extracellular matrix.7–10 Among other effects, Loxosceles venoms and their toxins exhibiting sphingomyelinase activity induce platelet aggregation in vitro.11–14 However, it is unknown whether it could be the result of either agglutination or platelet lysis. In addition, the effects of Loxosceles venoms on other platelet functions, such as adhesion and activation, have not been studied yet.
Thus, in the present study, we investigated the in vitro actions of L. gaucho whole venom and its sphingomyelinase fraction on the main functions of platelets: activation, adhesion and aggregation. Moreover, in order to extrapolate experimental studies in animals to the human envenomation, and taking into account that rabbit is the most used experimental animal model to investigate loxoscelism, we also compared the effects of L. gaucho venom and its sphingomyelinase fraction on both human and rabbit platelets.
Material and methods
Loxosceles gaucho venom and its sphingomyelinase35-kDa fraction
Specimens of L. gaucho were collected in São Paulo State, Brazil. After electrostimulation of the cephalotorax region, spider venom extraction was performed by micropipette aspiration. The venom extracted of hundred of spiders was pooled and frozen until the moment of use, as previously described. 15
The protein content was determined by the bicinchoninic acid assay, 16 using bovine serum albumin (BSA) (Sigma-Aldrich, USA) as standard. The studied fraction from L. gaucho venom was purified using a Mono-S column, as reported by Cunha et al. 17 This fraction showed a single band of 35 kDa by SDS-PAGE, and it was able to induce dermonecrosis in rabbits and exhibited sphingomyelinase activity assayed as described previously. 7
Platelet aggregation assay
Human blood was withdrawn from the brachial vein of healthy volunteers and then mixed with 3.8% trisodium citrate (9:1, v/v). Only donors declaring that they had not used any drugs known to interfere with platelet function for at least 14 days earlier were included in this study. Rabbit blood was withdrawn from the auricular central artery through a butterfly scalp needle (21 g) and then added to 3.8% trisodium citrate (9:1, v/v). For each experiment using human or rabbit blood, a new collection of blood was carried out. The procedures involving animals were performed in accordance with the Guide for the Care and Use of Laboratory Animals (1996; http://www.nap.edu/readingroom/books/labrats/) and they were approved by the Ethical Committee for the Use of Animals at Butantan Institute (process number 129/2003).
Platelet-rich plasma (PRP) or washed platelets were obtained from fresh blood samples of both species as described previously. 18 Briefly, PRP was obtained after blood centrifugation (190 g) for 20 min at room temperature. Washed platelets were obtained from PRP through sequential washings in Tyrode's solution containing prostaglandin PGE1 (Sigma-Aldrich, USA) and subsequent centrifugations (1900 g/15 min) at room temperature. Final platelet count in PRP or washed platelets was adjusted to 300 × 109/L. Platelet aggregation was performed in a chrono-log lumiaggregometer (model 560VS) by the method of Born 19 and results were expressed in percentage of change in light transmittance.
Whole venom or fraction was diluted in saline solution, and then 10 µL of whole venom (20 or 40 µg/mL for human or rabbit platelets, respectively, final concentration) or sphingomyelinase fraction (2 or 4 µg/mL for human or rabbit platelets, respectively, final concentration) were added to 400 µL of PRP. The intensity of platelet aggregation was measured for 10 min. The positive control consisted of human or rabbit PRP incubated with collagen (Chrono-log, USA) in a final concentration of 2 or 4 µg/mL, respectively.
As it is possible that venom sphingomyelinases act on lysophasphatidylcholines (LPC) to generate lysophosphatidic acid (LPA), which is known to mediate platelet activation through the stimulation of the LPA1 and LPA3 receptors, 20 the participation of the LPA pathway in the platelet aggregation induced by L. gaucho venom was investigated. Human and rabbit PRP were incubated with both venom (in the same concentrations described above) and dioctanoylglycerol pyrophosphate (DGPP 8:0; Avanti Polar Lipids, USA), a selective antagonist of the LPA1 and LPA3 receptors, 20 in the concentrations of 20 to 40 mM; 1-acyl(20:4) glycerophosphate (LPA 20:4; Avanti Polar Lipids, USA) was used as a positive control (concentrations of 20 to 40 mM) to induce platelet aggregation.
Lactate dehydrogenase leakage
In order to verify platelet lysis induced by the venom, PRP was incubated with L. gaucho whole venom as described above. After aggregation, it was centrifuged at 1900 g for 15 min. The supernatant was used to determine the leakage of lactate dehydrogenase (LDH) from platelets, according to a modification of the method described by Burtis and Ashwood. 21
Flow cytometry of platelets
In order to determine the expression of activation markers on platelet membrane, human or rabbit PRP incubated with venom or its fraction was used for flow cytometry studies. Briefly, 1 volume of 2% paraformaldehyde in phosphate buffered saline (PBS, pH 7.4) was added to 1 volume of PRP and then incubated with antibody-saturated solution. The labeling intensity of fluorescein isothiocyanate-conjugated (FITC-conjugated) or phycoerythrin-conjugated (PE-conjugated) antibodies was evaluated in PRP exposed to L. gaucho venom or its fraction and compared with quiescent or collagen-activated platelets. The following monoclonal antibodies were employed: FITC-conjugated anti-rabbit GPIIb-IIIa IgY, which reacts with both rabbit and human platelets 22 ; anti-ligand-induced binding site (anti-LIBS1; kind donation of Dr Mark H Ginsberg, Scripps Research Institute, La Jolla, USA), an antibody that recognizes the ligand-induced binding site 1 (LIBS1) of human GPIIIa following occupation of GPIIb-IIIa by either fibrinogen or peptides that contain the sequence RGD or HHLGGAKQAGDV, 23 and that also cross reacts with rabbit GPIIIa 22 ; 12A7 (kind donation of Dr Guy Reed, Harvard School of Public Health, Boston, USA), an antibody that specifically reacts with rabbit platelet P-selectin 24 ; PE-conjugated anti-human CD62P (Pharmingen, USA), that reacts specifically with human P-selectin.
Platelets were exposed, under the aggregation conditions described previously, to venom, its fraction or collagen (positive control) for 10 min. Quiescent platelets (negative control) remained under agitation for the same time at 37°C. Thereafter, each sample was fixed with an equal volume of 2% paraformaldehyde in PBS. Aliquots of fixed platelet suspension (25 µL) were incubated at room temperature in the dark with: (1) anti-LIBS1 (25 µL, 40 µg/mL) or (2) 12A7 (25 µL, 20 µg/mL) for 1 hour, and then 2 µL of PE-conjugated goat anti-mouse IgG (Sigma-Aldrich, USA) was added and incubated for 1 hour; (3) PE-conjugated anti-human CD62P (2 µL, 100 µg/mL) for 1 hour; (4) FITC-conjugated anti-rabbit GPIIb-IIIa IgY (2 µL, 500 µg/mL) for 2 hours. Samples were analyzed on a FACSCalibur flow cytometer (Becton Dickinson, USA). Light scatter and fluorescence data from 10 000 events were collected with all detectors in logarithmic mode. Data were saved in flow cytometry list mode files for subsequent analysis in WinMDI software, version 2.8.
Platelet adhesion assay
Platelet adhesion to collagen was performed in microplates using a modification of a previously described method. 25 Microplates were coated with 50 µL/well of 40 µg/mL equine collagen Type I (Chrono-log, USA), diluted in 5 mM acetic acid for 2 hours at 37°C. Microplates were then washed two times by plate inversion with PBS containing 2 mM MgCl2 (modified PBS). Blocking was carried out with 2 mg/mL BSA for 1 hour at 37°C. After washing with modified PBS, 40 µL/well of PRP (adjusted to 75 × 109/L and 150 × 109/L platelets for human and rabbit PRP, respectively) were added to prewarmed microplates at 37°C, and afterwards 10 µL of two-fold serial dilutions of venom or its fraction were added. Plates were incubated for 1 hour under static conditions in a humidified thermostat at 37°C. At the end of the incubation, plates were washed with PBS and wells were rapidly supplemented with 150 µL of 0.1 M citrate buffer, pH 5.4, containing 0.1% Triton X-100 in order to cause instantaneous lysis of platelets, and 5 mM p-nitrophenyl phosphate (Sigma-Aldrich, USA) as substrate for platelet phosphatases. After incubation at 37°C for 1 hour, the reaction was stopped and the color was developed by the addition of 100 µL of 2 M NaOH. The p-nitrophenol produced by the reaction was measured with a microplate reader at 405 nm against a blank, a platelet-poor plasma (PPP). The percentage of adherent cells was calculated on the basis of a standard curve obtained with known platelet counts in PRP preparations.
Statistical analysis
Two-way ANOVA and Tukey test were used to analyze data and compare the results among the different groups. Statistical analyses were carried out on STATATM software version 5.0 and SigmaStat software version 2.03 (SSPS, USA). Differences with p < 0.05 were considered statistically significant.
Results
The whole venom of L. gaucho spider and its sphingomyelinase fraction were able to induce platelet aggregation when they were exposed to either human or rabbit PRP (Figure 1 ). The intensity of this response was more evident when human PRP was used, since rabbit platelets required a two-fold higher concentration of venom to reach a similar extent of aggregation. When using washed platelets of both species (without the presence of plasma components), aggregation was not observed in the presence of either whole venom or sphingomyelinase fraction.

Platelet aggregation induced by L. gaucho venom and sphingomyelinase fraction in rabbit and human platelet-rich plasma (PRP). Human PRP was incubated with 2 μg/mL of collagen (positive control), 20 μg/mL of whole venom or 2 μg/mL of sphingomyelinase fraction. Rabbit PRP was incubated with 4 μg/mL of collagen (positive control), 40 μg /mL of whole venom or 4 μg/mL of sphingomyelinase fraction. Data are expressed as mean ± SEM (n = 5).
Dioctanoylglycerol pyrophosphate was able to inhibit aggregation induced by LPA 20:4, but not that induced by L. gaucho whole venom (data not shown). Increase in the levels of LDH during platelet aggregation of either rabbit or human PRP induced by the whole venom under conditions of controlled stirring and temperature was not observed (Figure 2 ). Since there was no response to the whole venom, the sphingomyelinase fraction was not tested in LPA and LDH experiments.

Effect of the incubation of rabbit and human platelet-rich plasma (PRP) with 20 μg/mL and 40 μg/mL L. gaucho whole venom, respectively, on lactate dehydrogenase (LDH) leakage. Basal levels were assayed in platelet-poor plasma (PPP) of both species. Data are expressed as mean ± SEM (n = 5).
The study of platelet membrane activation markers by flow cytometry (Figure 3 ) showed a significant increase (p < 0.05) in LIBS1 expression in both human and rabbit platelets exposed to the whole venom and its sphingomyelinase fraction. On the other hand, P-selectin expression was increased in human and rabbit platelets by exposition to the whole venom, but not to the sphingomyelinase fraction. Finally, GPIIb-IIIa expression in platelets of both species was not modified by either the whole venom or its fraction.

Expression of platelet membrane activation markers: ligand-induced binding site (LIBS1); P-selectin; GPIIb-IIIa. They were analyzed by flow cytometry and expressed in arbitrary units of fluorescence (AUF), on rabbit and human platelets incubated with L. gaucho whole venom and sphingomyelinase fraction. Human platelet-rich plasma (PRP) was incubated with 2 μg/mL of collagen (positive control), 20 μg/mL of whole venom or 2 μg/mL of sphingomyelinase fraction. Rabbit PRP was incubated with 4 μg/mL of collagen (positive control), 40 μg/mL of whole venom or 4 μg/mL of its fraction. Quiescent platelets were used as negative control for both species. Data are expressed as geometric mean ± SEM (n = 5). *p < 0.05 vs negative control.
Both human and rabbit platelets adhered with high affinity to microplates coated with collagen in a platelet count-dependent manner. Platelet counts of 150 × 109/L and 75 × 109/L for rabbit and human PRP, respectively, corresponded to the optimum count to be tested for platelet adhesion in the presence of L. gaucho whole venom or its fraction. The choice of these platelet counts let us obtain more homogeneous results for concentration-response curves. In the presence of the sphingomyelinase fraction, but not L. gaucho whole venom, platelet adhesion from both rabbit and human PRP responded in a concentration-dependent manner (Figure 4 ); the percentage increase in platelet adhesion as a function of the sphingomyelinase concentration was more intense in rabbit PRP than in human PRP. In spite of an irregular response caused by the whole venom, platelet adhesion was more intense when using human PRP than rabbit PRP (Figure 4).
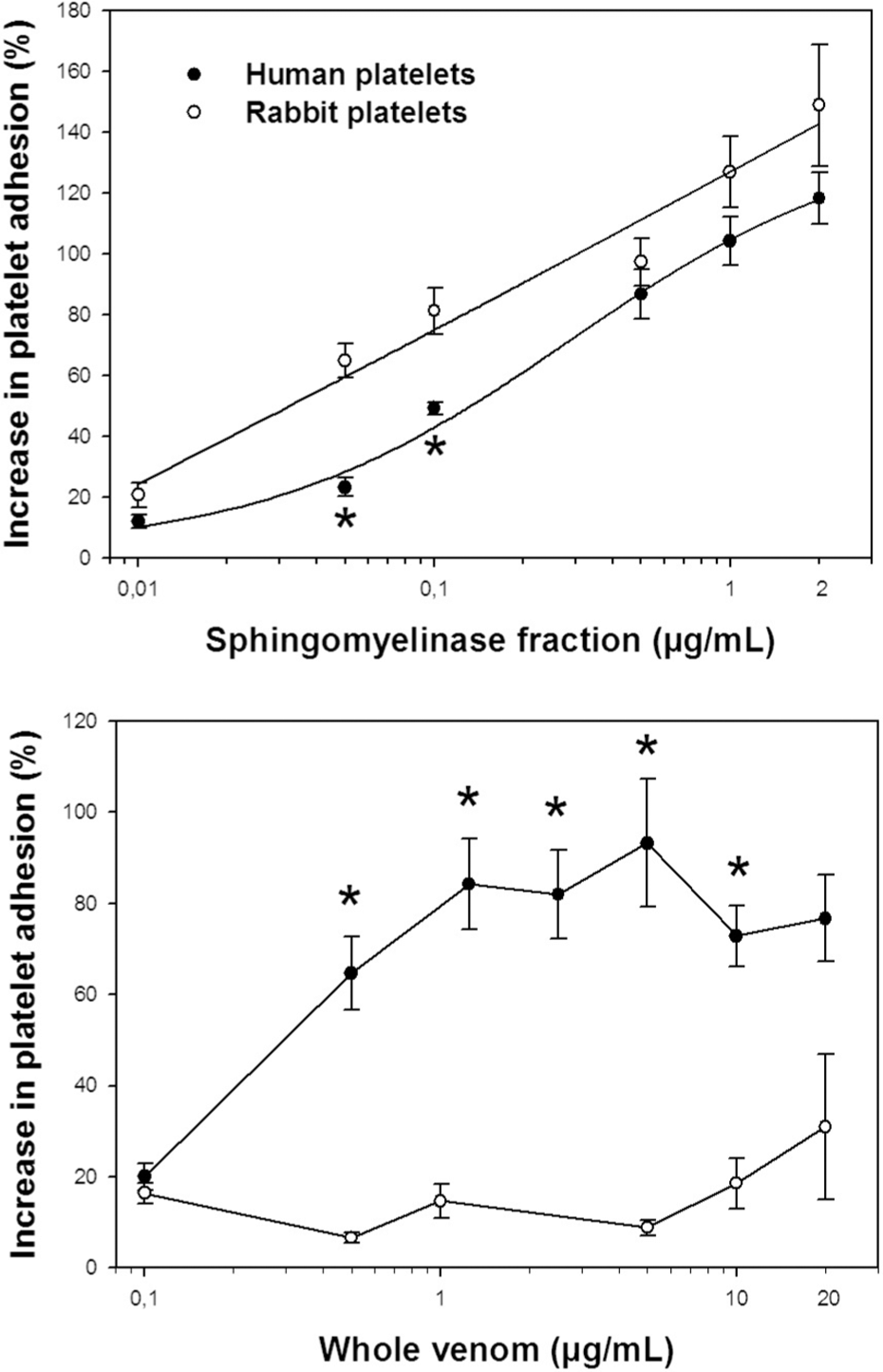
Platelet adhesion of rabbit and human platelet-rich plasma (PRP) exposed to: L. gaucho whole venom; sphingomyelinase fraction. Concentration-response curves were analyzed using a fixed platelets count (rabbit: 150 × 109/L; human: 75 × 109/L) and varying concentrations of whole venom and its fraction. Data are expressed as mean ± SEM (n = 4). *p < 0.05 vs rabbit platelets.
Discussion
Loxoscelism is a necrotizing pathological process, which may cause systemic hemolytic and coagulation disorders. Loxosceles venoms and their toxins affect a wide spectrum of substrates as well as disturb diverse types of cells, such as keratinocytes, endothelial cells, leukocytes, red cells and platelets.5,26,27 Importantly, we show herein that L. gaucho venom and its sphingomyelinase fraction promote activation, adhesion and aggregation of both rabbit and human platelets even in the absence of other cells.
Although the intensity of the aggregation was more evident in human PRP exposed to the whole venom or its fraction, since rabbit platelets required a two-fold higher concentration of venom to reach a similar extent of aggregation, both human and rabbit platelets were susceptible to venom- and sphingomyelinase fraction-induced aggregation. Moreover, LDH levels in the milieu of platelets exposed to the venom were not different from basal levels, ruling out the possibility that the increase in transmittance observed during aggregation assay of human and rabbit PRP was caused by platelet lysis. Furthermore, we confirmed that L. gaucho venom-induced platelet aggregation, in PRP of both species, only occurs in the presence of plasma components, such as observed elsewhere for L. reclusa venom in human PRP. 14
The mechanism of platelet aggregation induced by Loxosceles venoms has been poorly investigated. In the last decade, it was reported that sphingomyelinases cloned from L. intermedia and L. reclusa venoms promote LPC hydrolysis to generate LPA,28,29 which can activate endothelial cells and leukocytes in inflammatory events. 30 This phospholipid can also induce platelet activation and aggregation. 31 Our results suggest that L. gaucho venom-induced platelet aggregation is not dependent on the LPA pathway, which depends on the activation of LPA1 and LPA3 receptors on platelet surface. 20 However, a potential participation of the LPA pathway in Loxosceles venom-induced inflammation remains unknown and requires investigation.
In quiescent human platelets, about 20% of GPIIb-IIIa complexes are on membranes of the α-granules and open canalicular system, but depending on activation intensity, they can be exposed on the surface membrane. 32 Although our results indicate that there were no differences in GPIIb-IIIa expression under any treatment, L. gaucho venom and its fraction induced significant increases in the expression of an important marker of platelet activation - the LIBS1 epitope - on both human and rabbit platelets. Once platelets are activated, the GPIIb-IIIa complex suffers a conformational change and then anti-LIBS1 recognizes the epitope on the glycoprotein complex.23,33 In addition, L. gaucho venom-induced increased expression of P-selectin, a widely used marker of platelet activation, 34 confirms the platelet activation state induced by the venom in both human and rabbit platelets. Thus, under in vivo conditions, P-selectin may play an important role in venom-induced inflammation, especially in interactions between platelets and leukocytes/endothelial cells in the bite site. 35
Loxosceles venoms have been investigated only for platelet aggregation activity; therefore, the present work is the first report about the platelet adhesion induced by a Loxosceles venom. L. gaucho venom induces a response that is independent of its concentration, whereas its sphingomyelinase fraction promotes a dose-dependent increase in platelet adhesion. A plausible explanation for this differential response is the existence of many components in Loxosceles venom - e.g., phosphatases, hyaluronidases, phosphohydrolases, metalloproteinases-,4,36 which may interfere in platelet adhesion to collagen, thus preventing the construction of a dose-response curve with the whole venom. On the other hand, the homogeneity of the adhesion results obtained with the sphingomyelinase fraction may encourage the use of this active fraction as a tool to future investigations about molecular mechanisms of platelet adhesion.
Studies about the biological actions of Loxosceles venoms have some limitations, and one of the most important is the spider's small size, which make possible to obtain only a tiny amount of venom for research purposes. Besides these difficulties, comparative studies about the biological effects of Loxosceles venoms and their toxins are important to understand and validate the pathophysiological phenomena occurring in human beings, since certain kind of cells from different species are affected by Loxosceles venoms in different ways. For instance, Loxosceles venoms promote hemolysis in humans 9 but not in rabbits, neither in vitro 37 nor in vivo. 6 Thus, the rabbit is not a suitable animal model to study hemolytic disturbances, even though Barreto et al. 38 reported that L. gaucho venom affects the resistance to both osmotic and shear stress of rabbit red cells in vivo. Regarding platelets, although our results evidence some differences between human and rabbit platelet sensitivity, platelets in both species presented a similar general pattern of response when they were exposed to the loxoscelic venom and its sphingomyelinase fraction.
The current results are in line with recent insights about the participation of platelets in inflammation. Platelet activation, adhesion and aggregation play a prominent role in homeostasis as well as in several pathological phenomena. Thus, platelets can link hemostasis and inflammation through the release of vasoactive monoamines (e.g., serotonin and histamine), chemokines (e.g., PF4/CXCL4, RANTES/CCL5 and CXCL7) and cytokines (e.g., interleukin-1β), and also through cell-to-cell interactions elicited by expression of adhesion and immunologically related molecules (e.g., P-selectin and CD40L).39–41 Taking into account this evidence and that Loxosceles venom-induced dermal necrosis involves hemostatic and inflammatory disturbances in its pathogenesis, platelets may play a pivotal role in the dermal necrosis, not only for their participation in local thrombotic disorders, 6 but also for exacerbating the local inflammatory response induced by the Loxosceles venom.
In conclusion, our data demonstrate for the first time that L. gaucho venom and its sphingomyelinase fraction trigger the main platelet functions in both human and rabbit platelets, thus justifying the use of the rabbit as an experimental animal model to investigate the platelet disturbances induced by Loxosceles venoms. The role of platelets in the pathogenesis of loxoscelism should be addressed in further investigations, which may help to develop more efficacious therapeutic interventions following the envenomation.
Footnotes
Acknowledgment
We thank Marcela S Lira for her assistance with L. gaucho venom extraction.
This work was supported by FAPESP (process 03/00841-0), and it was part of the PhD thesis of F.L.T, accomplished at the Department of Pathology, Faculty of Veterinary Medicine and Zootechny, University of São Paulo, Brazil.