
Abstract
Reef cores that have been radiometrically dated are increasingly used to examine Holocene reef geomorphology, accretion rates, and community changes. Further, these accretion rates can be compared to sea level change regimes to see if, and how coral reefs have been keeping up with sea level. As Sea level rise is expected to accelerate over the next 100 years, it is important to examine whether these reef complexes are able to “keep up” and avoid “drowning.” Coring has been conducted in many highly studied areas such as the Great Barrier Reef and the Caribbean. However, in the Indo-Pacific region, an area of the highest coral diversity, there have been only a handful of reef cores collected. Here, we present data of 16 cores from two islands of different reef zones from the Spermonde Archipelago, South Sulawesi, Indonesia. Cores were taken from the reef flat and slope, with recovered length varying from 0.41 to 3.53 m, providing a history of each reef after radiocarbon dating. From 7200 to 5500 YBP, sea level rise rapidly increased and these reef complexes accreted at rates able to match this. This is in part driven by a higher occurrence of massive and foliose corals. After this point and toward the present day, sea level declined, causing sub-aerial exposure of these sand cays and the slope now sustaining the growth. This shows that these reef complexes (some of which contain heavily populated islands) are able to keep growing in the face of sea level rise regimes.
Introduction
Tropical coral reefs are unique ecosystems with a high concentration of biodiversity that provide numerous ecological and economic benefits for animals and humans alike (Hoegh-Guldberg et al., 2019). For tropical coastal communities, reefs function as protection, a natural and self-regenerating break from waves and storms (Kench et al., 2022). Further, as reefs accrete and erode, they provide a source of sediment and framework for low lying islands to develop and maintain sub aerial exposure (Kench and Mann, 2017; Montaggioni et al., 2021). Besides the geomorphological processes, reefs also act as a valuable socioeconomic resource for both the fishing and tourist industries in the tropics (Bejarano et al., 2019; Teichberg et al., 2018). The reef functions are maintained, in part, by the complex 3-dimensional structures that reef building corals provide as habitat for other species such as reef fish.
One of the key functions that drives reef growth and resilience, is vertical reef accretion (Gischler and Hudson, 2019; Kench et al., 2022; Morgan et al., 2020), which allows for a sustained buildup of carbonate habitats for countless organisms in the face of sea level rise, bio erosion, and other stressors. This is done by carbonate producers, driven largely by scleractinian corals, but also by foraminifera, mollusks, and calcareous algae (both red and green) (Gischler and Hudson, 2019; Januchowski-Hartley et al., 2017; Ryan et al., 2019). Further, sediment input from terrestrial sources, as well as biogenic silica can add to a reef’s net accumulation (Łukowiak et al., 2018). Accretion rates are impacted by several factors including water depth and therefore light availability and accommodation space (the space available for coral driven sediment accumulation) (Hubbard, 2009; Hubbard et al., 1986). As an increase in water depth increases accommodation space, sea level rise is a major extrinsic factor impacting net vertical accretion (Gischler, 2008; Morgan et al., 2020). Reefs must accrete at rates fast enough to keep water depth at an ideal limit, ensuring that they still receive the optimal amount of light. This leads to a “catch-up” or “keep-up” scenario where reefs are either forced to accrete faster (catch-up) or to maintain their current pace (keep-up) of accretion. If they are unable to do either, then the water depth above becomes too great and they risk becoming a “drowned” reef (Hubbard, 2009). Accretion rates varied across the Holocene, but on average reefs accreted in the 3–10 mm/yr range (Gischler and Hudson, 2019; Hynes et al., 2024; Morgan et al., 2016), which was sufficient to ensure continued accretion in the face of sea level rise. As reefs in the present and into the future are facing unprecedented levels of coral cover loss due to changing climate regimes and direct human impact (through pollution and overfishing), this may result in negative impacts to the broader ecological services that they provide. It is therefore vital to understand how reefs have been able to overcome environmental pressures in the past, and how reef accumulation was able to continue a net positive accretion rate (Morgan et al., 2020; Perry et al., 2015, 2020; Ryan et al., 2019).
Reef growth can be quantified by either measuring the carbonate production or sediment capture side of reef accumulation. Carbonate budgets are a well-calibrated tool used to measure a reef’s net carbonate production at a locality (Januchowski-Hartley et al., 2017; Lange et al., 2020). However, this requires a thorough knowledge of several local factors, such as coral coverage and associated accumulation rates, bioerosion, and specific coral growth rate measurements (Lange et al., 2020; Perry et al., 2013, 2020). Further, these carbonate budgets only measure net production at the locality, and do not account for sediment transport into or out of the system. Alternatively, reef cores, in conjunction with radiometric dating, are a unique and well-studied tool for examining variations in vertical reef accretion (from the capture side) across the Holocene (Cramer et al., 2017; Gischler and Hudson, 2019; Hammerman et al., 2022; Hynes et al., 2024; Morgan et al., 2016; Łukowiak et al., 2018). Such studies build knowledge of past reef accretion at many local, and sometimes even regional, scales. Understanding how a reef has grown in the past under varying climatic scenarios helps us to understand current and future accretionary patterns (Dechnik et al., 2019; Gischler and Hudson, 2019; Morgan et al., 2020; Perry and Smithers, 2011; Toth et al., 2015). The large majority of the cores used in previous studies were taken from well-studied reef systems in the Caribbean and the Australian Great Barrier Reef, lesser so islands in the Pacific and Indian Ocean, and to an even lesser extent, Japanese islands make up most of the cores for the Coastal West Pacific Region (Hynes et al., 2024). Noteworthy is the lack of Holocene reef data from reef cores in the region with the highest coral reef diversity and harboring 30% of the world’s coral reefs, namely the Coral Triangle.
The Coral Triangle is an area of convergence of high biodiversity and speciation, holding over 600 species of scleractinian corals, and is delineated from central and east Indonesia in the West, to Papua New Guinea in the East, and the Philippines in the North (Hoeksema, 2007; Lane and Hoeksema, 2016; Renema et al., 2008). Despite this area long being known as a harbor of tropical diversity, most research only goes back decades and only three Holocene core studies are known from the area, one from Bohol in the Philippines (Grobe et al., 1985) and two more recently from Sulawesi, Indonesia (Kappelmann et al., 2023, 2024). These two other core studies from Spermonde are focused on cores from the islands themselves, and have not investigated the geomorphological history of the reef flat or slope (which make up a large portion of the patch reef complex). Hence, there is little insight into past reef accumulation rates and their relation to environmental conditions in the area, which in turn hinders the ability to predict whether reefs in the Coral Triangle are able to accrete at rates fast enough to keep up with changing environmental conditions, including under future sea level rise scenarios (Morgan et al., 2020). This increase in sea level will especially have an impact on low lying islands, such as those populated throughout the Indo-Pacific regions, which face an increased risk of flooding, or disappearing altogether. Coupled with overfishing and other destructive fishing practices, communities who rely on these reefs could be facing extreme uncertainty for their future (Ballesteros et al., 2018; Bejarano et al., 2019; Teichberg et al., 2018). Furthermore, other more intensively studied regions all have their own varying accretion regimes, and major influencing factors cannot be directly applied from one locality/region to the next. With the knowledge gained from these Holocene reefs, we can apply this to studying the context of current and future reef accumulation within the Coral Triangle.
To better understand how reef complexes (some with sand cays) in the Coral Triangle formed and continued to grow throughout the Holocene, we collected and examined a suite of reef matrix cores from two islands in the Spermonde Archipelago, South Sulawesi, Indonesia. The geomorphological evoultion and accumulation history captured within these cores will allow for an understanding of how these reefs fared under different sea level and climate scenarios during the Holocene. Sedimentary facies, grain size analysis, and other geologic data was collected from these cores. Radiometric dates have been obtained from all cores to determine the timing of these events on the reefs. The Early to Mid-Holocene is a period where significant sea level rise occurred, and during this time low-lying island platforms began to form and accrete. In turn, we can then use this knowledge of reef accretion and formation in the area to better predict how they might respond under present, and potentially even future scenarios. It is especially important and relevant for local populations living in low lying islands and coastal areas to understand how these islands will fare under future sea level rise, as the rate of sea level rise has already doubled in the last 30 years (Walker et al., 2021) and IPCC estimates for the region under an RPC4.5 (moderate emissions scenario) that sea level could be rising by a rate of 5.5 mm/yr over the next 100 years.
Materials and methods
Location and setting
The Spermonde Archipelago, located in Southwest Sulawesi, Indonesia, is a series of some 120 coral islands along a carbonate shelf (Figure 1). Approximately 80 of these islands see continuous sub-aerial exposure (Yusuf et al., 2021). The coral islands are patch reefs that developed on zonated ridges running roughly parallel in a North-South direction, with a patchy barrier reef separating it from the Makassar strait (Janßen et al., 2017; Kench and Mann, 2017). The location of these islands is determined by underlying karst pillars from the Eocene to Miocene Tonas limestone, likely topped by older Pleistocene reefs. The islands tend to follow a general composition pattern - The reef patch complex has a crest (and sometimes an accompanying rampart) at or above sea level on the seaward (Western) rim, which protects a shallow reef flat and accompanying island. The reef flat currently has relatively low coral cover, decreasing in area coverage as you move toward the island itself. On the leeward (Eastern) edge, there is now a less pronounced crest, and the platform slopes off gently to reef base. Outside of the crest on the seaward side is a reef slope where the majority of coral cover and diversity is contained (Girard et al., 2022; Renema and Troelstra, 2001). Here we see approximately 70 genera and over 225 species of coral (Best et al., 1989; Moll, 1983; Veron, 1995, 2000). The most common genera are Acropora, Montipora, Seriatopora, Porites, Pocillopora, as well as the family Fungiidae (Moll, 1983; Moll and Borel-Best, 1984). Between islands, the inter-reef is sandy bottomed, with water depth increasing up to approximately 70 m.
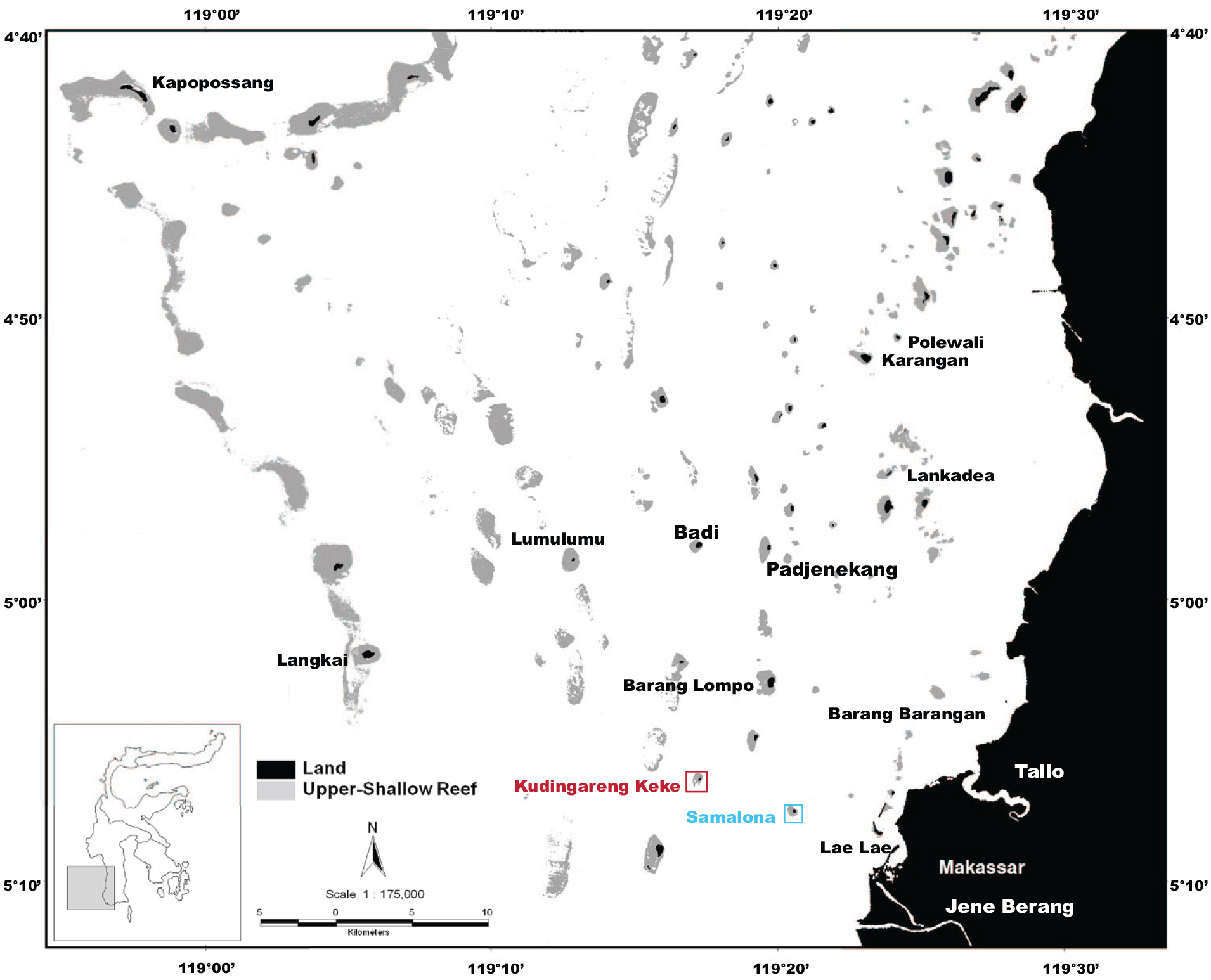
Map of the Spermonde Archipelago adapted from (Plass-Johnson et al., 2016; van der Windt et al., 2020). Blue shows the island Samlaona, red the island Kudingareng Keke.
Adjacent to the archipelago is the large city of Makassar (over 1.5 million people) which sits between three river mouths, the Jene Barang to the south, and the Tallo and Maros rivers North of the city. There are also several smaller tributaries to the north. This influences the entire shelf system via the introduction of nutrients and sediments from the city and rivers onto the reef islands. This influential turbidity regime sees a gradient of high turbidity near the shoreline, with decreasing turbidity moving toward the outer barrier islands. Further, almost half of these islands are inhabited with a dense population covering nearly the entire surface of the island, and anthropogenic structures (such as jetties and breakwaters) being built onto the reef, impacting the ecosystem’s development. Biologically, the Spermonde archipelago is at the center of The Coral Triangle (hereafter referred to CT) and therefore is one of the most diverse reef systems in the world (Hoeksema, 2007; Moll, 1983; Moll and Borel-Best, 1984). However, from in-depth and long-term studies conducted on the area, a decline in coral diversity and coral cover of over 30% has been identified in the region since the 1980s (Best et al., 1989; Moll, 1983; Yasir Haya and Fujii, 2017).
Two of these islands, Samalona and Kudingareng Keke, are close to Makassar in the Mid-Shelf Zone, and the closest part of the Outer Shelf Zone respectively. There is a size difference between these two islands, with Samalona island having an approximate area of 2.2–2.8 ha, whereas Kudingareng Keke is only 1.3–1.6 ha (Kench and Mann, 2017). This difference in size is also accompanied by a difference in shape, with Samalona being more rounded and stable, whilst Kudingareng Keke is a narrow cay. Kudingareng Keke has a deposition bar/beach on the North side of the island that is subject seasonal shifts in beach shape and position due to seasonal changes of wind/wave directions (de Klerk, 1983; Kench and Mann, 2017). Wind, wave, and current energy is directed from the Southeast during the monsoon season which impacts the formation of these islands. However, during the dry season, the dominant energy direction is from the Northwest, but with a much lower magnitude of energy. This same dynamic on the northern shore is also seen on Samalona (de Klerk, 1983), but is less conspicuous due to Samalona’s size, and its high level of vegetation. Interestingly, this vegetation was largely planted by humans in the beginning of the previous century. The total reef area is slightly larger on Kudingareng Keke at 55.8 ha, versus Samalona reef which is only 40.3 ha. Samalona is populated, and was and has been a tourist spot where locals and visitors come to snorkel, dive, and use the island for other forms of activity. Kudingareng Keke has never been permanently inhabited, although a tourist resort was planned to be built there in the 1990s, and it is used as an overnight stay for fishers. In the past decades, a similar style of day tourism as on Samalona has developed. Further, a viewing tower has since been built on the island. Both islands are seeing more frequent use for tourism, and therefore are experiencing an increased impact of anthropogenic activities.
Core collection
We collected 16 Reef Sediment/Framework Cores from two islands (Samalona and Kudingareng Keke, Figures 1 –3) in August 2022 using open barrel push coring techniques (Cramer et al., 2017; Macintyre and Glynn, 1976; Perry and Smithers, 2010; Łukowiak et al., 2018). Six meter-long aluminum tubes with an external diameter of 75 mm and internal diameter of 69 mm were hammered into the reef with a slide hammer. This was done both at the water level on the reef flat, and by aid of SCUBA for the reef slope. Reef slope cores were placed on rubble patches near living coral colonies to avoid harming live coral. Water depth of each core was measured relative to Mean Sea Level (MSL) as calculated previously (Bender et al., 2020; Mann et al., 2016). Cores were taken along an approximate SE-NW transect to follow the island platform shape. From Samalona, five cores were collected on the reef flat (Core Names SLF-[1-5], see Table 1), and three from the reef slope (SLS-[1-3], Table 1), whilst on Kudingareng Keke one core was collected from the island (KKI-1), four from the flat (KKF-[1-4], Table 1) and another three on the reef slope (KKS-[1-3], Table 1). The coring tube was hammered in until it no longer was able to penetrate, usually accompanied by a feeling of hitting hard carbonate.
Metadata for each reef core collected and described within this study.
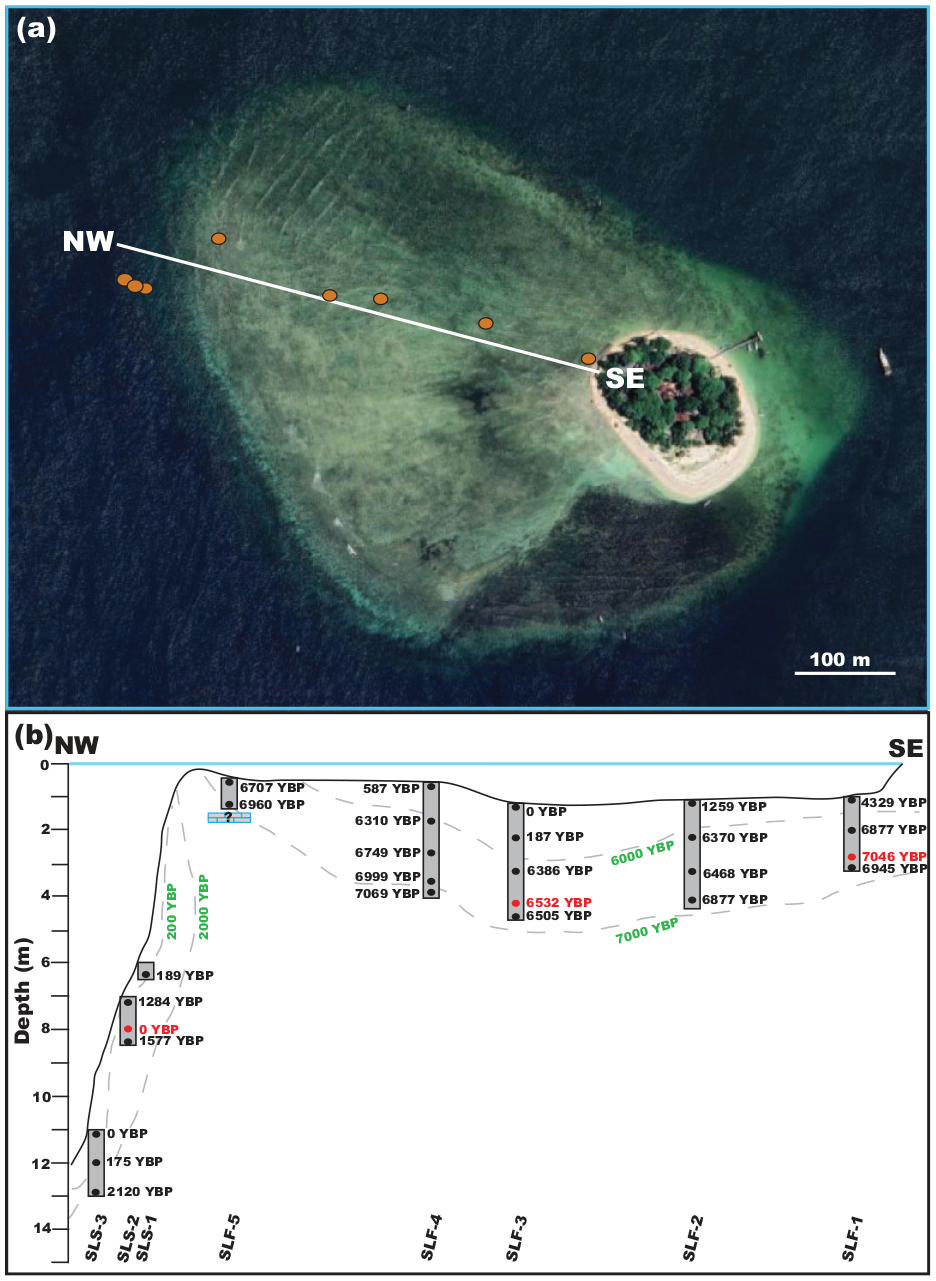
(a) Core transect of Samalona on the island platform (Satellite image Bing 2024). (b) Cross section of coring transect from Samalona. Black dates are calYBP 14C dates. Red dates/* are calYBP 14C dates representing an age reversal. Green dates/dashed lines are geomorphological isochrons. Core names are at the bottom.

(a) Core transect of Kudingareng Keke on the island platform (Satellite image Bing 2024). (b) Cross section of coring transect from Kudingareng Keke. Black dates are calYBP 14C dates. Red dates/* are calYBP 14C dates representing an age reversal. Green dates/dashed lines are geomorphological isochrons. Core names are at the bottom.
Cores were first logged and photographed for sedimentology and stratigraphy following standard sedimentological procedures (Boggs, 2012). They were then broken down into 5 cm subsamples, a common practice for reef cores (Cramer et al., 2017; Palmer et al., 2010; Perry et al., 2011), which helps compensate for compaction, mixing, bioturbation, and time averaging (Kidwell, 1997, 2013). The subsamples were vacuum packed using a food sealer and shipped to Naturalis Biodiversity Center in the Netherlands for further research. The subsamples were sieved into two size fractions (larger and smaller than 2 mm) using standard geological techniques. Sieving was conducted by rinsing with fresh water to remove any salt on the sediments. The >2 mm fraction (pebbles and larger) includes the majority of coral pieces, as well as other carbonate producing organisms (such as mollusks, algae), while the <2 mm fraction (sand and smaller grains) is mostly eroded carbonate matrix material as well as smaller carbonate producers, such as foraminifera, and siliceous sponge spicules. Sieved fractions were then placed in an oven at 55°C for 24–48 h until fully dry.
Radiometric dating
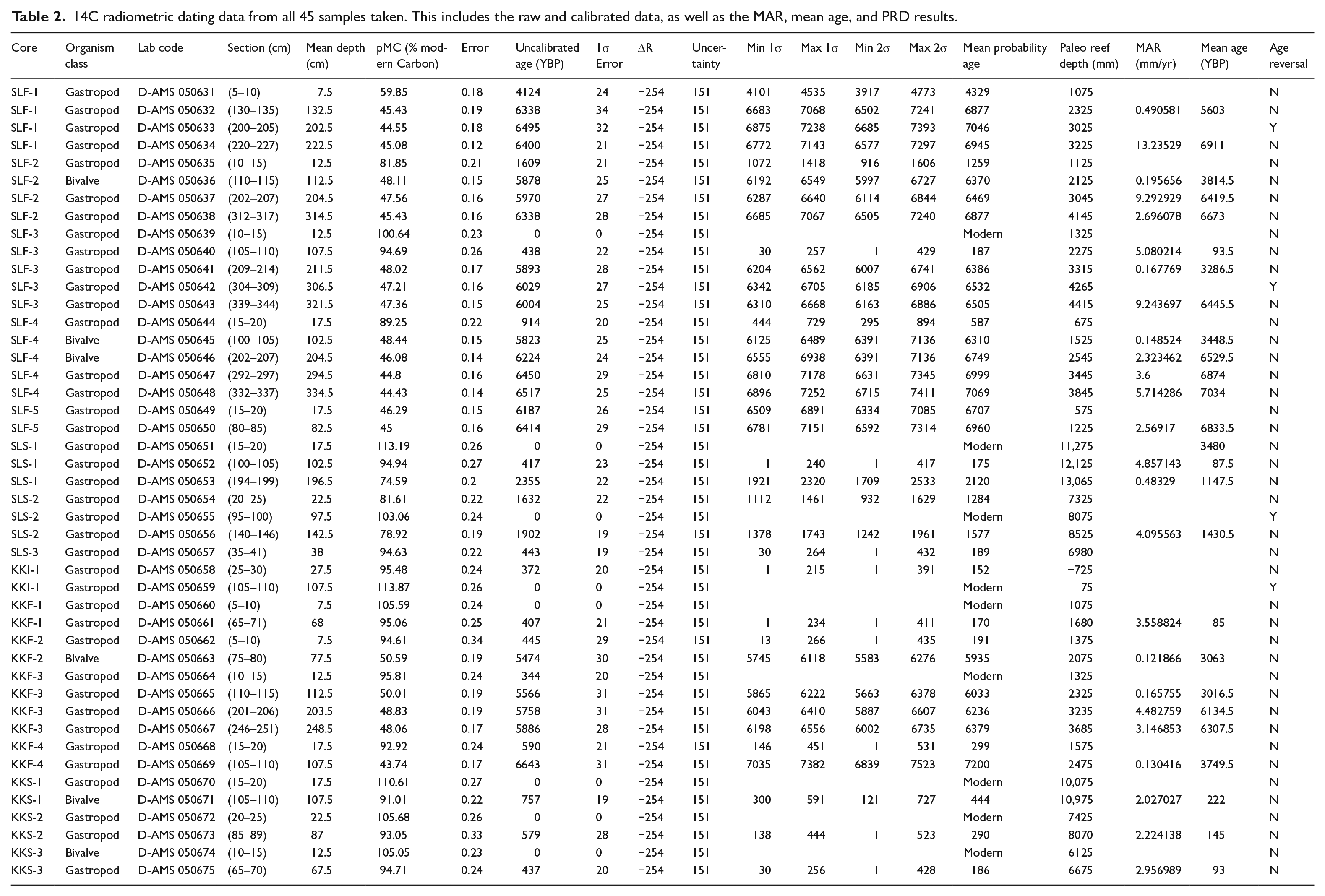
To determine the geochronological framework along the island platform, we conducted radiocarbon (14C) dating on mollusk shells from the cores (Table 2). Carbonate shells were collected from the >2 mm fraction approximately every 1 m downcore. Only shells with enough carbonate material and without visible diagenesis, no external encrusting carbonate organisms, and minimal erosion were used. We chose mollusk shells over in situ corals, as the mollusk shells had much better preservation than the corals, especially for the oldest samples and are easier to determine if there is any allochthonous carbonate (Woodroffe et al., 2007). Shells were cleaned by hand using paintbrushes and dental picks, soaked in Deionized (DI) water, dried at 55°C overnight, then placed into a sonic bath of DI water for 15 min. Samples were then shipped to DirectAMS, an isotope lab in Washington State, where they were acid-etched and placed into an Accelerator Mass Spectrometer (AMS) to obtain conventional radiocarbon dates reported in years before present (YBP). Conventional 14C dates were calibrated using CALIB version 8.2.0 using the Marine20 curve (Heaton et al., 2020; Stuiver et al., 2022; Stuiver and Reimer, 1993) to correct for the marine reservoir effect and then reported in calYBP. As there was no exact local reservoir correction value (ΔR), we averaged the 10 nearest points from the Marine20 Reservoir Database (ΔR = −254 ± 151) and used this to correct the marine reservoir effect.
14C radiometric dating data from all 45 samples taken. This includes the raw and calibrated data, as well as the MAR, mean age, and PRD results.
Calculation of accretion rates
Mean Accretion Rates (hereafter called MAR) were calculated as previously described in the RADReef database (Hynes et al., 2024). This requires a minimum of two dates per core as it determines the difference between depths (D) divided by the difference in ages (t) using the following formula:
Grain size analysis of sediments
Grain size analysis was conducted on the <2 mm fraction to determine if reef sediment infill has changed through the Holocene. A few grams of sediment were collected approximately every 25 cm downcore. These were added to a Beckman Coulter Laser Diffraction Particle Size Analyzer (0.04–2000 µm size range) with an Aqueous Liquid Module (LS 13 320) until obscurity was between 8% and 12% then run multiple times until the grain size curve stabilized. These multiple runs were done to account for clumped sediment as well as air bubbles that may have entered the laser scanner from the Reverse Osmosis water system. The final run of each sample was then used for comparison. This was done to determine the distribution of sand, silt, and clay particles for each subsample.
Sea level curve
To examine the effect of sea level on accretion rates and island geomorphology, we generated a regional Relative Sea Level (RSL) curve for the Coastal West Pacific. These were generated by using RSL and accompanying calibrated 14C dates previously calculated (Lambeck et al., 2014). Dates were already calibrated within the study for uniformity and were therefore not recalibrated here. The Paleo Sea Level Region examined here, Coastal West Pacific, was created using data points previously collected to determine sea level regions (Khan et al., 2019). The RSL curve was generated using R package ggplot2 to create a loess regression with a 95% confidence interval (Wickham, 2016).
Results
Cores
Recovered lengths of the cores varied between 0.41 and 3.53 m (average of 1.61 m, SD 1.03) where post compaction recovery % (recovery/penetration × 100) ranged from 30.6% to 85%, with an average of 58.3% (Figures 2 and 3, Table 1). The low average recovery percentage is likely due to compaction from percussion coring, as well as some incidental material loss from the bottom of the cores during core retrieval. The longest cores were all collected from the reef flat and the average length of recovery was almost 1 m longer than on the slope (2.06 m, SD 1.18 flat, vs 1.13 m, SD 0.59 slope). We collected three reef flat cores with a recovered length of over 3 m from Samalona, and only one from Kudingareng Keke. Furthermore, on Samalona we collected one reef slope core of over 2 m recovered length. Average recovery percentage on Samalona was slightly higher (61.6%) compared to Kudingareng Keke (55.4%), but both are comparable to the total average recovery value. On the flat of Kudingareng Keke, three out of four cores hit an impenetrable carbonate surface (where coring could no longer continue, and the coring pipe had a metallic ring due to hitting solid carbonate) which terminated coring, whereas on Samalona this only occurred once on the flat.
Core lithologies/facies
Facies were determined by combination of the most dominant coral morphology (branching, foliose, or massive), as well as by determining if they were rudstone (the >2 mm fraction is more than 50% of the subsample) or floatstone (the >2 mm fraction is less than 50% of the sub sample). Rudstone was used for the clast supported facies as the clasts were in contact with each other. This was done as we could not definitively say that all of the >2 mm components were in situ, even though they all were produced locally within the same patch reef system. Within all cores collected here, six different facies were identified. Figure 4a represents the branching-dominant rudstone facies, where there are large and numerous pieces of branching framework supporting the infilled matrix. The branching floatstone facies (Figure 4b), again sees branching corals dominant, but with the infilled sediment matrix being the majority of the core. A massive dominant rudstone facies (Figure 4c) has a majority of massive coral pieces, and in many cases the massive coral takes up the majority (if not all) of the coring tube. The massive/branching rudstone facies (Figure 4d) has a mixture of branching and massive corals dominating, often there are pieces of a massive coral within the subsample (most frequently pieces of a massive Galaxea colony). Figure 4e shows the branching/foliose rudstone facies comprised of a majority (but relatively equal amounts) of branching and foliose corals dominating the sub sample. Finally, Figure 4f, is the mixed rudstone facies, a clast supported facies of all three major morphologies mixed together. The majority of cores returned a rudstone, sand matrix-infilled facies (Figures 4 and 5). These facies frameworks were dominantly branching coral (Figures 4 and 5). However, there also were 4 and 3 reef flat cores from Samalona and Kudingareng Keke respectively, with a facies of massive dominantly or massive mixed rudstone facies appearing before and up to 5500 YBP. Reef slope cores tended to have more massive and foliose corals, with Kudingareng Keke slope cores having dominant foliose/massive/branching coral mixes. Samalona slope cores showed more dominantly branching facies, but with a higher occurrence of foliose and massive corals than seen on the Samalona flat cores.

Core and >2 mm size fraction corals for each facies described in this studies cores. (a) Branching dominant rudstone facies. (b) Branching dominant floatstone facies. (c) Massive dominant rudstone facies. (d) Branching/massive rudstone facies. (e) Branching/Foliose rudstone facies. (f) Mixed morphology rudstone facies.
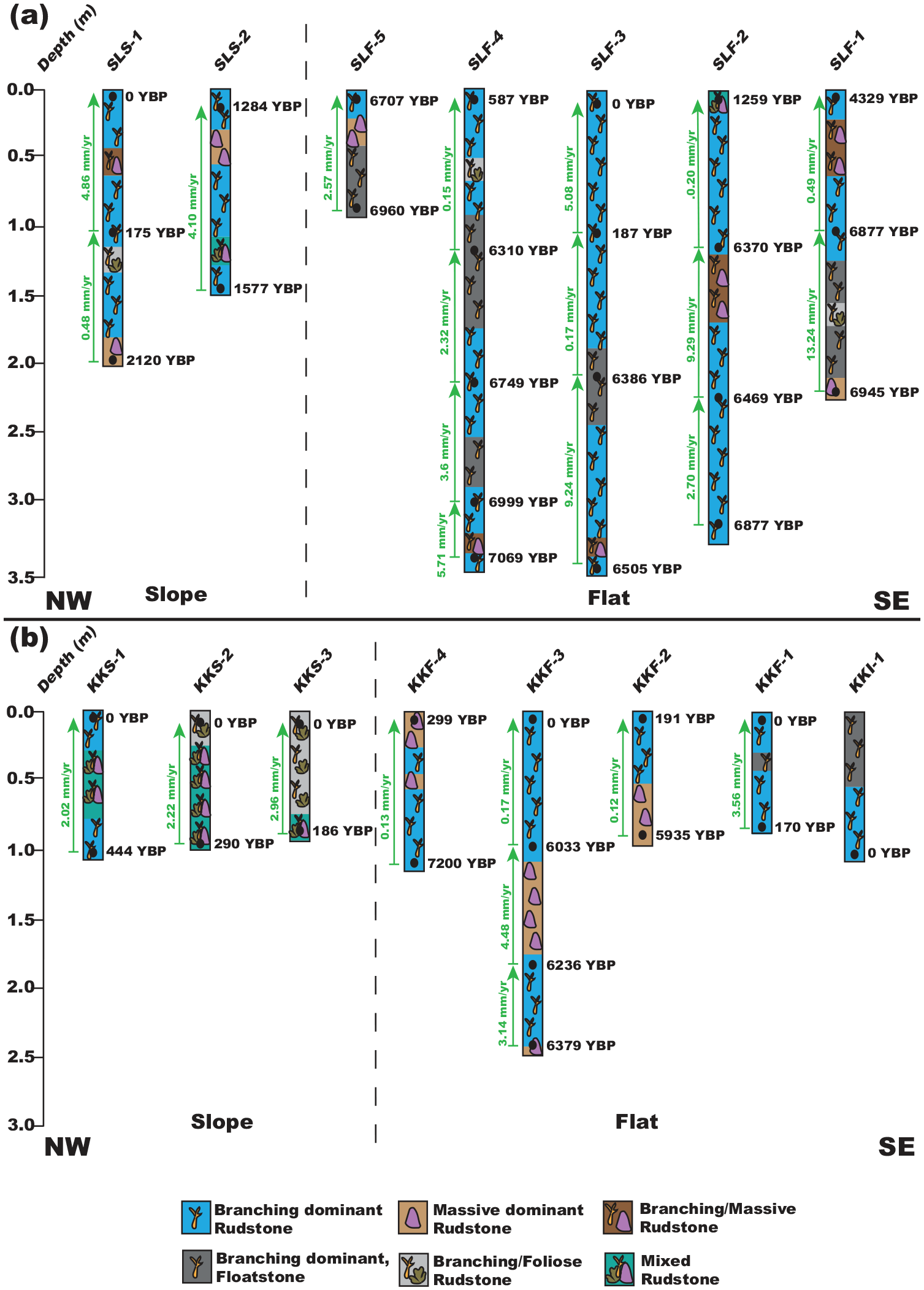
(a) Mean accretion rates (green/arrows), calYBP 14C dates (black), and facies of Samalona cores. Core names are at the top. (b) Mean accretion rates (green/arrows), calYBP 14C dates (black), and facies of Kudingareng Keke cores. Core names are at the bottom.
Almost all reef cores showed a general grain size fining downcore trend (mean grain size). Grain size is very to extremely poorly sorted across all cores (Figure 6, Supplemental Figure 1). Reef flat cores went from coarse sand mean grain size at the top of the core to fine sand at the bottom, with the transition beginning between 0.5 and 1.5 m downcore (Figure 6, Supplemental Figures 2 and 3). As the mean grain size reduces toward medium to fine sand, with finer sand more likely at the bottom of the longer reef flat cores, the remaining grain size is increasingly filled by silt and mud sized particles. This is accompanied by a visual shift in color from a brown coarse sand matrix to an increasingly gray lime mud matrix. Reef slope cores (Supplemental Figures 1 and 4) on Samalona show a very slight fining downward sequence, from coarse to medium sand. On Kudingareng Keke slope, there is a much more pronounced fining downward sequence from very coarse/coarse sand to fine sand. The Kudingareng Keke island core is dominantly sand matrix, with most coral fragments of >2 mm being eroded when present. This creates an almost entirely matrix dominant floatstone facies with minimal reef framework. Coral is minimal in the >2 mm fraction of this core, and the grain size shows poor sorting and does not follow the downward fining sequence we see in other cores from this study.
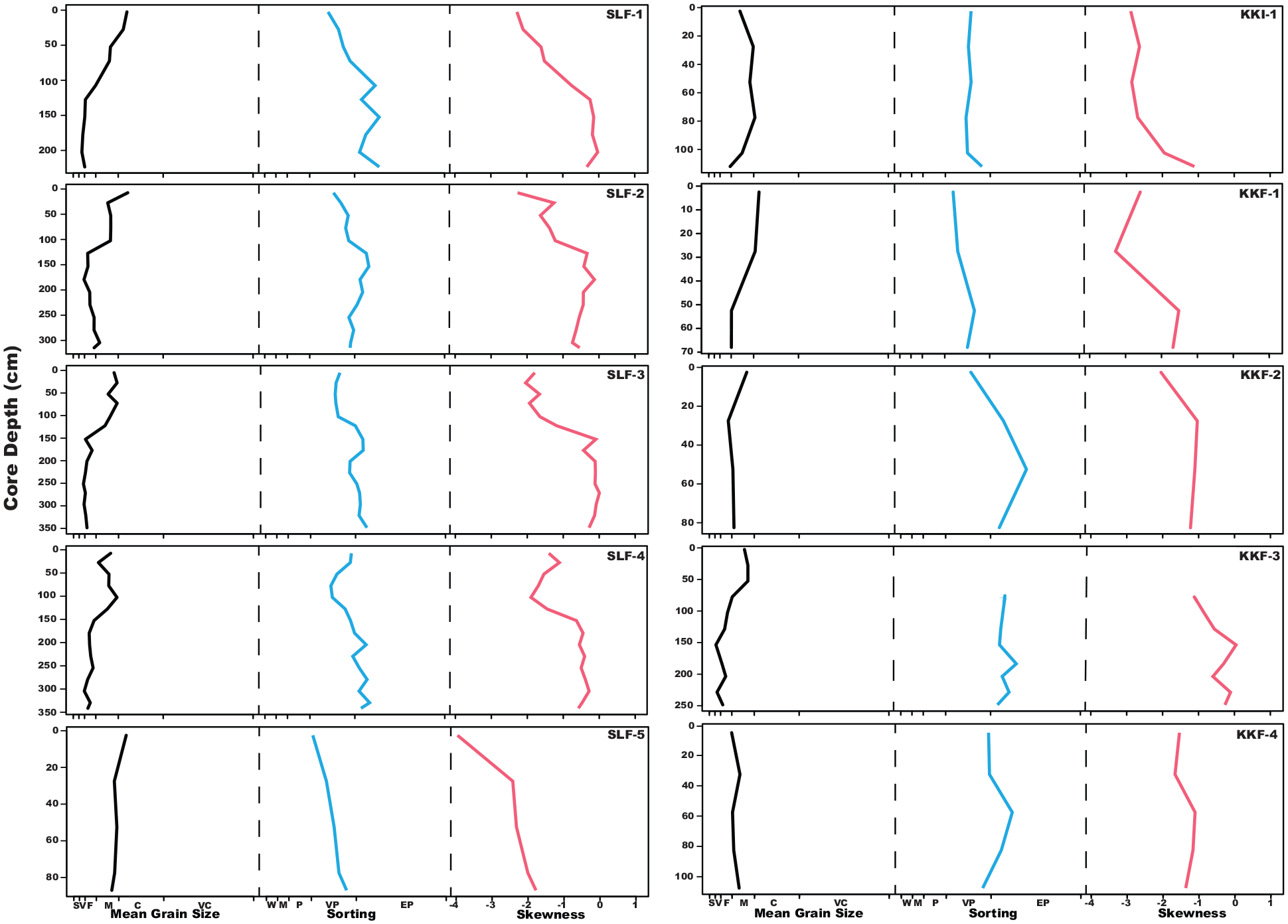
Mean Grain Size, Sorting, and Skewness Curves for Samalona and Kudingareng Keke reef flat cores. Core name is on the top right of each set of curves, Samalona cores on the left, Kuginagreng Keke on the right. . For Mean Grain Size, S is silt/clay, V is very fine sand, F is fine sand, M is medium sand, C is coarse sand, VC is very coarse sand. For Sorting, W is well sorted, M is moderately sorted, P is poorly sorted, VP is very poorly sorted, EP is extremely poorly sorted.
Radiometric dates
We returned 45 radiocarbon dates, of which four represented age reversals, three on Samalona and one from the Kudingareng Keke island core (Figures 2, 3, and 5, Table 2). These reef flat age reversals are less than 100 years and are close to the 2σ range of the 14C dates, and therefore may not be a true age reversal. Still, all age reversals were excluded from calculation of MAR for more accurate comparisons both between islands and to global and regional MAR (Hynes et al., 2024). On both reef flats the majority of cores are Modern – 1300 YBP at the core top. The only exceptions to this are the Samalona cores taken near the island and the reef crest (4329 and 6707 YBP respectively). Moving downcore by 1 m then yields an age between 5900 and 7200 YBP on all flat cores except SLF-3 (mid reef flat), where this transition occurs approximately 2 m downcore. Reef slope cores are much younger, with all but one being modern at the top and reaching up to 2120 YBP at the bottom. Paleo reef depth (PRD) was plotted against the calibrated Ages (YBP) as seen in Figure 7, and compared and contrasted with the other core study from Spermonde (Kappelmann et al., 2023).
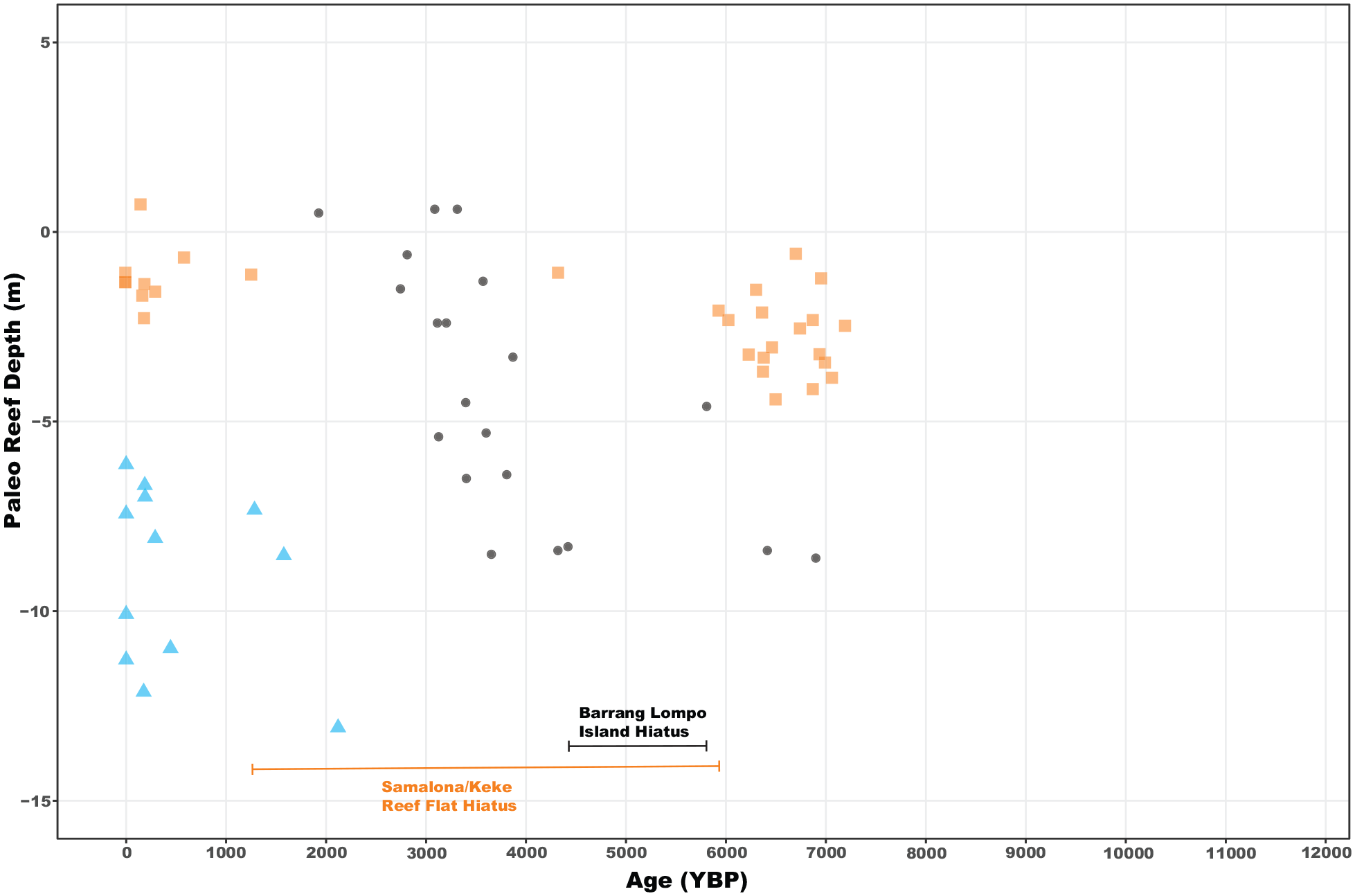
Paleo reef depth (core depth + water depth) versus age (YBP) plot. Orange squares are the reef flat samples and blue triangles are the reef slope samples from this study. Black dots are from Kappelmann et al. (2023) from Barang Lompo an inhabited island in the Spermonde Archipelago.
Accretion rates
Reef flat accretion can be broken up into two distinct periods, Early to Mid-Holocene (approximately 7200–5500 YBP) and Mid to Late Holocene (5500 YBP to Modern). On Samalona in the Early Holocene we found accretion rates ranging from 2.32 to 13.25 mm/yr with a mean of 6.08 mm/yr. After this point accretion slows down drastically to an average of 1.20 mm/yr (0.15–5.08 mm/yr). Similarly, on Kudingareng Keke we found accretion rates ranging from 3.14 to 4.48 mm/yr with a mean of 3.81 mm/yr in the Early Holocene. The Late-Holocene on Kudingareng Keke saw MAR ranging from 0.12 to 3.56 mm/yr, with a mean of 1.00 mm/yr, mirroring the MAR slowdown observed on Samalona. Reef slopes only cover the last 2100 years, and therefore can only tell us about MAR from this period. Here we observed a MAR ranging from 0.48 to 4.86 mm/yr with a mean of 2.77 mm/yr. These rates, while lower than Early Holocene reef flat accretion, are still higher than modern accretion on the flat.
Sea level and paleo reef depth
Our reconstructed PRD are plotted relative to the Coastal West Pacific (Lambeck et al., 2014) in Supplemental Figure 5. We distinguish two phases: (1) a rapid increase in sea level from the Last Glacial Maximum (LGM) into the Mid-Holocene (Supplemental Figure 5). In this phase all reefs were rapidly accreting to follow sea level rise up to approximately 5500 YBP, a sea level highstand in the Middle Holocene, followed by a maximum 2 m sea level drop. Accretion rates dropped on the reef flat, but remained higher on the reef slope. It is only in the last few decades that sea level has started to rise again (Walker et al., 2021), but this is not visible in Supplemental Figure 5 due to 14C dating limitations.
Discussion
To examine reef platform formation and island growth in the Spermonde Archipelago (SW Sulawesi, Indonesia), reef cores were examined for geologic and geomorphologic factors. We built a robust geochronologic framework by integrating 45 radiocarbon dates from 16 cores, and we managed to reconstruct the past 7200 years of reef development. We calculated the mean accretion rates during this time interval for two islands, Samalona positioned in the Mid-Shelf, and Kudingareng Keke from the Outer Shelf. These are roughly similarly sized and shaped islands that are subject to different environmental conditions and have different coral cover patterns in the present day, but seemingly formed under similar circumstances in the Early to Mid-Holocene. It was found that both islands have similar age to depth ranges (Figures 2, 3, and 5) and accretion rates from both reef flat and reef slope cores. In addition, they have similar sedimentary facies, dominantly branching rudstone, on the flats, with massive and foliose dominant facies more commonly seen prior to 5500 YBP. The grain size regimes also show similar patterns, and therefore geomorphological processes for both islands are subject to the same controlling factors, albeit with minor local differences. Accretion rates are faster approaching the Mid-Holocene (over 6 mm/yr) but slow significantly to just over 1 mm/yr toward the present day reflecting a lack of accommodation space due to declining sea level. Based on these observations we discuss the geomorphological development of these islands in relation to environmental conditions. We do this by discussing two phases in reef development, the first during the Early to Mid-Holocene sea-level rise, the second during the Mid- to Late-Holocene sea-level drop. Finally, we integrate these data into a model of reef development of sand cay reefs.
Early to Mid-Holocene reef flat development (7200–5500 YBP)
While the exact point of reef initiation could not be determined due to the cores not recovering Pleistocene basement, it is likely that the oldest ages are close to point of reef imitation as the shallow depths of the mid to inner shelf of Spermonde (Kudingareng Keke and Samalona respectively) were flooded by 9000 YBP (Mann et al., 2016, 2019). Core data collected here from both islands demonstrate that the early Holocene started with reefs growing in deeper water conditions. In two reef flat cores from each island we observe a transition from a massive/foliose coral dominated rudstone facies to a branching coral dominated facies, which we interpret here as a shallowing trend. However, it cannot be ruled out that part of this trend is the result of lateral variation in facies (Webster and Davies, 2003). This period coincides with the final stages of sea level rise as the result of deglaciation following the Last Glacial Maximum (LGM) (Bender et al., 2020; Kappelmann et al., 2023; Mann et al., 2016, 2019). Both reef complexes are primarily accreting to fill the accommodation space being created by sea level rise (Dechnik et al., 2017; Hubbard et al., 1986; Twiggs and Collins, 2010). Rates of accretion have a mean of 3.81 and 6.08 mm/yr on Kudingareng Keke and Samalona respectively, which fall well within the range of the global average of accretion rates during this period (Hynes et al., 2024). As a way to keep-up with increased accommodation space, the reef shifts to a dominant branching coral rudstone facies. (Hubbard, 2009). This facies becomes the norm toward the Mid-Holocene on both reef flats.
The reef crest likely reached maximum accommodation space (1–2 m maximum water depth) early within this period, as seen by core SLF-5 where the top and bottom are between 7000 and 6000 YBP. As the reef likely become more rugose and complex, the sediment influx by both erosion of corals/rubble and input from extrinsic sources built up from the Western margin (the crest) toward the leeward side, especially during the wet season (Janßen et al., 2017). A true crest never developed on the leeward side, and to this day exists more as a gentle slope on both islands. By the end of this period, the island itself is already forming on the eastern half of the platform, toward the leeward margin. The reef flat approached sea level by 5500 YBP (sea level highstand), and is formed mostly of carbonate sediments derived from local carbonate production, subsequent reef bioerosion, and transport of sediments across the reef flat.
Mid holocene hiatus and Late-Holocene reef flat development (5500 YBP–present)
From the Mid-Holocene, where regional sea level rise not only slows, but actually begins to decline by up to 1 m (Mann et al., 2016, 2019), reefs here shifted from an accretion phase to a windward progradation/leeward accumulation (Dechnik et al., 2015, 2017; Hubbard, 2009; Twiggs and Collins, 2010). In fact, this decrease in sea level likely also contributed to the changing reef flat morphology, as well as to the diminishing amount of accommodation space (Harris et al., 2015). The facies across the reef flats of both islands show that branching corals are still dominant, though accretion rates have slowed significantly to about 1 mm/yr. This is a MAR that is low, even for global trends during this reef accretion hiatus period (Hynes et al., 2024). During this time, global MAR slows to a mean of 5 mm/yr (or less in some regions), while in the Coastal West Pacific it drops to below 5 mm/yr from 6000 to 3000 YBP. This pattern is present globally, and has been interpreted as a Mid-Holocene Reef Hiatus (Dechnik et al., 2017; Grossman and Fletcher, 2004; Leonard et al., 2020; Toth et al., 2015). A global reef hiatus could be explained by the change in sea level regime, where most regions see a slowdown of sea level rise, often shifting toward a decline in the Mid- to Late-Holocene (Hynes et al., 2024; Khan et al., 2019; Lambeck et al., 2014). Therefore, accretion for Spermonde slowed down well below the global and regional averages. This is due to the fact that by 6000 YBP, with sea level fall there is minimal accommodation space left on the outer margins of the reef flat, with only the middle part of the flat infilling with sediment (Kappelmann et al., 2023; Mann et al., 2019). Sediment transport is still in the seaward to leeward direction, but as we move ever closer to the present there is less room for sediment to be trapped on the flat, and therefore tending to move toward building up the island, and down the leeward slope (Janßen et al., 2017; Kench and Mann, 2017).
By the last few millennia, the island on both platforms has fully formed and is above sea level, continually growing (albeit slowly) due to sediment build up and sea level decline. Samalona island likely fully emerged after the drop in sea level post 5500 YBP (SLF-1 at the top of the core is 4329 YBP, in front of the island). On Kudingareng Keke, we see that the island possibly emerged more recently than Samalona, but certainly the emergence of the sand cay here was also aided by the decline in sea level. The reef flat has all but infilled, and there is stability on the platform of sediment generation, as well as coral growth on the surrounding reef slopes. The age reversal on Kudingareng Keke island is likely due to constant reworking of this part of the island by seasonal changes and sedimentation through erosion and transport (Kench and Mann, 2017). Coral coverage has declined on the flats over the last few decades (Best et al., 1989; Moll and Borel-Best, 1984; Yasir Haya and Fujii, 2017) as accommodation space is less than a meter at points, and between 0 and 1.5 m (MSL) on the majority of the flat. As water depth decreased, there was little room for vertical accretion and therefore coral growth. This decline is also due to the high anthropogenic influence of the population through nutrification and runoff, destructive fishing practices and increased tourism (Yasir Haya and Fujii, 2017). With the crest, flat, and island all being near, at or above sea level, the platforms then begin to prograde on the seaward (Western) slope, and accumulate on the island and eastern edge.
Late-Holocene to recent reef slope development
In the reef slope cores we observed radiometric ages up to 2120 years old on both islands. Samalona slope cores SLS-[1-2] have the oldest dates compared to Kudingareng Keke, but these cores are also up to 1 m longer, and therefore capture more of the reef’s history. These maximum ages are younger than those seen in the reef flat cores, and overlap with the last phase of reef flat development. These observations match a shift to the slope prograding outwards to the seaward slope where there is maximum accommodation space, and the eastern edge now sees increased accumulation, especially on the island (Gischler et al., 2019; Smithers et al., 2006). Mean accretion rates on the slope show continuous sedimentation, and are faster than the reef flat on both islands over the last 2000 years at an average of 2.77 mm/yr (although this is still significantly lower than global and regional averages for this time period). This is to be expected due to there being plentiful accommodation space and higher levels of carbonate production on the reef slope, which matches observations seen elsewhere (Bender et al., 2020; Gischler and Hudson, 2019; Hubbard, 2009; Mann et al., 2019).
Model of island formation
We have summarized these findings in a model of reef formation in the Spermonde (Figure 8). These reefs initiated on top of earlier Cenozoic carbonates, with a mixed coral community consisting of foliose and massive growth forms. Until 5500 YBP, the reefs predominantly accreted to keep up with sea level change. This is documented in our reef cores by comparable accretion rates across the entire reef flat. While Kudingareng Keke and Samalona are relatively small islands, in larger reefs, accretion rates might be more variable resulting in an uneven top of the reef. This is different from the bucket infill model (Harris et al., 2015; O'Leary and Perry, 2010), during which in the initial phase accretion rates are higher around the reef edges, and these are subsequently filled in with sediment once the reef top reaches sea level. In this model, one would expect the oldest ages to be found at the reef edge, and younger sediments in the central part of the reef flat. The slowdown and cessation of sea level rise resulted in a reduction in accommodation space, and an accompanying strong reduction in accretion rates. A large part of this period is associated with an apparent hiatus in reef growth on the reef flat from 6000 to 2000 YBP, irregular morphology, and is driven by limited accommodation space and likely not by environmental conditions (although here we lack sediments of sufficient age on the reef slope to test this). This hiatus, in Spermonde, is also seen globally as a period of minimal reef accretion (Dechnik et al., 2017; Leonard et al., 2020; Perry and Smithers, 2011; Toth et al., 2012). The reef flat is capped by approximately 1 m of post-hiatus sediments. These have predominantly a branching rudstone facies and a coarser grain size, which might indicate a period of continued carbonate production and coral growth on the reef flat and sediment transport across the reef flat resulting in island formation. At the same time, accretion rates on the reef slope are faster than the flat during the Late-Holocene. This represents a shift from accreting to progradation on the western slope/flat, and accumulation on the island and the eastern slope.
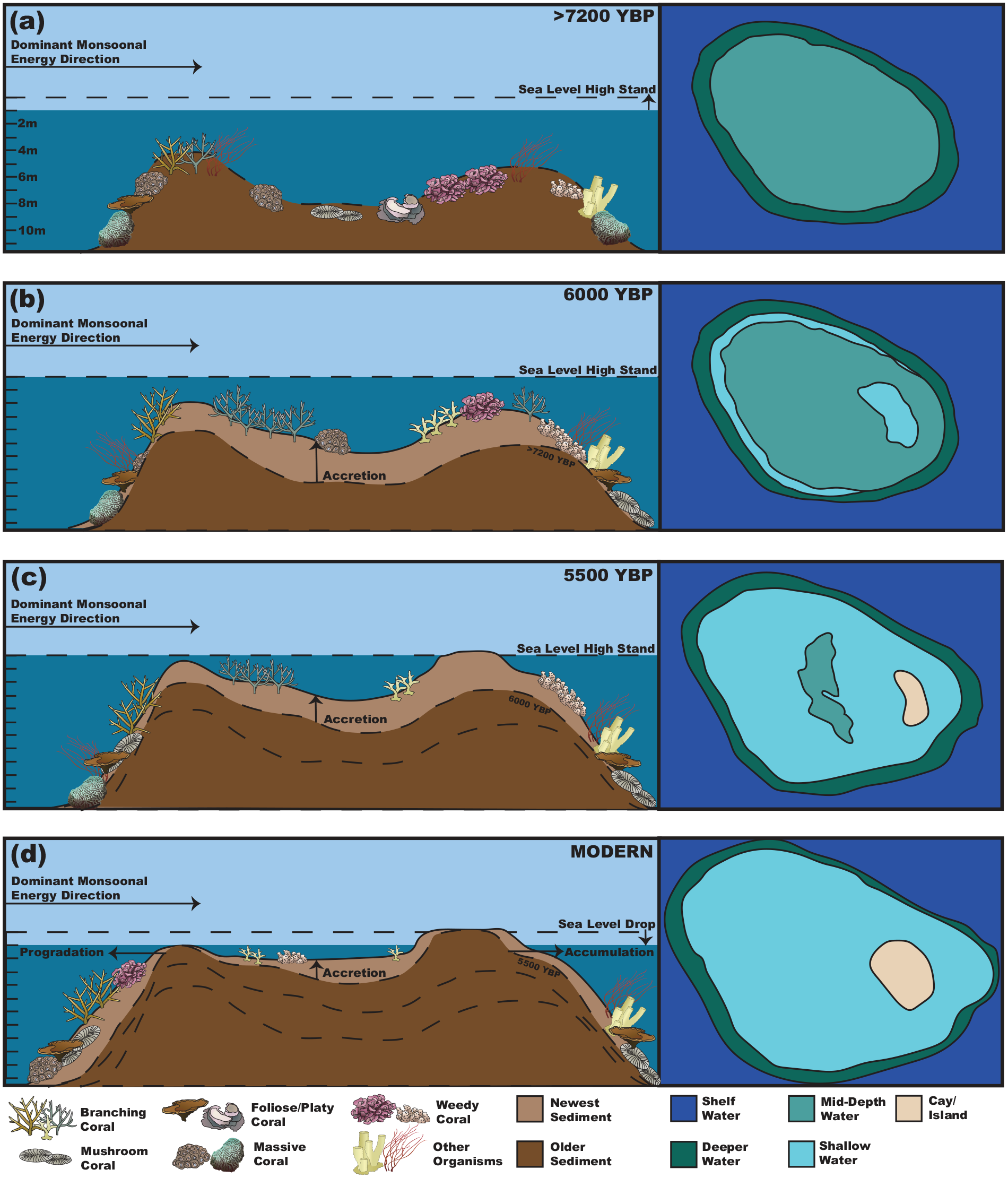
Model of island platform development for the Spermonde Archipelago. (a) Initial reef phase captured at the bottom of the cores. (b) Rapid vertical accretion phase to keep up with the last of the sea level rise regime coming out of the LGM. (c) A sand cay has formed and begun to emerge on the Eastern edge of the reef. (d) Modern day, where with sea level drop the island has grown and expanded due to accumulation of sediment on the island, and the Western margin is prograding. Organism symbols are from Integration and Application Network (IAN) at UCMES (ian.umces.edu/media-library).
Due to Spermonde being tectonically stable (Prasetya et al., 2001) it is clear that sea level is one of the major driving extrinsic factors of patch reef accretion and island formation (Bender et al., 2020; Janßen et al., 2017; Mann et al., 2019). This is seen in the shift from a more mixed coral rudstone facies, to dominantly branching corals, as well as a major reduction in accretion rates coinciding with sea level in the region hitting a Mid-Holocene high-stand and then declining slightly. Traditional models of sand cay formations on (patch) reef complexes include the lagoonal (or bucket) infill model (Harris et al., 2015; O'Leary and Perry, 2010). Another core study from the Spermonde archipelago suggests that a different island, Barrang Lompo, had seen a lagoonal infill from the Mid-Holocene to present where the island now exists (Kappelmann et al., 2023). In this study they also found a hiatus in reef growth from 5700 to 4200 YBP, which is both shorter in duration and occurs earlier than the Mid-Holocene reef growth hiatus observed in other regions and on a global scale (Dechnik et al., 2017; Hynes et al., 2024; Toth et al., 2012). However, this suite of cores was taken from the island itself, and therefore offers no direct insight to reef flat and slope formation. Further, Barrang Lompo is an irregular shaped patch reef complex (possibly formed by the junction of two smaller patch reefs), unlike the more uniformly formed islands such as Samalona and Kudingareng Keke.
In the cores taken in this study from Samalona and Kudingareng Keke, there is no clear evidence of a lagoon on the eastern portion of the island/leeward margin of the platform, though no core was taken on the eastward margin. Further, the reef flat growth hiatus occurs from approximately 6000–1500 YBP, which is more in line with what is observed globally (Hynes et al., 2024), and occurs during a period of consistent declining sea level in the region. What is likely represented by this change in facies and accompanying reef accretion hiatus is due to sea level regimes and the shift from vertical accretion to progradation/accumulation on the patch reef margins (Janßen et al., 2017; Kench and Mann, 2017). In fact, island formation here is more akin to what is seen in the Capricorn bunker group of the Great Barrier Reef (Dechnik et al., 2015, 2016, 2017). This is shown by the reef margins forming first (Figure 8a), followed by the flat accreting vertically from 7200 to 5500 YBP (Figure 8b and c) and subsequently a Mid-Holocene growth hiatus (due to declining sea level). After 5500 YBP the sand cays become sub-aerially exposed, especially after the sea level drop of 0.5–1 m (Figure 8c and d), and reef accretion is now mostly restricted to the slopes, especially on the seaward margin in a progradation phase (Figure 8d). The island and eastern margin see an accumulation of reef sediment transported across the flat from the production zone of the slope and crest. However it should be noted that in Spermonde, subsidence has had minimal impact on reef formation throughout the Holocene, as opposed to the Capricorn bunker group and the Great Barrier Reef as a whole (Crameri and Lithgow-Bertelloni, 2018).
As many islands in the Spermonde Archipelago are heavily populated today and the coral reef complexes surrounding them are a significant economic source for these people, it is pertinent that we understand if these islands can still generate enough sediment and grow at rates fast enough to combat future erosion as sea level regimes change. We show here that in the past, these islands initiated under rapid sea level rise and were able to grow at rates needed to survive and not become “drowned” reefs (Hubbard, 2009). Finally, as there was previously a lack of data on Holocene reef accretion in the Coral Triangle, we can use the MAR calculated here to understand the region’s history, as well as future forecast models of reef accretion in South Sulawesi, and the Coral Triangle.
Conclusion
The islands in the Spermonde Archipelago were heavily influenced by rapid Relative Sea Level rise (RSL) during their formation and subsequent accumulation. During the Early to Mid-Holocene period, it was found that Mean Accretion Rates (MAR) were rapid as reefs were in a catch-up/keep-up phase of accretion. As RSL plateaued in the Mid-Holocene and then declined moving toward the Modern, MAR slowed significantly, especially on the reef flat, as these reefs filled up the remaining accommodation space. This then led to a progradation phase on the slope which is still active today, as reef slope accretion is still within normal ranges. Therefore, island formation did not happen by lagoonal infill (or bucket infill) model, but by continuous accretion controlled extrinsically by sea level.
This shows that islands in the area were able to keep up with varying levels of sea level rise in the past, including the fast rates seen in the Early to Mid-Holocene (Hynes et al., 2024). As significant increases in sea level are expected to occur in the coming decades and centuries, this demonstrates that reefs in the Spermonde Archipelago have the ability to accrete at rates necessary to avoid drowning. However, this assumes that they do not face high levels of additional stressors other than sea level rise (such as thermal stress/bleaching, excess nutrification/eutrophication, and overfishing). As only three sets of core studies for the Spermonde archipelago, including this suite of cores (Kappelmann et al., 2023, 2024), and only one additional core study from the Coral Triangle (Grobe et al., 1985) exist, it would be beneficial to obtain more records of Holocene reef accretion within this region, which is the area of highest coral diversity and growth globally.
Supplemental Material
sj-eps-1-hol-10.1177_09596836251313628 – Supplemental material for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia
Supplemental material, sj-eps-1-hol-10.1177_09596836251313628 for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia by Michael G Hynes, Halwi Masdar, Dedi Parenden, Nicole J de Voogd, Jan-Berend W Stuut, Jamaluddin Jompa, Jody M Webster and Willem Renema in The Holocene
Supplemental Material
sj-eps-2-hol-10.1177_09596836251313628 – Supplemental material for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia
Supplemental material, sj-eps-2-hol-10.1177_09596836251313628 for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia by Michael G Hynes, Halwi Masdar, Dedi Parenden, Nicole J de Voogd, Jan-Berend W Stuut, Jamaluddin Jompa, Jody M Webster and Willem Renema in The Holocene
Supplemental Material
sj-eps-3-hol-10.1177_09596836251313628 – Supplemental material for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia
Supplemental material, sj-eps-3-hol-10.1177_09596836251313628 for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia by Michael G Hynes, Halwi Masdar, Dedi Parenden, Nicole J de Voogd, Jan-Berend W Stuut, Jamaluddin Jompa, Jody M Webster and Willem Renema in The Holocene
Supplemental Material
sj-eps-4-hol-10.1177_09596836251313628 – Supplemental material for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia
Supplemental material, sj-eps-4-hol-10.1177_09596836251313628 for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia by Michael G Hynes, Halwi Masdar, Dedi Parenden, Nicole J de Voogd, Jan-Berend W Stuut, Jamaluddin Jompa, Jody M Webster and Willem Renema in The Holocene
Supplemental Material
sj-eps-5-hol-10.1177_09596836251313628 – Supplemental material for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia
Supplemental material, sj-eps-5-hol-10.1177_09596836251313628 for Impact of Holocene relative sea-level changes on patch reef-island development in the Spermonde Archipelago, South Sulawesi, Indonesia by Michael G Hynes, Halwi Masdar, Dedi Parenden, Nicole J de Voogd, Jan-Berend W Stuut, Jamaluddin Jompa, Jody M Webster and Willem Renema in The Holocene
Footnotes
Acknowledgements
MGH thanks Dr. Elizabeth Eaves for discussion of ideas and language edits. Dr. Singgih Putra and Astrid Verkerk are thanked for their help with collecting cores. Tiar Bahari is thanked for captaining the boats and helping with core collection. Dr. Rohani Ambo-Rappe is thanked for assistance with permits and local logistics. Dino Ramos, Andrew Torres, Daniel Schürholz, and Estradivari are all thanked for help with fieldwork logistics and support. Dr. Bert Hoeksema is thanked for discussion of Spermonde Archipelago coral taxonomy and ecology, as well as fieldwork logistics. MGH would like to thank Dr. Aaron O’Dea for discussion of coring techniques and materials, as well as post collection analysis methods. Dr. Frank Wesselingh is thanked for discussions on mollusk shells and 14C dating. Rob Langelaan and Bertie Joan van Heuven are thanked for assistance with lab analysis and imaging techniques, as well as for obtaining needed lab materials/chemicals.
Author contribution(s)
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was partially funded through Naturalis Biodiversity Center, Leiden, The Netherlands. The fieldwork of this research was predominantly funded by the European Union’s Horizon 2020 Marie Skłodowska-Curie grant agreement (4D-REEF, No. 813360). Work in Indonesia made possible through BRIN Research Permit number 97/SIP/IV/FR/7/2022. Support to JMW was provided by the Hanse-Wissenschaftskolleg (HWK) Institute for Advanced Study, Delmenhorst, Germany.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.