
Abstract
Palaeoecological records from the Maya lowlands indicate that there is an emerging pattern of complex vegetation change across the Terminal Classic Period, a time that was characterised by frequent droughts and collapse of major Maya centres. To fully understand the extent of anthropogenic influences on Yucatan forest composition in space and time, more palaeoecological records are needed, as well as a better understanding of the relationship between forest cover and modern pollen deposition in lakes. Here, we investigate pollen-landscape relationships from lakes in the northern central Yucatan (19°50′N, 88°44′W) through comparison of forest cover data with arboreal pollen in recently deposited sediments. Twelve pollen assemblages from across two lakes, Lagunas Esmeralda (ca. 0.6 × 0.1 km) and Chichancanab (ca. 14.5 × 0.7 km), indicate that the lakes capture local and regional pollen signals, respectively. This spatial understanding was applied to the interpretation of a new 386 cm palaeoecological record dating to 4200 BCE from Esmeralda and an existing record from Chichancanab dating from ca. 200 CE until present. The data show that fire use and maize cultivation occurred at Esmeralda as early as 3500 BCE, but impact on forest cover and composition was minimal throughout. The Chichancanab record shows maize cultivation through the Classic period and relatively stable forest cover. In contrast, high variability of forest pollen values from Laguna Esmeralda during the Late and Terminal Classic periods shows localised changes in forest composition, likely driven by human activity in response to cultural change and climate stress.
Keywords
Introduction
The Ancient Maya civilisation has been studied extensively to address questions of socio-environmental interactions under periods of climate stress (Aragón-Moreno et al., 2012; Diemont et al., 2011; Turner and Sabloff, 2012; Whitmore et al., 1996). Palaeoclimate records have identified periods of drought across the Maya Lowlands of the Yucatan Peninsula (Brenner et al., 2018; Carrillo-Bastos et al., 2010; Hodell et al., 2005, 2007). The temporal coincidence of archaeological evidence for periods of political instability (Hodell et al., 2007; Kennett et al., 2012; Webster et al., 2007) and frequent, sometimes high-magnitude drought events, has highlighted the crucial role of climate change as a key driver of the collapse of the Classic Maya civilisation between ca 750 and 1100 CE (Terminal Classic). Despite the co-occurrence of societal collapse and drought, however, there exists a spatio-temporal complexity in both the patterns of collapse and duration and degree of drought, which has not been resolved (Aimers, 2007; Douglas et al., 2015).
The interactions between societal collapse and regional climate are further complicated by debate over the sustainability of land use practices of the ancient Maya inferred from fossil pollen records, with some indicating extensive forest clearance during the Classic Period (Dull, 2007; McNeil, 2012; Rosenmeier et al., 2002). Reduced tree cover may have exacerbated drought in the Classic Period (Cook et al., 2012) and increased soil erosion and run-off. The latter of these impacts is supported by the occurrence of ‘Maya clay’, the deposition of high abundances of clays in lake sediments in the southern lowlands which is linked to accelerated erosion in the catchment (Deevey et al., 1979; Dunning et al., 1998; Rosenmeier et al., 2002). In contrast, pollen evidence of agroforestry practices during the Classic Period (McNeil et al., 2010; Rushton et al., 2013, 2020) and terracing for soil management and conservation (Beach et al., 2002) demonstrate Maya land use strategies that were sustainable and adaptable to climate stress. These palaeoenvironmental reconstructions support a view that the Maya farming strategies could promote soil retention, canopy cover and biodiversity (Ford and Nigh, 2016).
Maya polities in the northern lowlands experienced growth following the Terminal Classic Period, when southern lowland sites were abandoned (Aimers, 2007), showing a spatial complexity to the socio-political change associated with collapse. The associated environmental changes also suggest that land use in the northern Yucatan was spatially and temporally heterogenous. For example, the record from Coba (site 3 in Figure 1) demonstrates a 60% increase in arboreal pollen in the Postclassic Period, whereas San José Chulchaca (site 1 in Figure 1) in the northwest (Leyden, 2002) and Lake Tzib to the south (Carrillo-Bastos et al., 2010; site 4 in Figure 1), do not show evidence of extensive vegetation change across the period. These differences in vegetation changes are, in part, due to the limited numbers of pollen records across the Yucatan peninsula (Aragón-Moreno et al., 2012; Carrillo-Bastos et al., 2010; Leyden, 2002; Torrescano-Valle and Islebe, 2015) and lack of systematic sampling that captures the full range of past land use strategies, ecosystem types and population densities. Furthermore, the selection of sites for reconstruction of vegetation through pollen analyses needs to be appropriate for the study aim (Jacobson and Bradshaw, 1981) in that (i) the size of the pollen catchment determines the spatial scale represented by the pollen record and (ii) the signal of anthropogenically-driven vegetation change in pollen records will degrade with distance from the site of past human impact (e.g. sites of crop production or habitation). Both of these factors need to considered in the selection of sites to reconstruct past land use in the Yucatan.
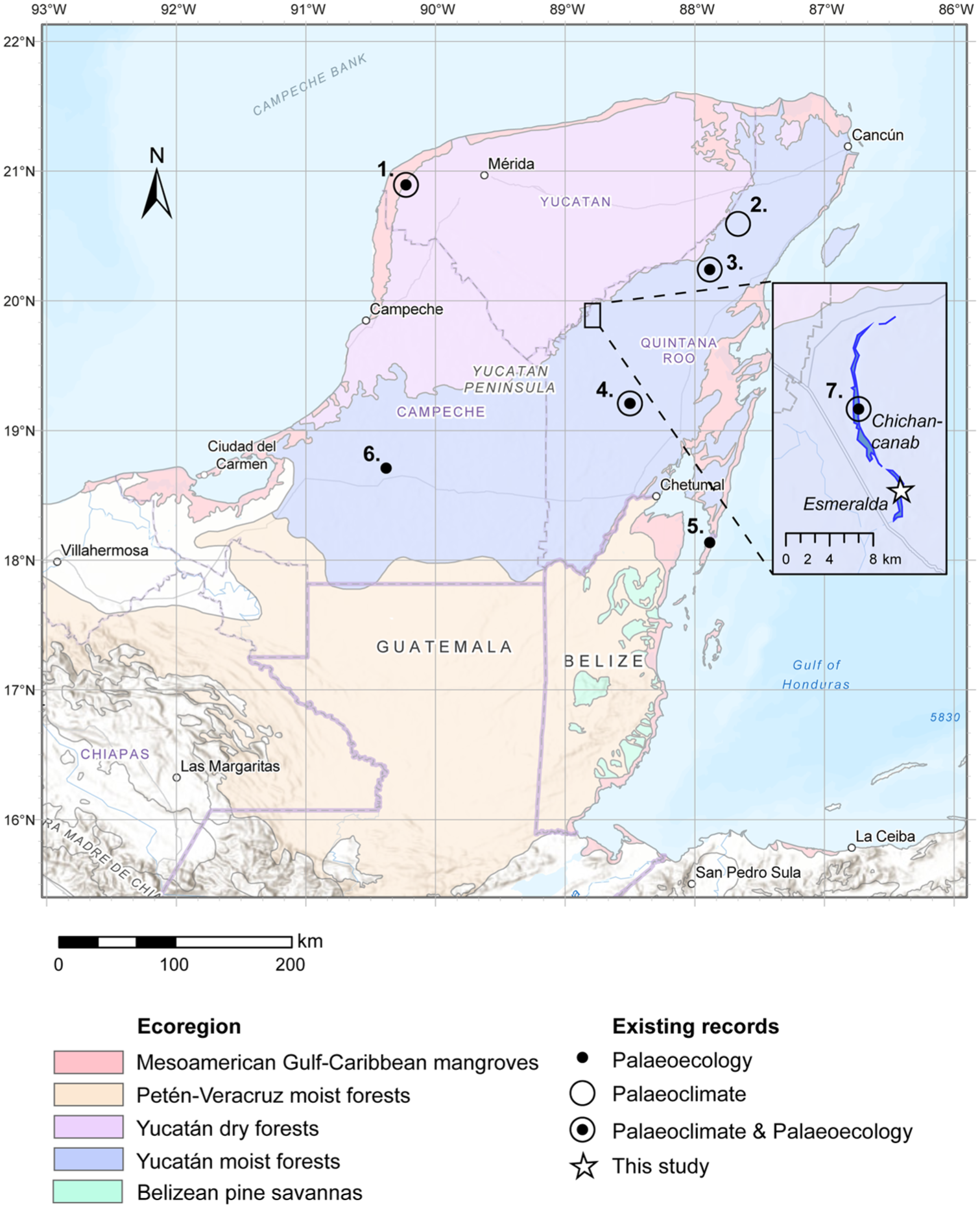
Map of the study area with the key ecoregions of the Maya lowlands (Olson et al., 2001). Sites of the northern lowlands discussed in text are shown: 1. San José Chulchaca; 2. Punta Laguna; 3. Coba; 4. Lake Tzib; 5. Basil Jones; 6. Laguna Silvituc. Inset: 7. Lagunas Chichancanab and Esmeralda (this study).
Although studies linking pollen and vegetation community assemblages have been conducted in the region (Bhattacharya et al., 2011; Correa-Metrio et al., 2012; Villalpando-Aguilar et al., 2022), the relationships between modern pollen deposition, pollen catchment and landscape properties, which are required to fully understand the range and extent of past Maya land use, remains unexplored. The analysis of the most recent sediments (or ‘surface samples’) in depositional environments provides understanding of how the landscape and vegetation structure are represented in the modern pollen signal. This understanding of the spatial representation and uncertainties of pollen records is needed for informed reconstruction of past vegetation. Spatial representation of pollen records is a key consideration in creating robust palaeoecological interpretations (Jacobson and Bradshaw, 1981) which is particularly important for ancient Maya research because evidence of forest clearances derived from pollen-based studies have shaped a narrative of significant environmental degradation in the Classic Period (Cook et al., 2012; Leyden, 2002). Equally, doubt has been raised regarding the representativeness, and therefore interpretation, of the pollen data in reconstructing past land use of the Ancient Maya (Ford, 2022).
The aims of this study are: (i) to investigate the relationship between the pollen deposited in a lake and forest cover in two adjacent lake systems of varying size in the northern Yucatan, and (ii) to apply this understanding to the interpretation of land cover change across key cultural transitions among the ancient Maya through the (re)examination of new (Esmeralda) and existing (Chichancanab) fossil pollen records from the same lakes. Furthermore, the selected lake sites represent different spatial scales of vegetation cover based on pollen source theory, allowing for local land use practices to be distinguishing from wider (regional) climate- and human-driven vegetation change.
Materials and methods
Site description
This study focuses primarily on paired lake systems, Esmeralda and Chichancanab, located in the northern Yucatan Peninsula (19°48′50.4″N, 88°43′40.8″W; Figure 1). The Yucatan Peninsula is mainly a platform made up of carbonates, dolomites and evaporites. Much of the northern part of the peninsula is at less than 40 m a.s.l. The peninsula’s geology means that there is little surface water, with no permanent streams in the north (Perry et al., 2003). There are, however, large numbers of solution lakes and structurally controlled lakes, adjacent to fault lines, including Chichancanab and Esmeralda which are related to the Tikul fault, which forms a 20 m ridge on the eastern shore. There are areas of evaporites to the east and south of the lakes, although they are geochemically different, possibly due to their exact locations within this evaporite region (Perry et al., 2009). The lakes appear to connect during periods of high water level (Hodell et al., 2005). Soil cover across the region is generally thin (Perry et al., 2009).
The climate of the area is typical of the lowland northern hemisphere Neotropics, with a summer rainfall dominated regime driven by the NE Trade Winds and associated easterly waves, convective storms and tropical cyclones (including hurricanes) originating over the tropical Atlantic/Caribbean. There is a general gradient of increasing precipitation across the Peninsula from the NW to SE (De la Barreda et al., 2020). Mean annual precipitation at the meteorological stations closest to the two lakes (Dzuiche for Chichancanab and La Presumida for Esmeralda) is 1191 and 1339 mm respectively. Most rain falls between June and October, with September being the wettest month at both stations. There is limited evidence of the canícula (period of reduced rainfall within the wet season) at these stations. Mean annual temperature at Dzuiche is 26°C, with temperature varying more diurnally than annually. Precipitation is, therefore, the main driver of interannual climate variability.
Seasonally-dry tropical forests (SDTF) are the predominant vegetation formation of the Yucatan peninsula. Forests are deciduous in the northwest where climate is driest and transition to moist seasonal forests in the southeast (Leyden, 2002) across the steep precipitation gradient of the region. Characteristic of SDTF, Yucatan forests contain abundant species of the Fabaceae (Gentry, 1995; Leyden, 2002) and with widespread occurrence of Bursera simaruba, Caesalpinia gaumeri, Gymnopodium floribundum, Piscidia piscipula, and Thouinia paucidentata. Brosimum alicastrum and Croton lundellii are commonly associated with forest growing around Maya ruins (White and Hood, 2004). The pollen spectra of the region are dominated by Brosimum, but many common forest taxa are represented in fossil pollen records, including Acalypha, Bursera, Celtis, Coccoloba, Gymnopodium, Piscidia, Brosimum, Maclura (Chlorophora) and Trema (Leyden, 2002). Fires in Yucatan SDTF are associated with anthropogenic activities and hurricane damage (Vargas et al., 2008). Laguna Chichancanab (Figure 1) has extensive palaeoclimatic datasets that show evidence of droughts during the Terminal Classic Period (Brenner et al., 2018; Douglas et al., 2015; Hodell et al., 1995, 2005), and a pollen record focussed on vegetation change across the late Preclassic to Postclassic Periods (Leyden, 2002). In contrast, no data have yet been published from Laguna Esmeralda. Laguna Chichancanab has a larger surface area (ca. 14.5 × 0.7 km) than Esmeralda (ca. 0.6 × 0.1 km). Both lakes are long and narrow but the width of Chichancanab is sevenfold higher than Esmeralda and therefore is likely to capture a broader (regional or extra-local) scale signal of forest cover and change compared with the pollen signal in Esmeralda.
The lakes are situated in the Cochuah region of the northern Yucatan which contains the ancient Maya centres of Yo’okop and Ichmul, located approximately 20 km away (Shaw, 2015a). Ceramic evidence of human occupation at Yo’okop dates from the Preclassic Period (ca 600–300 BCE) which was followed by increasing size and number of sites through the region in the Late Preclassic Period (Late Formative Period) dating from ca. 300 BCE–CE 250. The transition to the Early Classic Period was characterised by a hiatus in construction across much of the Cochuah region with little to no population in many areas (Shaw, 2015a). In line with the florescence in the Puuc region, populations expanded at Yo’okop during the Terminal Classic Period after several 100 years of abandonment (Shaw, 2015b) and archaeological surveys show significant settlement and economic activities in the Classic and into Postclassic Periods of the Cochuah region.
Field methods
Laguna Esmeralda was cored in 2014 and 2016 using a Livingstone-type corer (Livingstone, 1955; Wright, 1967). There is no visible evidence of human activity/agriculture close to these coring sites, the only ranch reaching to the lake margin was at the extreme southern tip of the lake. Three overlapping cores ES-14 (four drives), ES-16-01 (four drives) and ES-16-02 (four drives) were collected from a depth of 2.63 m (19°46′59.9″N, 88°44′10.7″W). A continuous sequence of 386 cm (Figure 2) was developed using eight individual cores from these sequences based on visual comparison and 14 distinct stratigraphic markers. Carbonate rich sediment dominates the bottom part of the core (below 95 cm, >ca. 2000 cal BP, 50 CE), with bands or fine laminations. Organic content (based on LOI) is generally less than 5%. Above this, organic content (based on loss-on-ignition) increases to 20% or more and banding is less distinct, although there are occasional packets of laminations. In January 2019, surface sediments were collected from lakes Esmeralda and Chichancanab using an Eckman grab from a boat. At both sites, transects were run across the width of the lakes to characterise the differences in pollen signatures from the centres to the lake shores, and in Esmeralda, additional samples were collected at both ends of the lake (Figure 3).
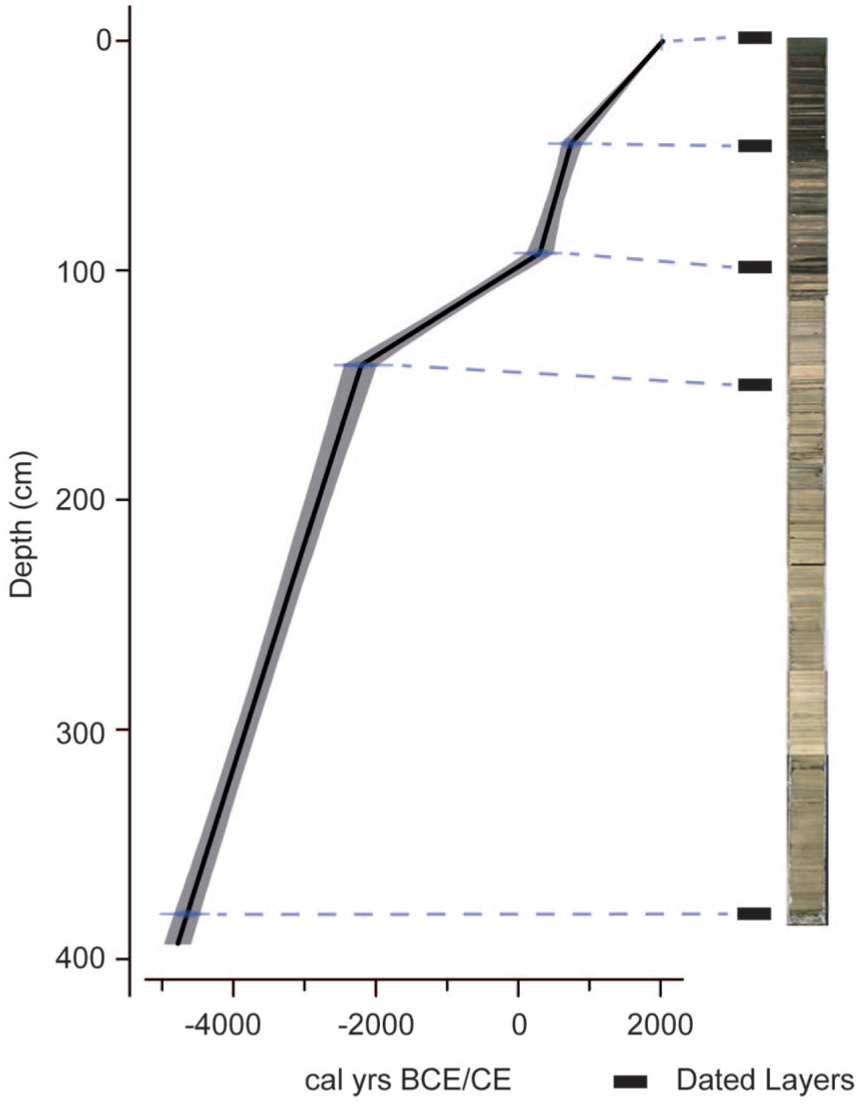
Age-depth model for Laguna Esmeralda. Adjusted (master) depths were used to create the model (Table 1). A composite photograph of the sediment sections (right) marks the locations of the dated horizons. Note that the core images were not adjusted to align to the master sequence. See text for calibration and adjustment of dates for hardwater effect.
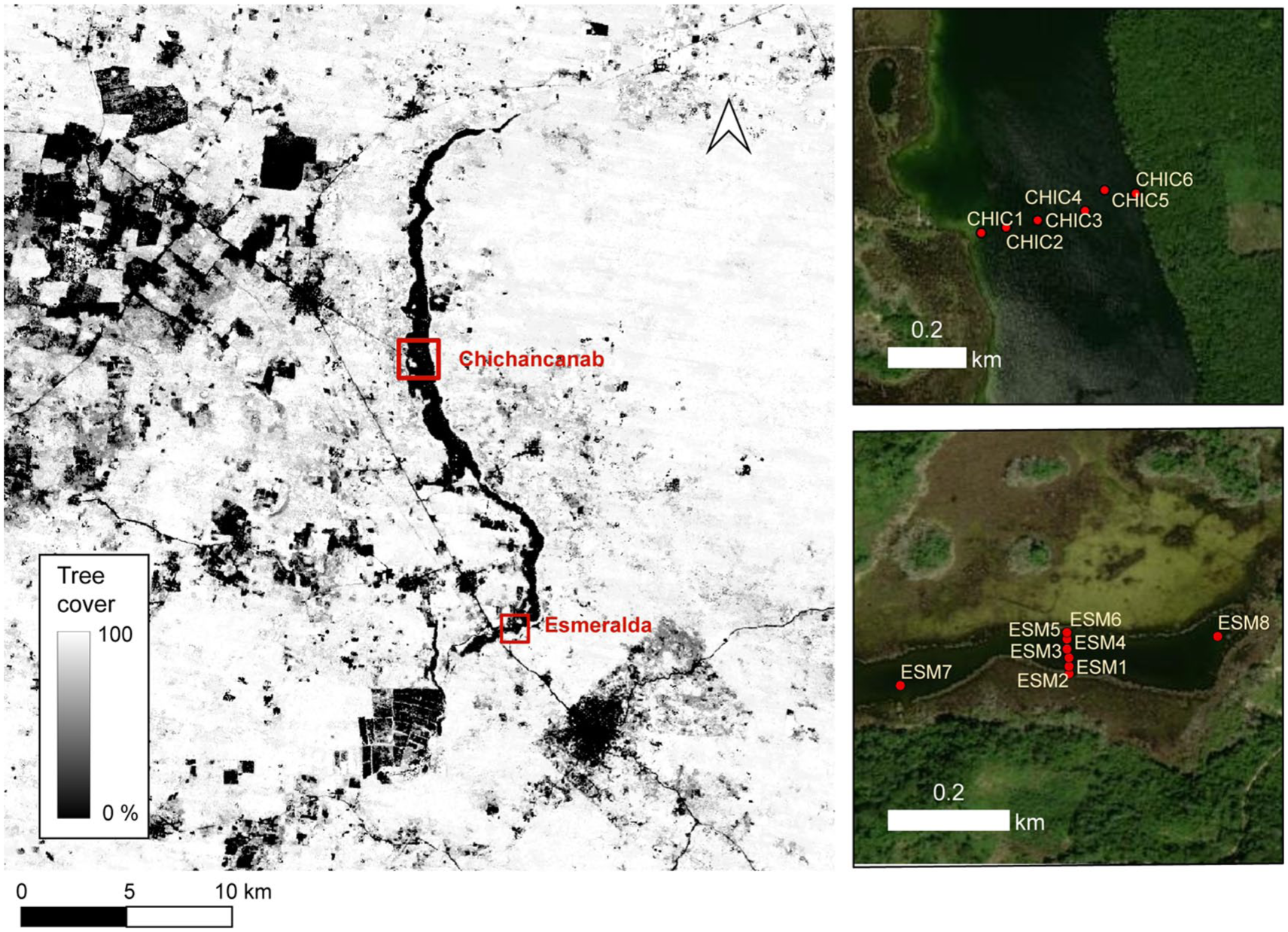
Percent forest cover for the region surrounding Lagunas Chichancanab and Esmeralda from Hansen et al. (2013). Insets show locations of surface samples analysed for modern pollen signals.
Radiocarbon dating and age modelling
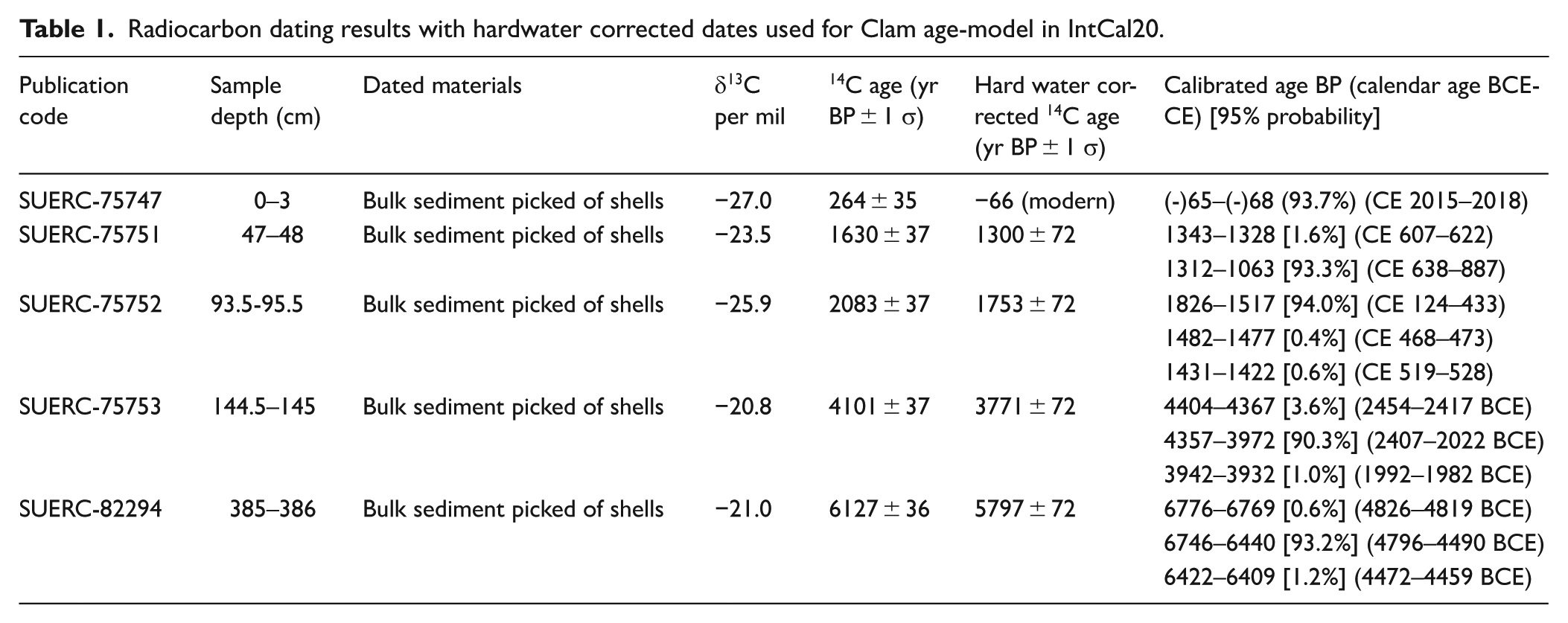
Bulk sediments from the Esmeralda core were dated owing to the lack of available terrestrial macrofossils and five sediment samples, with shells removed, were sent to the NERC radiocarbon facility. The shells were removed as, depending on habitat, they could incorporate a wider range of carbon sources than in open water and there is also the possibility of vital effects. Esmeralda is situated in a karstic region therefore a hardwater offset was calculated and applied to the radiocarbon dating results obtained from sediments. A modern ‘core top’ surface sediment sample (collected using a Glew corer) was dated to calculate the hardwater offset (Table 1), which is the difference between the modern age of the surface sediments (2016 CE) at time of coring and the radiocarbon age of the sediment. This correction factor (330 years) was applied to all uncalibrated radiocarbon dates with a propagated error (±72 years). The corrected dates and errors, and an assumed modern core top, were modelled in R Clam (Blaauw, 2010) with a linear interpolation in R version 4.2.1 using dates adjusted for the hardwater effect (Figure 2). A linear interpolation was chosen because smoothed functions could not adequately incorporate the abrupt changes to sedimentation rates which we have no reason to reject as a true reflection of the age-depth relationship, and the better fitted Bayesian age-models strongly resembled a linear interpolation. The simplest model was therefore chosen. Dates were calibrated using IntCal20 (Reimer et al., 2020). The age model for Chichancanab, following the published dataset (Hodell et al., 1995), was used to constrain dates for the digitised pollen data from Leyden (2002).
Radiocarbon dating results with hardwater corrected dates used for Clam age-model in IntCal20.
Laboratory methods
Pollen and charcoal analyses were performed on 1 cm3 sediment samples from the Esmeralda core, except the lowest 40 cm where pollen was poorly preserved. A 20 cm sampling resolution was adopted for the entire core, with higher 10 cm resolution at key transitions and 2 cm resolution at key Maya cultural periods. Charcoal analysis was performed at every 2 cm. Pollen preparation followed a modified version of the original protocol by Faegri and Iversen (1989), with an additional final sieving stage to allow for the concentration and easier identification of cultigen pollen (Whitney et al., 2012). A standard limit of >200 pollen grains on fossil samples and >150 on surface samples were counted owing to low pollen preservation and concentrations. Ninety five percent confidence intervals were calculated for the surface pollen data using Maher’s lognormal distribution in Psimpoll (Bennett, 2005; Maher, 1972) to show that variation in pollen types to confirm that a 150 count adequately captured variation among the samples (see Supplemental Figs. 1 and 2). Grains were examined at 400× and 1000× magnification using a Leica DM2000 microscope. Pollen identification was achieved with the use of Central American mounted reference materials at the University of Northumbria and a regional reference atlas (Palacios-Chavez et al., 1991). The identification of Zea mays from separate species of the Poaceae family was achieved through the relative size of the grain and the unique exine characteristics (Holst et al., 2007). Published fossil pollen data from Laguna Chichancanab were digitised using the dominant taxa presented in Leyden (2002) and replotted for comparison with Laguna Esmeralda (Figure 6).
Charcoal samples were treated with 10% sodium hexametaphosphate and heated for 4 min (Clark and Patterson, 1997; Whitlock and Larsen, 2001). Samples were then sieved using a modified sieving device at 250 and 125 μm, and counts were amalgamated for CHARanalysis (see below). Charcoal counting was completed using a binocular stereoscopic microscope (Nikon) and a gridded petri dish. Charcoal identification used standard identification techniques in which black, angular and opaque objects were classified as charcoal (Mustaphi and Pisaric, 2014; Whitlock and Larsen, 2001).
Landscape analysis
Estimations of modern forest cover were obtained from the Global Forest Cover dataset from 2000 (Hansen et al., 2013), a Landsat derived dataset of global forest cover resolved to 30 m. Buffers of varying radii 100, 500 and 1000 m were extracted from each surface sample location and the percentages of tree cover were calculated within each delineated area. Water bodies were masked to avoid misidentification of water as ‘non-trees’. Buffers beyond 1000 m yielded similar percentage forest cover values and therefore were not analysed. Additional extracted landscape parameters for comparison with surface pollen assemblage data were ‘distance to nearest forested edge (m)’ and ‘distance to nearest non forested edge (m)’ to characterise the representativeness of forest and non-forested components of the landscape in the pollen signatures of each lake (Table 2).
Landscape parameters derived from forest cover data that were used for pollen-landscape analysis.
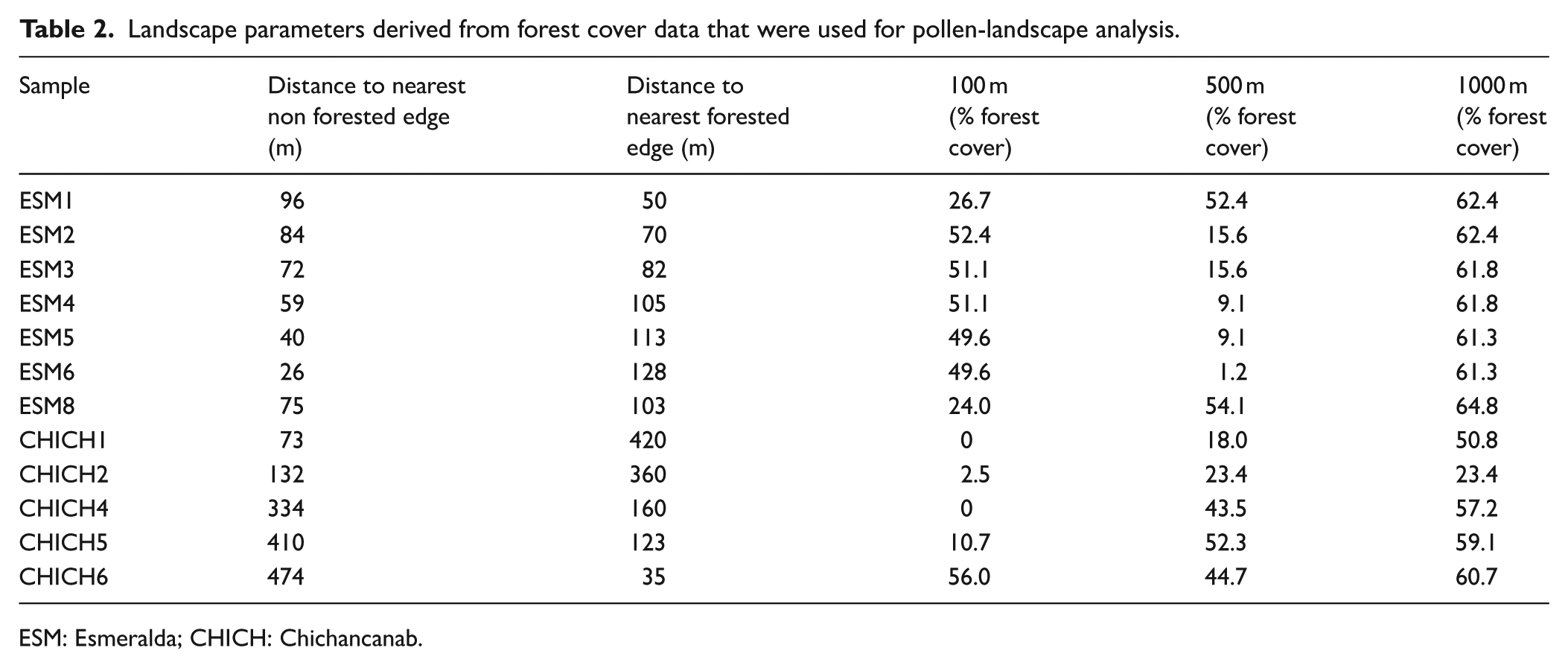
ESM: Esmeralda; CHICH: Chichancanab.
Hansen’s categorisation of trees included in the forest cover data as ‘canopy closure for all vegetation taller than 5 m in height’ (Hansen et al., 2013), therefore pollen types in the surface pollen dataset were classified according to this criterion (‘trees’, ‘non-trees’ and ‘no group’). Where pollen taxonomic resolution is coarse, such as Fabaceae which is a family that includes plants with a wide range of habit (trees, shrubs, herbs), pollen types were grouped as ‘other’. Taxa such as Quercus, Pinus, Rhizophora, Betulaceae and Avicennia were also grouped as ‘other’ because they are not representative of a seasonally dry tropical forest. Cyperaceae were excluded from the non-arboreal sum because of the emergent vegetation fringe within the margins at Esmeralda.
Numerical analysis
Analyses were performed in R version 4.2.1 (R Core Team, 2021). Fossil pollen data were zoned using CONISS in the R package Rioja (Juggins, 2023) and the significance of the zonal boundaries was tested using a broken stick model (Bennett, 1996). For the modern pollen-landscape analysis, the relationship between the surface pollen assemblages and tree cover or shoreline parameters was modelled using beta regression to account for the boundedness and non-normal distribution of proportion data. Analyses were performed using the ‘Betareg’ R package (Cribari-Neto and Zeileis, 2010) and results are presented in Table 3. Analysis of charcoal data to detect peak fire events relative to background levels was conducted using the R package ‘Tapas’ (Finsinger and Bonnici, 2022) based on CHARanalysis originally conceived by Higuera et al. (2009). Charcoal concentration data were resampled at 20-yr intervals (the median sampling interval for the raw data) and these interpolated data were smoothed to model low frequency trends using a lowess function over a 700-yr window. Peak detection was determined using a local threshold provided by a Gaussian mixture model applied through a moving window. The resulting signal-to-noise ratio (SNI) was deemed to be sufficiently high for detecting fire events (Kelly et al., 2011; median = 4.2 ; min, max = 2.2, 11.5; 77% of record with SNI > 3%).
Results of landscape components modelled against summed pollen variables.
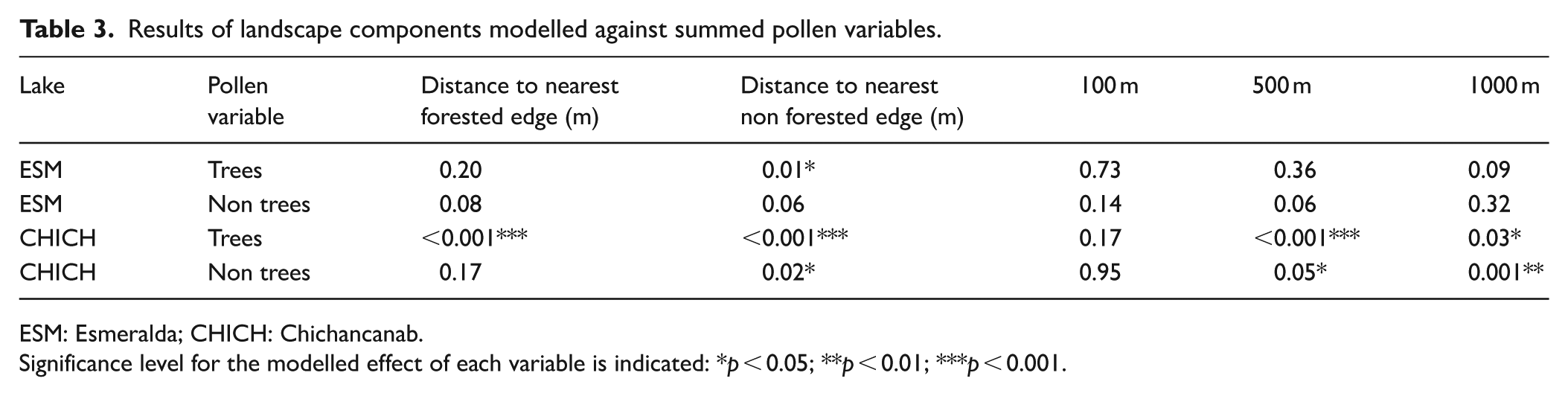
ESM: Esmeralda; CHICH: Chichancanab.
Significance level for the modelled effect of each variable is indicated: *p < 0.05; **p < 0.01; ***p < 0.001.
Results
Landscape-pollen analysis
Summed pollen counts from Laguna Esmeralda and Laguna Chichancanab (Figure 4) show that Laguna Esmeralda (51–76%) has a wider range of arboreal pollen values compared to Chichancanab (65–73%). Moraceae/Urticaceae contribute most to the arboreal pollen assemblages with abundances from the Esmeralda samples ranging from 25% to 61%, compared to the smaller range of Chichancanab (24–39%). Poaceae are the main contributor to the ‘non-tree’ assemblage ranging from 2% to 24% in Esmeralda samples and 2–10% in Chichancanab. Undifferentiated Fabaceae grains were the highest contributors to the ‘other’ assemblage, ranging from 1% to 5% in the Esmeralda samples and 2–9% in the Chichancanab samples (Figure 4). Surface samples ESM-7 and CHICH-3 were not included in the analysis due to low pollen concentrations.

Relative percent abundance of dominant pollen types recovered in surface sediments at Lagunas Chichancanab (CHICH) and Esmeralda (ESM). Panel A. Pollen types classed as Trees; Panel B. Non Trees and No Group; Panel C. Box and whisker plots of total Trees and Non Trees pollen types at each lake.
Although there are limited surface pollen samples for each site, the relationships between pollen and landscape features are distinctive between the lakes (Table 3). Esmeralda pollen values show a significant (positive) relationship only between distance to closest non-forested margin, which relates to highly localised spatial scale. There are no significant relationships in the extra local and regional buffers among the Esmeralda surface pollen samples. Chichancanab demonstrates a significant relationship at all landscape scales (except 100 m) for the forest pollen variable and correlations with non-forest pollen with distance to non-forest edge, 500 and 1000 m buffers.
Esmeralda fossil pollen and charcoal
Fossil pollen and charcoal are presented in Figure 5 with surface sample ESM-4 shown as the core top due to a lack of material for analysis in the core. Pollen taxa were included for plotting if >2% was achieved in one sample and that are not likely to have resulted from long-distance transportation (e.g. Pinus, Quercus, Alnus). Zonation of the pollen record does not produce statistically significant zones, but three zonal boundaries were added for data description. The boundaries were placed in the top three divisions of the dendrogram, expect at the top of the sequence where the divisions would result in a two-sample zone (Figure 5). None of the CONISS clusters align to Maya cultural periods (Figure 5). Moraceae/Urticaceae totals are plotted with separate Brosimum and Maclura (synonym Chlorophora) curves. These genera are morphologically distinctive (Burn and Mayle, 2008) and are common to seasonally dry forests of the Yucatan and abundant in pollen records of the region. Brosimum alicastrum is commonly associated with Maya ruins (White and Hood, 2004). Results of the CHARAnalysis reconstruction which indicate peak events that passed the 95% threshold and fire frequency per 700 years are presented alongside the pollen zones (Z-ESM1, Z-ESM2, Z-ESM3). CHARanalysis resulted in a low number of detectable peaks.
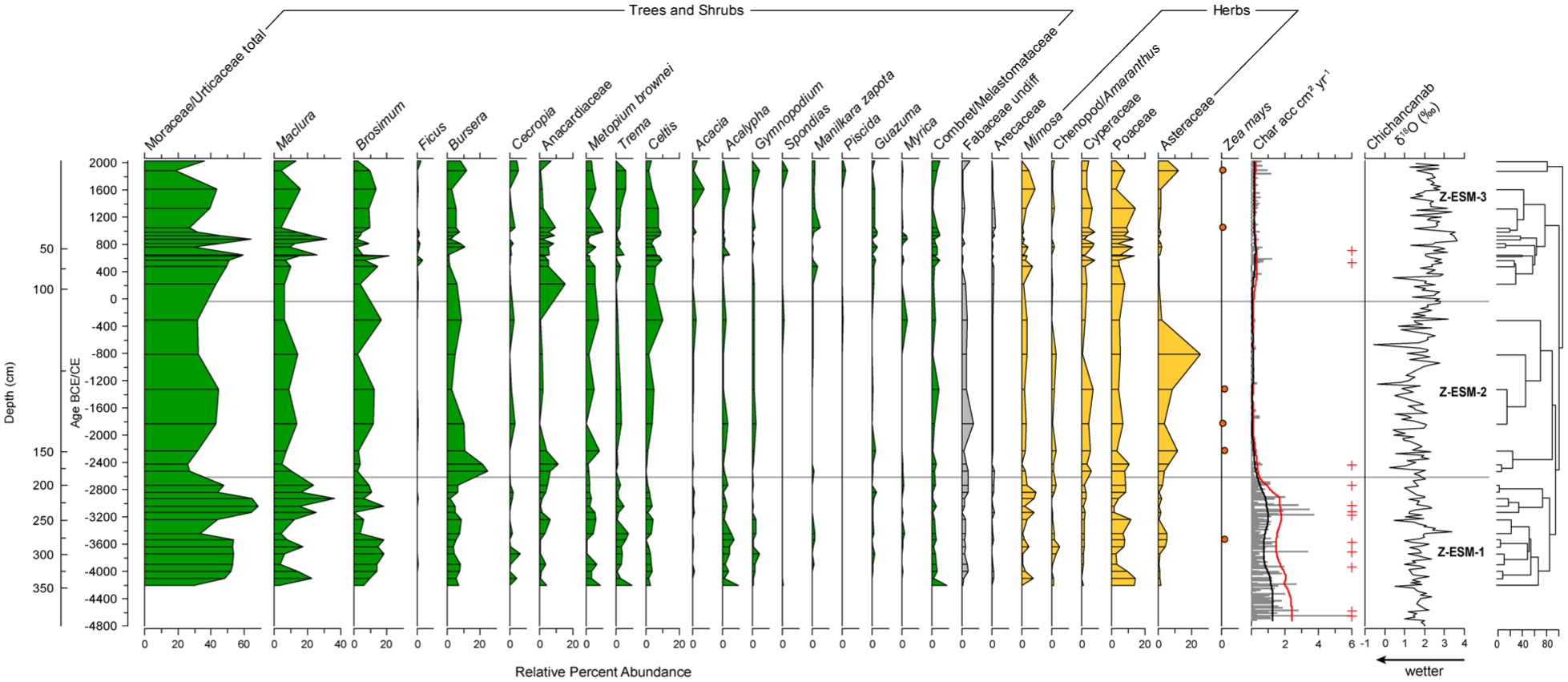
Relative percent abundance of key pollen types recovered from the Esmeralda sediment core. Zea mays is indicted by presence/absence. Charcoal concentrations and resampled accumulation rates are shown alongside the results of the char analysis. Crosses represent fire events and red and black lines indicate 95% threshold value and estimated background charcoal accumulation rates, respectively. Palaeoclimate reconstruction from Chichancanab (Hodell et al., 1995) is shown on the right.
Z-ESM1 344–187 cm: 4200–2630 BCE (14 samples)
This zone captures the first appearance of Zea mays in the record at ca 3530 BCE (3670–3400 BCE, 95% calibrated range). Forest pollen dominates, with Moraceae/Urticaceae (30–68%), Brosimum and Maclura are strongly represented. Disturbance indicators (Poaceae, Asteraceae) and smaller tree abundances (Celtis) are low. Asteraceae show similar low percentages (<3%). Poaceae vary, peaking at the base sample (15%) and then ranging from 1% to 11% throughout. Bursera cf. simaruba (9%) is consistent throughout the zone. Mean assemblage values of other taxa include Trema (3%), Combretaceae/Melastomataceae (2%), Metopium brownei (3%), Fabaceae (2%), Mimosa (3%), and Acalypha (3%). Aquatic taxon Cyperaceae (<3%) appears inconsistently and in low abundances. Nine fire peaks are recorded throughout the zone.
Z-ESM2 187–101 cm: 2630–50 BCE (7 samples)
Forest pollen dominates but there are compositional changes compared with the previous zone. Bursera cf. simaruba, common in secondary forest (Palacios-Chavez et al., 1991) and disturbance indicator Asteraceae reach their highest abundances (25%) in this zone, potentially representing forest degradation. Asteraceae peaks at 820 BCE (25%). Zea mays are recorded between ca 2230–1330 BCE. B. cf. simaruba abundances decline after 2530 BCE through the remainder of the zone, reaching a low of 2% at 1330 BCE. Forest taxa Moraceae/Urticaceae are lower compared to Z-ESM-1 (25–42%). Brosimum is low at the beginning of the zone (ca. 2%) and then increases (2–16%) to reach a peak at 310 BCE. Celtis remains in low abundances until the final sample where it reaches a peak of 10%. Dry forest indicators Anacardiaceae (undiff) peak early in the zone (10% at 2430 BCE) but then decline to <1% and Metopium brownei is consistently low (1–7%). Mimosa (2%), Poaceae (3–10%) and Cyperaceae (ca. 4%) values are consistently present throughout. One fire peak is recorded at 2440 BCE at the beginning of the zone, but fire events are negligible through the rest of the zone.
Z-ESM3 101 cm-surface (50 BCE–modern; 14 samples)
Forest pollen values remained high but variable in Z-ESM3 and show compositional shifts throughout. Moraceae/Urticaceae increase in abundance and spike at 840 CE (64%), followed by a reduction through to the modern surface sample. Higher values of Anacardiaceae are recorded in Z-ESM3, otherwise, the forest community is broadly similar to previous zones but shows high variability of constituent taxa, such as Bursera cf. simaruba (1–12%), Celtis (1–10%), Brosimum (9–22%), Maclura (4–26%), and Anacardiaceae (1–17%). Additional forest taxa represented by the pollen record include Trema (2%), Combretaceae/Melastomataceae (2%) and Metopium brownei (4%). Poaceae values in Z-ESM3 show a muted rise compared to Z-ESM2 and reach >10% abundances after 620 CE. Asteraceae declines to <3% abundances throughout the entire zone, except the final sample where it records 13%. Zea mays pollen is recorded at 1040 and 1870 CE. Cyperaceae ranges from 2% to 8% across the entire zone. Two fire events are recorded at 520 and 700 CE in this zone.
Discussion
The results of the modern pollen-landscape analysis show that pollen spectra of both systems follow expected patterns (Jacobson and Bradshaw, 1981; Sugita, 1994) in that the pollen signal from the smaller Lake Esmeralda is correlated with non-tree pollen in samples closest to the non-forested edge (Table 3). The statistical analysis shows fewer significant correlations between surface pollen at Esmeralda and landscape parameters compared to the analysis of Chichancanab, which suggests that distribution of pollen types is highly heterogeneous and requires more sampling to better capture the spatial relationships at Esmeralda. Nonetheless, the data indicate that localised clearance patches are more strongly represented by this record. By contrast, forest cover and arboreal pollen are correlated across a range of spatial scales at Laguna Chichancanab, showing that the pollen signal reflects a mix of local and regional pollen sources. This study, however, was based on a relatively small sample size and there are additional uncertainties regarding unmeasured environmental variables. In particular, wind direction was not analysed, and the dominant easterly winds may have over-represented the forest cover that dominates the eastern bank of Laguna Chichancanab in the arboreal pollen values of the surface samples. Therefore, more sampling in lakes and across forests of the northern Yucatan, is needed to improve both qualitative and quantitative (i.e. pollen dispersal and deposition modelling) vegetation reconstructions. Nevertheless, this difference in spatial representation of the two lakes allows us to employ a nested sampling strategy (Carson et al., 2014; Urrego et al., 2013) where local and extra-local/regional signals are compared to provide better understanding of the spatial representation of ancient Maya land use across key cultural transitions.
Pollen evidence of maize cultivation occurs early in Esmeralda during the Archaic Period (pre-Maya) at ca. 3530 BCE and 2230 BCE, and the relatively high numbers of fire events probably reflects human presence on the landscape. High arboreal pollen, however, shows that forest dominated the pollen catchment area from ca. 4200 BCE showing that crop cultivation had limited impact on the local forest. Although anthropogenic impact is limited in the Archaic period, rising forest abundances, low charcoal and absence of cultivation indicators between ca. 3530–2230 BCE suggest that early land-use and associated impact was not persistent and allowed for forest canopy cover to remain intact, as also shown in the southern lowlands (Mueller et al., 2010; Wahl et al., 2006, 2007). The landscape was potentially sporadically used to support cultivation during the Archaic, as permanent settlements were not prominent at this time (Lohse, 2010; Rosenswig et al., 2014). High levels of mesic forest elements such as Moraceae are mirrored at Lakes Coba and San Jose Chulchaca, northeast and northwest of Esmeralda, respectively, which indicate relatively moist conditions and minimal anthropogenic disturbance (Leyden, 2002).
A reduction in mesic forest elements and a rise in pollen of Bursera cf. simaruba and Asteraceae at ca. 2630 BCE may be indicative of human-driven forest disturbance, but charcoal analysis shows that burning around Esmeralda declined after Z-ESM1. The isotope-derived palaeoclimate record from Chichancanab indicates that moist conditions generally prevailed until 1050 BCE (3000 years BP; Hodell et al., 1995), therefore changes in the fire regime and vegetation are more likely to have been driven by anthropogenic activities than climate change. Existing palaeoecological records demonstrate the Preclassic is a period of increased ancient Maya influence on the environment, with evidence of deforestation throughout the lowlands (Leyden, 2002; McNeil et al., 2010; Mueller et al., 2010; Rosenmeier et al., 2002; Walsh et al., 2014) and evidence for increasing reliance on maize (Douglas et al., 2015; Leyden, 2002). In contrast, the Laguna Esmeralda record shows some indications of land use, but maintenance of forest cover and few identified fire events.
The pollen record for Chichancanab begins just prior to the Classic period at ca. 200 CE, therefore, land use as represented by these two lakes can be compared across the key Maya cultural transitions from the Classic to Post Classic periods (Figure 6) to understand local forest use (Esmeralda) relative to vegetation changes in the wider region (Chichancanab). At Chichancanab, the record indicates consistently high forest pollen from the late Preclassic Period, although there are no samples dating from the Archaic Period for comparison. Despite these high arboreal pollen values, evidence of maize cultivation is present throughout the Classic period. By contrast, there is high variability of tree pollen at Esmeralda with no recovered maize pollen. Given the modern pollen data show the lake is sensitive to recording local vegetation change, this variability may reflect stochasticity of forest communities near the shoreline. The presence of crops at Chichancanab at this time, however, means that sporadic clearances and shifting cultivation would likely have occurred in the local pollen catchment. It is probable that both people and natural variability are drivers of arboreal pollen changes in Laguna Esmeralda.
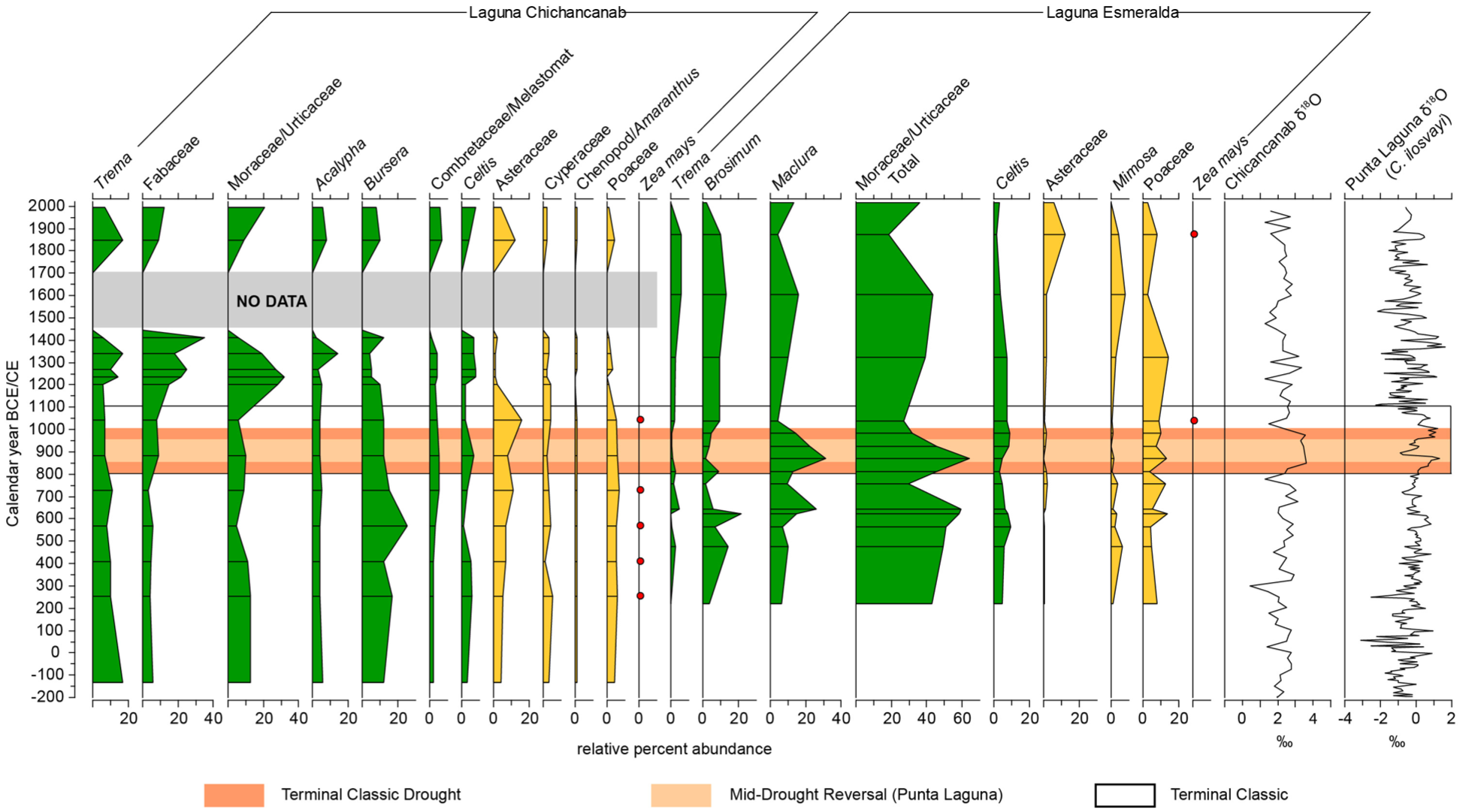
Comparison of pollen from Classic to Terminal Classic periods from Laguna Chichancanab (digitised from Leyden, 2002) and Esmeralda. Palaeohydrological indicators from Chichancanab and Punta Laguna are included for comparison. The Terminal Classic Drought is shown in orange with lighter orange to indicate the reversal identified in the high-resolution Punta Laguna record.
Both Esmeralda and Chichancanab records have relatively high forest cover in contrast with other records from the northern Yucatan, which show pressures on forest resources in the Preclassic to Classic Periods (Aragón-Moreno et al., 2012; Leyden, 2002; Torrescano-Valle and Islebe, 2015). Pressures on forest in this period relate to the development of Classic Maya society and the establishment of major political centres in the northern Maya Lowlands, which is borne out by a decline in arboreal pollen in several records (Leyden, 2002). In the Cochuah region, Yo’okop was one of the few sites occupied in the Early Classic Period whereas many parts of the region had little or no population (Shaw, 2015a). A concentrated population at Yo’okop may explain the relatively low impact on forest cover in our study lake.
The palaeoclimate record at Chichancanab provides clear evidence of droughts around the Terminal Classic, spanning 800–1000 CE (Brenner et al., 2018; Hodell et al., 1995, 2005; Figure 6). The archaeological record indicates complex changes associated with the droughts across the Yucatan. Sites in the south-central Quintana Roo region experienced some form of a decline during the Terminal Classic Period (Villamil and Sherman, 2017), whilst the Cochuah region, including Yo’okop, saw a reversal of an earlier population decline (Shaw, 2015a). In line with these significant cultural transformations and droughts, the pollen record from Laguna Esmeralda shows a complicated pattern of change across the Terminal Classic Period, with highly variable Moraceae (and Brosimum) values. Mesic forest elements initially decline in Esmeralda as the drought event began, but subsequently peak (64%) at ca. 870 CE which broadly aligns with a return to moister conditions that is identified in a later higher resolution reconstruction from Chichancanab (ca. 870–920 CE; Hodell et al., 2005) and a palaeoclimate record at Punta Laguna (ca. 900 CE) located ca. 150 km to the northeast (Figure 1). Although tropical dry forests are tolerant of droughts (Bullock et al., 1995), the timing of the reduction and subsequent rise in Moraceae pollen suggests that the magnitude and cumulative lengths of Terminal Classic droughts was sufficient to impact forest composition. There is only minor variation in pollen abundances of marginal vegetation, such as Cyperaceae and Poaceae, across this period which suggests that varying lake size due to precipitation change (Bush, 2002) was not a key driver of these inferred changes to forest composition. Elsewhere in the region, low forest abundances and high disturbance indicators have been used as evidence of vegetation response to high magnitude drought events at the Terminal Classic Period (Aragón-Moreno et al., 2012; Carrillo-Bastos et al., 2010; Torrescano-Valle and Islebe, 2015). In contrast, Chicanacanab shows a more typical afforestation signal after the Terminal Classic with little indication of drought having impacted the forests, but sampling resolution is low across this period.
In addition to climate pressures, the Terminal Classic period (ca. 750–1100 CE) represents the only established synchronised evidence for cultivation around Laguna Esmeralda and Lake Chichancanab, which suggests that human pressures contributed to the inferred changes in vegetation. The lower forest pollen values at Esmeralda and Chichancanab, alongside presence of maize (1000–1100 CE), suggest there were increased pressures on forest resources at the transition from the Terminal Classic to Postclassic Periods. The Terminal Classic Period was characterised by higher populations at Yo’okop and migration to the region driven, in part, by access to water resources (Shaw, 2015b). It is therefore likely that land use increased around both lakes at this time. Elsewhere in the Yucatan Peninsula, evidence for crop production during the Terminal Classic Period is variable; Lake Silvituc shows no evidence for Zea mays during the Terminal Classic (Torrescano-Valle and Islebe, 2015), while at Lake Tzib, Zea mays and disturbance indicators continue (Carrillo-Bastos et al., 2010; Wahl et al., 2014) and maize cultivation also continued into the Postclassic Period at Coba (Leyden, 2002).
Postclassic reafforestation occurs at Lake Chichancanab from 1100 CE, reaching the highest levels of the record, noting that this record does not include the Preclassic Period. At Laguna Esmeralda, forest recovery occurs between ca. 1000 and 1400 CE but does not reach levels observed in the Archaic Period (Figure 6). The forest composition shifted to include a higher abundance of drought-tolerant taxa (e.g. Anacardiaceae), therefore the lower forest cover may be the result of drier climate compared to the Archaic period. At both lakes, land-use activities during the Postclassic Period show a disappearance of Zea mays along with an increase in arboreal pollen abundances. Although lack of maize pollen does not signal an absence of human presence, the coherence between the Chichancanab and Esmeralda pollen records strongly suggests that people had shifted focus away from the immediate lake area in the Postclassic Period. The trend of reafforestation aligns with pollen records from across the Maya lowlands that show forest recovered over the space of ca. 250 years following the Terminal Classic Period. Both Coba (Leyden, 2002) and Lake Tzib (Carrillo-Bastos et al., 2010) show higher levels of forest cover compared with the Classic Period, even with persistent crop (maize) cultivation. The Postclassic changes observed in our study, however, are muted in comparison and underscore the spatial complexity of anthropogenic impact on forest resources across the peninsula across the Maya cultural periods.
Conclusions
The analysis of modern pollen-landscape relationships demonstrates the need for palaeoecological site selection targeted to specific research questions, especially where there are significant debates on the extent of past environmental degradation and deforestation and heterogeneity of land use in time and space. Here we show that understanding the sensitivity of lakes to detecting changes in the environment improves interpretations of the scale and intensity of past changes in forest cover. The use of paired lakes representing broadly different spatial scales (Esmeralda = local, Chichancanab = mixture of local and regional) has shown increasing pressures in the Terminal Classic that would not have been revealed in an analysis of either lake alone and demonstrates greater land use around these lakes during the Terminal Classic Period. We cannot discount the influence of climate change on forest composition given that tree pollen values coincided with precipitation change during this period, thus human and climatic drivers shaped these forests. High temporal resolution sampling at Esmeralda across the Maya cultural periods shows localised forest clearances but counters prevailing narratives of widespread deforestation. Low charcoal abundances through the Preclassic to Postclassic Periods show that although fire was present and used to manage crops, burning was not extensive in this area. The spatio-temporal complexity of inferred vegetation change throughout the northern lowlands highlights the need for systematic sampling throughout the region to understand the extent of forest cover across a range of past population densities. In particular, palaeoecological records can detect shifting land use in time and space which is vital to uncovering evidence of human migrations during periods of political decentralisation and provide valuable information to address gaps in the archaeological record.
Supplemental Material
sj-docx-1-hol-10.1177_09596836251378027 – Supplemental material for Forest change in the northern Yucatan lowlands (Mexico) during Maya cultural periods: Insights from a paired lake reconstruction
Supplemental material, sj-docx-1-hol-10.1177_09596836251378027 for Forest change in the northern Yucatan lowlands (Mexico) during Maya cultural periods: Insights from a paired lake reconstruction by Adam Bermingham, Bronwen S Whitney, Sarah E Metcalfe, Matthew D Jones, Haydar B Martinez-Dyrzo and Roger Medina-Gonzalez in The Holocene
Footnotes
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by Northumbria University PhD studentship (to AB); Consejo Nacional de Ciencia y Tecnología (CONACYT) studentship 440756 (to HMD); NERC Radiocarbon allocation 2057.0417 (SEM); the University of Nottingham (SEM, MDJ, HMD).
Data availability statement
Data will be made available upon request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.