
Abstract
We present a multiproxy analysis of a sediment core from Freshwater Pond, Barbuda, one of just a few inland paleoenvironmental records from the Lesser Antilles. Our results shed light on the relative contributions of climate variability and Pre- and Post-Columbian human activities to vegetation and fire dynamics on Barbuda. The presence of macroscopic charcoal and pollen of ethnobotanically-useful and disturbance-indicator plant taxa in the sediment record suggests that Pre-Columbian subsistence activities occurred within a few kilometers of the pond between ~150 BCE and ~1250 CE. Our record extends anthropogenic fires back into the early Ceramic (500 BCE–1500 CE) and possibly late Archaic Ages (3000–500 BCE) adding evidence to the timing of arrival of the island’s earliest inhabitants. The history of island-wide biomass burning inferred from microscopic charcoal fragments showed heightened fire activity between ~540 and ~1610 CE followed by a period of quiescence that reflected the transition from Pre- to Post-Columbian land-use practices associated with European colonization of the region. The British established a permanent settlement on Barbuda in the 1660s, but given Barbuda’s unsuitability for large-scale agriculture, timber harvesting, small-scale farming, and livestock rearing, activities that left no detectable charcoal footprints likely dominated post-colonial land use. The lack of any clear correspondence between the reconstructed histories of fire and effective moisture at Freshwater Pond supports the idea that Late-Holocene fire activity on Barbuda was driven primarily by human activity.
Keywords
Introduction
The Holocene history of fire activity and its drivers across the lowland landscapes of Caribbean islands has yet to be clearly elucidated (Murphy and Lugo, 1986, 1995; Sánchez-Azofeifa and Portillo-Quintero, 2011) due to a paucity of long fire histories for the region, especially on the smaller islands. Concomitant increases in sedimentary charcoal and the pollen of disturbance and cultivated taxa have been interpreted by some as evidence of human activities (Burney and Burney, 1994; Hooghiemstra et al., 2018; Jones et al., 2018; Kjellmark, 1996; Siegel et al., 2015). Others have pointed to climate as the primary control on fire activity across the Caribbean and tropical central and southern America (Caffrey and Horn, 2015; Marlon et al., 2013; Power et al., 2012). For example, Early- to Mid- and Late-Holocene increases in the sedimentary charcoal content of Laguna Saladilla (Dominican Republic; Caffrey and Horn, 2015), Laguna Tortuguero (Puerto Rico; Burney and Burney, 1994), and Lake Miragoâne (Haiti; Higuera-Gundy et al., 1999) were attributed to enhanced fire activity caused by increasing winter insolation (Caffrey et al., 2015; Caffrey and Horn, 2015). Disentangling charcoal that originated in fires started by the earliest human settlers of the Greater Antilles (~5000 BCE; Napolitano et al., 2019) from fires started by lightning is impossible without archaeological or other long records of human activity from the same area. Thus, some interpretations may underestimate the contribution of anthropogenic fires to the overall sedimentary charcoal record during the Pre-Columbian period. By contrast, interpretations of ~9000-year old sediment records from Peten, Guatemala, draw on a combination of climatic and anthropogenic factors to explain the history of fire (Anderson and Wahl, 2016). Those sediment records (Anderson and Wahl, 2016) provided a rare window into the Holocene fire history for the Maya lowlands, during which fire activity shifted from infrequent, climate-driven fires during the early Holocene, to more-frequent anthropogenic fires during the Mid-Holocene, following settlement by sedentary agriculturalists.
Studies of sedimentary charcoal influx from a range of elevations and forest types across the insular and mainland Caribbean have shown decreases in charcoal influx after ~1450 CE that coincide broadly with the onset of the Little Ice Age (LIA, ~1400–1850 CE; Dull et al., 2010; Nevle et al., 2011; Power et al., 2012), suggesting a link between natural climate variability and fire activity. While some proxy records from the circum-Caribbean region show evidence of relatively arid conditions during the LIA (Hodell et al., 2005; Lane et al., 2011), others have proposed that the region was not uniformly dry, but rather experienced greater spatial and temporal variability in effective moisture (Burn et al., 2016; Burn and Palmer, 2014; Fensterer et al., 2012; Holmes et al., 2023). Given the spatial and temporal complexity of precipitation across the Caribbean during the LIA, climate variability alone seems an incomplete explanation for the uniform decrease in biomass burning across the Caribbean during that period.
In contrast to the climate-driven hypothesis for reduced biomass burning during the LIA, Dull et al. (2010) and Nevle et al. (2011) invoked a population collapse hypothesis that proposed the spread of regional pandemics throughout the Americas after first European contact leading to rapid declines in indigenous populations, which in turn, reduced anthropogenic fire ignitions in tropical forest communities. Reforestation following the indigenous population collapse may have transformed the region from a carbon source to a carbon sink, explaining, in part, the decrease in atmospheric CO2 during the LIA (Dull et al., 2010; Nevle et al., 2011). That hypothesis conceptualized decreased anthropogenic burning and increased carbon sequestration as direct consequences of colonization that in turn became significant climate forcing mechanisms for the LIA (Dull et al., 2010).
The attribution of biomass burning to human ignitions rather than climate variability appears to also hold true for forest communities of the Lesser Antilles. Siegel et al. (2015) demonstrated the impact of indigenous subsistence activities on Antillean fire regimes and vegetation, and further, pointed to the need for paired paleoenvironmental and archaeological studies. Multiproxy analyses of a suite of sediment cores from the southern islands of the Lesser Antilles (Trinidad, Grenada, Martinique, and Guadeloupe) documented the co-occurrence of sedimentary pollen and spores of ethnobotanically significant and disturbance taxa as well as sustained increases in microcharcoal particles (Fitzpatrick and Keegan, 2007). The datasets showed evidence of landscape transformation associated with the initial human colonization of the Lesser Antilles by 4000–2500 years BCE.
The variable scientific explanations for long-term fire patterns in the Caribbean region, some emphasizing human activities (Mueller et al., 2018; Siegel et al., 2015) and others climatic causes, highlight the need for new studies focused on Holocene interconnections between climate, human activities, and fire. In this paper we examine the relative roles of climatic variability and human activity as drivers of fire and vegetation dynamics during the Mid-Late-Holocene on the island of Barbuda in the northeastern Caribbean. We reconstructed the long-term environmental history of Barbuda from micro- and macro-charcoal fragments, pollen, and spores preserved in the sediments of Freshwater Pond and placed in the context of recent paleoclimatic (Burn et al., 2016) and archaeological research (Bain et al., 2017; Perdikaris and Boger, 2023; Rousseau et al., 2017). The island of Barbuda, with its rich history of scholarship (Bain et al., 2017; Berland et al., 2013; Boger et al., 2014; Burn et al., 2016; Faucher et al., 2017; Perdikaris and Boger, 2023; Perdikaris et al., 2017; Rousseau et al., 2017; Watters,1980a, 1980b; Watters et al., 1992) provides an ideal study site to examine the links between Late-Holocene environmental changes and its increasingly well-documented human history.
Study area: Climate, vegetation, and human history
Barbuda (17°38′1″N 61°48′30″W, Figure 1), a small (~160 sq. km) low-lying limestone island, lies on the northeastern edge of the Lesser Antilles island arc, which separates the Caribbean Sea from the Atlantic Ocean (Brasier and Donahue, 1985). The island comprises Holocene and Pleistocene limestone beach ridges covered mostly by carbonate sands and thin clays. The Highlands, a plateau (~45 m a.s.l.) of older Pliocene limestone deposits (Brasier, 1975) constitutes the island’s highest topography occupying a large area of the island’s east-central island zone. Codrington Lagoon dominates the northwestern portion of the island, which is characterized by a relatively flat coastal plain punctuated with fresh- to saline-water ponds, lagoons, and wetlands.
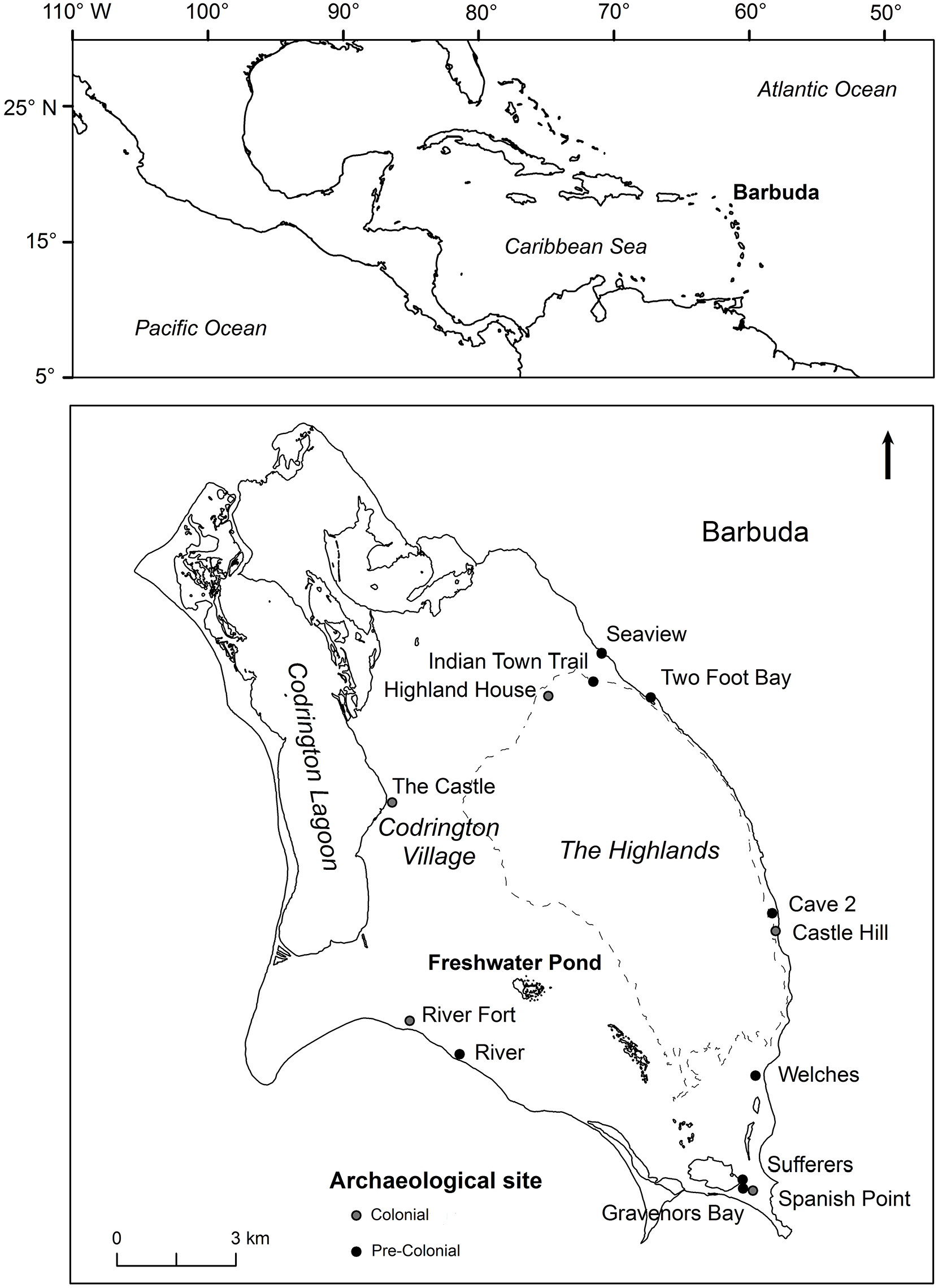
Top: Location of Barbuda within the northeastern Lesser Antilles. Bottom: Location of Freshwater Pond and prehistoric (black circles) and historical (gray circles) archaeological sites on the island. The dashed lines denote the approximate boundary of the Highlands.
Barbuda is one of the driest Caribbean islands (Jackson, 2001), owing to its flat topography and consistent sinking air from the Bermuda High, a semi-permanent high-pressure system that dominates climate during the winter dry season. Annual temperatures ranged from 24°C to 29°C and precipitation averages were 882 mm from 1965 to 2000; 10 of those 35 years were designated as “meteorological” droughts (Jackson, 2001). About half of Barbuda’s annual rainfall occurs during the wet season (May–November) with ~20% falling during the January–April dry season. Tropical storms and cyclones contribute substantially to annual precipitation totals, creating considerable interannual variability in yearly rainfall totals (Jones et al., 2018). Climatic variability on interannual to millennial timescales is influenced by El Niño-Southern Oscillation (ENSO), the Atlantic Multidecadal Oscillation (AMO), North Atlantic Oscillation (NAO), and migration of the Intertropical Convergence Zone (ITCZ) (Burn et al., 2023; Fensterer et al., 2012; Haug et al., 2001; Peros et al., 2015).
Barbuda’s vegetation is dominated by secondary growth that is the footprint of a long history of clearance, charcoal production, timber removal, and wind disturbance from repeated tropical storms (Bain et al., 2017, Faucher, 2018; Tweedy, 1981). From the 1660s onwards, land-use activities included open-range cattle and goat grazing and small-scale farming. The lowlands are characterized by a mixed deciduous-evergreen xeric woodland (Francis et al., 1994; Lindsay and Horwith, 1997) of low canopy height (3–10 m) with constituent species that have developed common xerophytic adaptations including densely-packed gnarled trees and shrubs, often with spines and hard evergreen leaves of markedly reduced size. Isolated tree genera include Haematoxylon, Tabebuia, Bucida, Plumeria, Canella, Pisonia, and Ficus citrifolia, the only native/naturalized species of the Moraceae family. Representatives of the Fabaceae (legumes) family are abundant as shrubs and small trees (e.g. Acacia, Caesalpinia, Pithecellobium, and Prosopis). Important non-Fabaceae shrub genera include Croton, Coccoloba, Comocladia, Lantana, and Solanum. Succulents such as Pilosocereus (tree-form), Opuntia, and Agave occur in the woodland undergrowth and on cliffs and rocky exposures in the Highlands.
The Highlands and neighboring low-lying areas support mixed evergreen-deciduous forests (Francis et al., 1994; Lindsay and Horwith, 1997), with emergents (e.g. Coccothrinax barbadensis, Bursera simaruba, Tabebuia heterophylla, and Ficus citrifolia) rising above a 10–15 m canopy. Canopy elements include Tabebuia, Bucida, Pisonia, Zanthoxylum, and Sideroxylon, while species of Canella, Capparis, Coccoloba, Eugenia, Malphigia, and Plumeria represent common understory taxa. Isolated stands of broad-leaved evergreen forest containing the palm Coccothrinax barbadensis, Ficus citrifolia, and species of Sideroxylon, Capparis, Canella occupy solution holes of the Highlands with reliable access to water (Francis et al., 1994; Lindsay and Horwith, 1997).
Coastal and wetland areas are dominated by seasonally flooded/saturated broad-leaved evergreen sclerophyllous forests dominated by mangrove species, including red (Rhizophora mangle), white (Laguncularia racemosa), black (Avicennia germinans), and button mangroves (Conocarpus erectus) (Stoddart et al., 1973; Lindsay and Horwith, 1997), and Annona glabra. Grasses, sedges, and the giant leather fern Acrostichum danaefolium, are important vegetation components. Aquatic plant genera such as Utricularia, Ceratophyllum, Nymphaea, Limnobium, and Ruppia and algal taxa (e.g. Chara, Nitella), are common members of submerged and floating plant communities in freshwater ponds (Lindsay and Horwith, 1997).
Barbuda’s history of human settlement spans the Archaic (~3000–500 BCE) through post-Colonial periods (Bain et al., 2017; Rousseau et al., 2017). The earliest Archaic Age peoples practiced hunter-gatherer activities, such as marine resource extraction and foraging for wild plants (Rousseau et al., 2017), while the Late Ceramic Age (600–1500 CE) peoples conducted shifting (slash and burn) agriculture, resulting in forest clearance and burning of the residual biomass (Bain et al., 2017). Carbonized remains of tropical dry forest and coastal taxa, such as lignum-vitae (Guaiacum officinale), cedar (Tabebuia), and mangrove species (Avicennia germinans, Conocarpus erectus, Laguncularia racemosa), identified from Ceramic Age archaeological microcharcoal samples (Bain et al., 2017), confirm that indigenous population exploited these plants for fuel wood and construction materials (Francis et al., 1994; Harris, 1965).
Complex social and political changes that occurred during the late Ceramic Age, 600–1500 CE, resulted in the Taíno polities that characterized the Greater Antilles at the time of European colonization (Newsom and Wing, 2004; Wilson, 2007). After ~1300 CE, many of the Leeward Islands, including Barbuda, appear to have either been abandoned or were sparsely populated by indigenous populations (Bain et al., 2017; Wilson, 2007). While initial attempts of European colonization failed, by the 1670s, Barbuda was exporting livestock including horses, cattle, sheep, and goats (Bain et al., 2017; Harris, 1965). In contrast to its higher-relief sister island Antigua (~62 km to the south), infertile soils and low precipitation prevented the establishment of plantation agriculture on Barbuda (Harris, 1965; Watters, 1980b). In the Colonial economy, Barbuda’s role was that of a provisioning island that produced wood products and a variety of foodstuffs for Antiguan plantations, grown by enslaved peoples. Most inhabitants and cultivated lands on Barbuda were limited to the vicinity of Codrington village allowing for the practice of open-range cattle herding. The current population of about 1700 lives clustered in and around the village of Codrington resulting in relatively well-preserved archaeological heritage across the island (Rousseau et al., 2017). Small-scale cultivation and some livestock browsing are still practiced on the island (Perdikaris and Boger, 2023; Potter and Sluyter, 2010).
The study site, Freshwater Pond (17.600754°N, 61.793873°E; 6 m a.s.l.; Figure 1), is a permanent, shallow (<1 m), flat-bedded, fresh to brackish water lake situated on the Codrington Limestone Group in southwestern Barbuda. The pond is hydrologically closed, with water levels tied to precipitation and groundwater flow, which vary seasonally. During the boreal summer (wet season) the lake can treble its surface area; in contrast, during the dry winter months, the lake contracts and water salinity increases (Burn et al., 2016). A seasonally flooded/saturated mangrove forest association (Lindsay and Horwith, 1997), with the mangroves Rhizophora mangle, Avicennia germinans, Laguncularia racemosa, and Conocarpus erectus, Annona glabra (pond apple), the giant leather fern, Acrostichum danaeifolium, and Chara growing within the pond itself (Burn et al., 2016). In the drier, and slightly higher elevations, east of the pond and adjacent wetlands, the vegetation transitions abruptly to mixed evergreen-deciduous xeric woodland.
Methods and materials
In January 2010, we recovered sediment core FP2 (0–84 cm) from ~1 m of water using a Colinvaux-Vohnout drop-hammer modified piston corer (Colinvaux et al., 1999). The core was extruded in the field, cut into 1 cm slices, and stored at 4°C at the University of the West Indies. Counts of the fossil gastropod and ostracod contents of the sediment record were combined with analyses of the stable isotopic composition of ostracod shells and are described in detail by Burn et al. (2016). Loss-on-ignition (LOI), charcoal, and pollen analyses were performed on samples shipped to the Paleoenvironments Laboratory at Virginia Tech. Four Accelerator Mass Spectrometry (AMS) radiocarbon dates recovered from plant macrofossils provided chronological control for the FP2 core (Table 1) and were fully documented in Burn et al. (2016).
FP2 (0–84 cm) radiocarbon date results through the facilities at the National Ocean Sciences Accelerator Mass Spectrometry Facility (NOMAS) at Woods Hole Oceanographic Institution and the Scottish Universities Environmental Research Centre (SUERC) at the University of Glasgow.
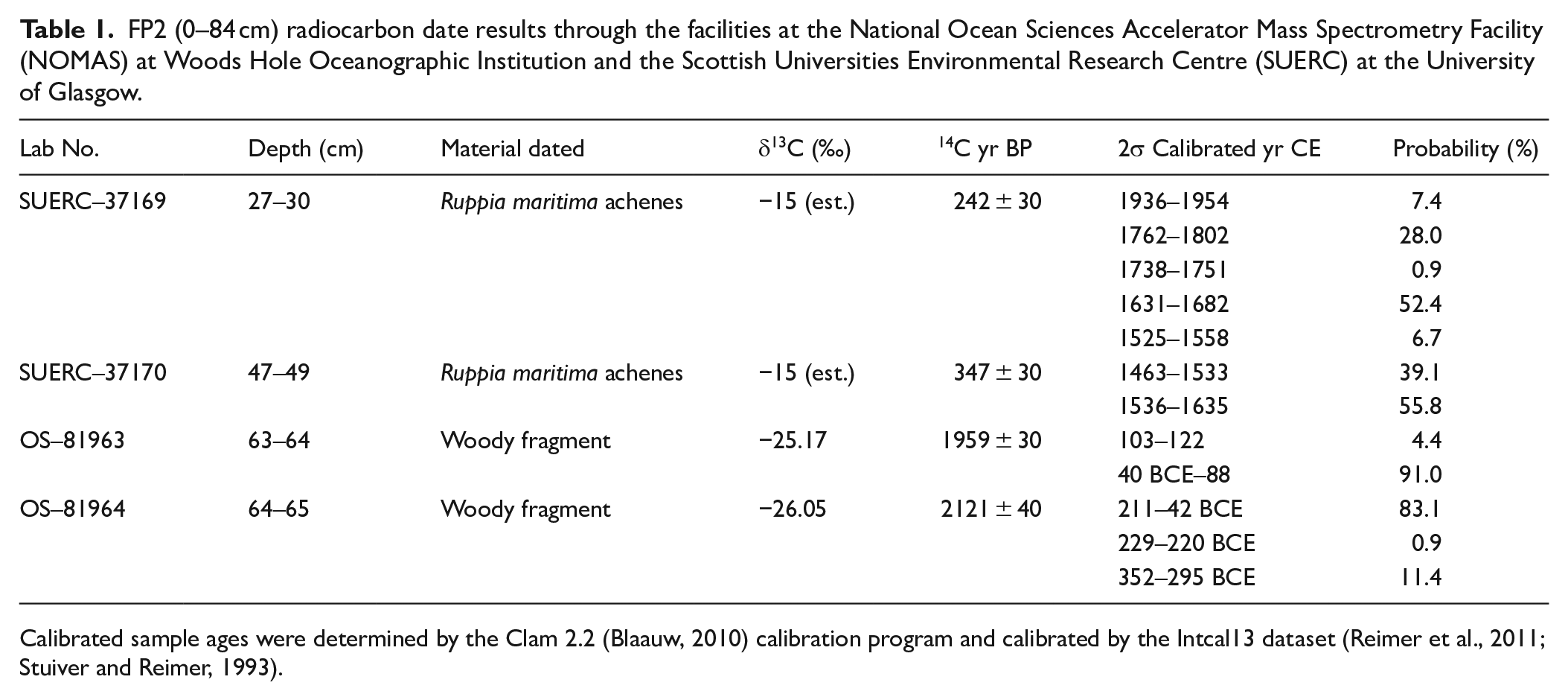
Calibrated sample ages were determined by the Clam 2.2 (Blaauw, 2010) calibration program and calibrated by the Intcal13 dataset (Reimer et al., 2011; Stuiver and Reimer, 1993).
We conducted loss-on-ignition analysis (LOI, Heiri et al., 2001) at 1 cm intervals along the entire core to determine sediment organic and inorganic carbon content. Samples were dried overnight at 100°C to determine water content, then combusted at 550°C for 2 h to determine organic matter loss, and at 1000°C for 1 h to determine the carbonate fraction. To reconstruct local fire history, we isolated macroscopic charcoal (>125 µm; Long et al., 1998; Whitlock and Anderson, 2003; Whitlock and Larsen, 2001) from 1 cm intervals for the entirety of the sediment core. We soaked 1 cm3 samples of sediment in 5% sodium hexametaphosphate to disperse clays (Whitlock and Anderson, 2003; Whitlock and Larsen, 2001) and gently sieved samples through 125 µm screens. Charcoal particles left on top of the screens were identified and tallied in gridded petri dishes under low magnification using a Leica stereozoom microscope. Depth levels 34–35, 49–50, and 53–54 cm did not contain sufficient sediment (used up in prior analyses) for macrocharcoal analysis but the presence of fragments >125 µm were noted from pollen slide scans for these depths. We also quantified microscopic (>10–125 µm) charcoal in pollen slides to reconstruct the extra-local to regional fire history (Patterson et al., 1987; Whitlock and Anderson, 2003; Whitlock and Larsen, 2001). For those slides rejected for pollen analysis due to low abundance, microcharcoal fragments and control spores were counted until a total of 200 control spores and charcoal particles were tallied (Finsinger and Tinner, 2005).
We processed 14 sample depths for pollen and spores based upon LOI results and observed changes in stratigraphy in the sediment core. We modified the standard pollen preparation technique of Faegri and Iverson (1989) to account for the high clay content of the sediment using the following treatments: 10% hydrochloric acid for the removal of carbonates, 10% potassium hydroxide to break up organics and for the removal of humic acids, 10% sodium hexametaphosphate to disperse clays, and 49% hydrofluoric acid for the removal of silicates (Traverse, 2007). Prior to processing, we added a known concentration of Lycopodium spores (one tablet containing 18,584 ± 371 spores) to each sample to allow calculation of pollen and spore concentrations (Stockmarr, 1971). Due to the relative scarcity of preserved pollen and spores within the samples, we lowered our usual pollen sum to a minimum of 200 pollen grains and spores (excluding indeterminate pollen/spores and fungal spores) per slide at 400× magnification using a Leica compound light microscope. We tallied indeterminate grains and classified them as either damaged or obscured.
We identified pollen and spores using reference slides, published reference collections (Colinvaux et al., 1999; Hooghiemstra, 1984; Roubik and Moreno, 1991; Snyder et al., 2007; Willard et al., 2004), and online pollen databases (Bush and Weng, 2006; Davis, 2001; Rowe, 2006; Snyder et al., 2007). To aid in interpretation of the pollen spectra, we grouped pollen types and spores (excluding indeterminate pollen) into forest and shrub or wetland and herbaceous vegetation categories based on the ecology of the taxa (Francis et al., 1994; Harris, 1965; Smith et al., 2004). We plotted and analyzed pollen/spore and charcoal data using the pollen plotting software TILIA v. 2.0.41 (Grimm, 1990).
Results
Core description and chronology
The basal unit (84–50 cm) was composed of greenish-gray clays overlain by 50 cm of authigenic marl rich in carbonate micro- and macro-fossils including fresh- to brackish-water ostracods, gastropods, and charophyte oospores. From 84–72 cm, the greenish-gray clay contained mangrove detritus and limestone gravel. Clays from 72–50 cm contained iron-rich red mottles, with a sharp transition from basal clays to marl at 50 cm. This abrupt change is reflected in the LOI-derived carbonate fraction, which sharply increases from an average of 8% in the basal clays (84–50 cm) to 27% in the upper marls (50–0 cm). Organic content also increased up-core, from ~5% to ~21% (Figure 2).
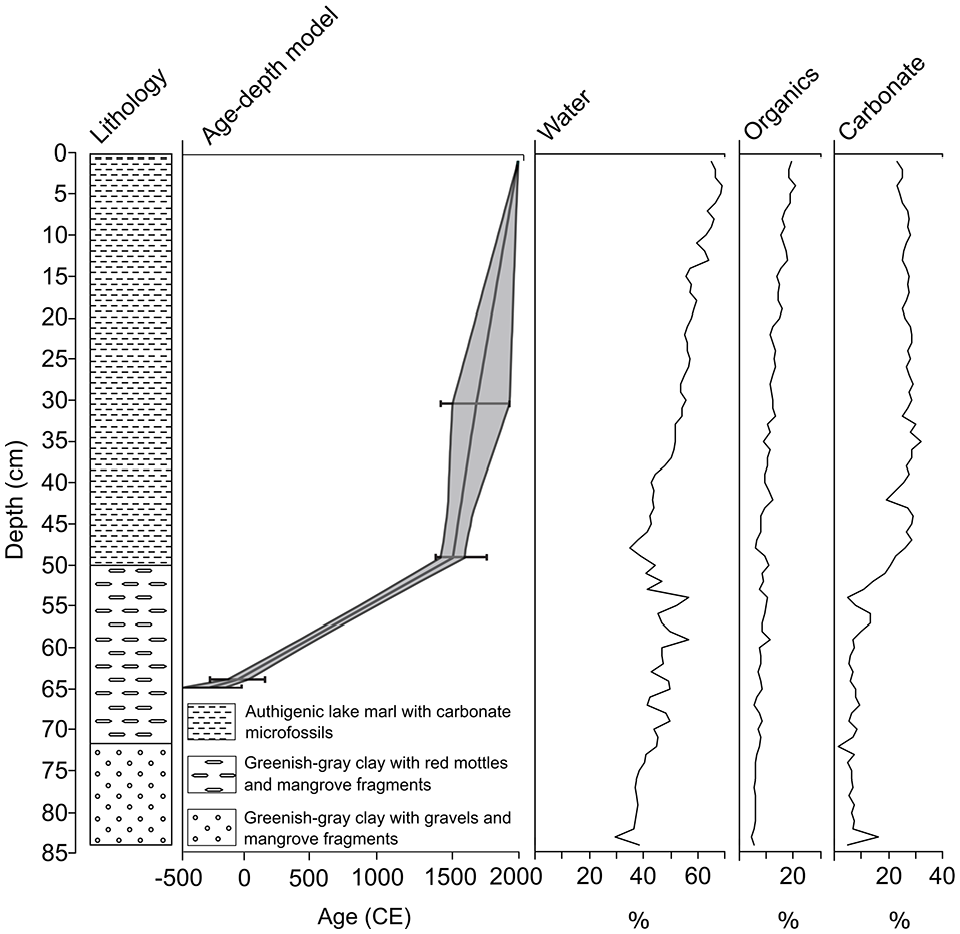
Lithology, age-depth model, and loss-on-ignition data for sediment core FP2.
The basal radiocarbon date at 65 cm is evidence of the initiation of sediment collection at the core site over 2000 years ago, by at least ~150 BCE. The sediments below that date deposited from 66–84 cm likely represent the early Ceramic (500 BCE–1500 CE) and possibly Archaic (3000–500 BCE) Ages. Thus, we include the Archaic Age under the category of “Cultural Age” of Figure 3, recognizing that this designation is for general comparison of our record to archaeological studies (Rousseau et al., 2017; Watters et al., 1992) from the island.
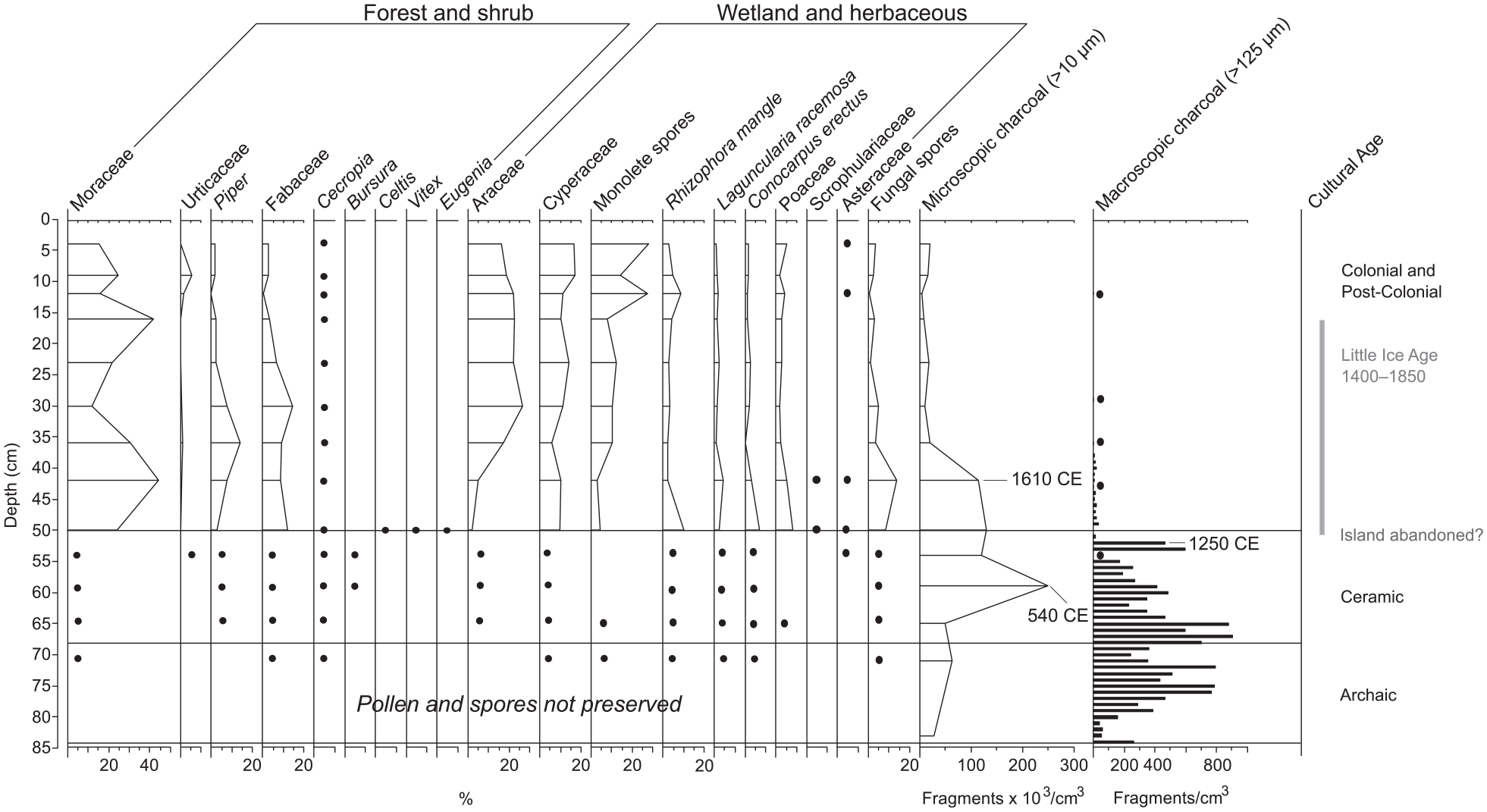
Pollen, spores, and charcoal data from the FP2 sediment core. Black circles denote presence of pollen, spores, or macroscopic charcoal for depth levels for which we could not perform full counts. Horizontal gray bars represent periods of interpreted drought from previous analysis of this sediment core (Burn et al., 2016). Horizontal black lines note boundaries of cultural periods, labeled on the right. A basal radiocarbon date at 65 cm precludes the establishment of a chronology for the entirety of our sediment core, but it is likely that sediments deposited from 66–84 cm represent the early Ceramic (500 BCE–1500 CE) and possible later Archaic (3000–500 BCE) Ages.
Microcharcoal and pollen stratigraphy
All 14 sample depths processed for pollen analysis contained microcharcoal particles (Figure 3) with concentrations ranging from a minimum of 4762 fragments/cm3 deposited at 11–12 cm to a maximum of 249,557 fragments/cm3 deposited at 58–59 cm. Charcoal fragments were moderately abundant from the bottom of the core (84 cm) to about 64–65 cm (mean of 47,187 fragments/cm3), after which concentrations increased dramatically and remained high until 41–42 cm (mean of 153,386 fragments/cm3). Above 41 cm depth, microcharcoal concentrations decreased and remained low (average of 13,777 fragments/cm3) to the top of the core.
Sediment samples contained pollen from 38 families and spores of Pteridophytes and spike moss (Selaginella). Pteridophyte spores were dominated by monolete, psilate types, likely Polypodiaceae. Pollen and spores were sufficiently preserved for analysis in nine levels between 0 and 50 cm (Figure 3). They were not preserved in the basal sediments (82–84 cm) and poorly preserved until the stratigraphic change from greenish-gray clay to authigenic marl at 50 cm. Low concentrations of pollen of mangroves (Conocarpus erectus, Laguncularia racemosa, Rhizophora mangle), sedges, and Fabaceae, and fern spores, in sediments deposited 70–71 cm indicated the establishment of a shallow mangrove wetland and surrounding vegetation by at least ~ 150 BCE. Damaged (oxidized, crumpled, and torn) pollen grains and spores, constituting the majority of unidentifiable palynomorphs, remained high in sediments deposited above 50 cm, accounting for 34–60% (maximum at 49‒50 cm) of sporopollenin on slides, calculated as a ratio of indeterminate pollen grains to combined indeterminate and identifiable pollen and spore count.
The pollen and spore record was heavily dominated by ferns and herbaceous taxa, including Araceae, Cyperaceae, and Poaceae. Other important pollen types included white (Laguncularia racemosa) and red mangroves (Rhizophora mangle), and the terrestrial mangrove, buttonwood (Conocarpus erectus). Important arboreal and shrub elements included Moraceae (probably Ficus citrifolia), palms, and Fabaceae, primarily Acacia and Mimosa. Pollen of Cecropia, (probably C. schreberiana; Francis et al., 1994; Lindsay and Horwith, 1997), a pioneer tree common in disturbed forested habitats and clearings throughout the tropics, was present in all depth levels processed for pollen analysis. Pollen of Piper, a genus often found in disturbed vegetation of the Neotropics (Smith et al., 2004), contributed substantially to some depth levels. Edible plants were sparsely represented in the Freshwater Pond record, but included the families of Cucurbitaceae (gourds) and Dioscoreaceae (yam). Pollen of Solanaceae, the nightshade family, includes edible (e.g. Capsicum and species of Solanum) and medicinal plants (Smith et al., 2004) and was treated as an ethnobotanically useful taxon by Siegel et al. (2015), so we treated them as such here.
Macroscopic charcoal
Macrocharcoal concentrations (Figure 3) varied greatly, with a maximum concentration of 905 fragments/cm3 deposited at 66–67 cm. The basal sediments (84–52 cm) contained the highest amount of macrocharcoal (average 415 fragments/cm3), above which concentrations decreased from 51–37 cm (maximum 30 particles/cm3). Macrocharcoal occurred at low concentrations in the upper sediments of the core, with single fragments appearing at 35–36, 28–29, and 11–12 cm. Macrocharcoal morphology exhibited a change at around 51 cm. Charcoal fragments in sediments deposited from 84–52 cm exhibited various morphotypes, including longer and thicker pieces with, and without branches; segmented, latticed polygons; and blocky/rectangular particles with no structure, as defined by Enache and Cumming (2006, 2007) and Mustaphi and Pisaric (2014). Above 52 cm, macrocharcoal fragments exhibited only one morphotype: shorter blocky/rectangular structure with no branches. Charred mangrove plant macrofossils were observed in samples in basal sediments below 52 cm.
Discussion
Mid- to Late-Holocene fire history of Barbuda
Microcharcoal particles are often dispersed high into the atmosphere during fires promoting long distance dispersal. Their presence in lacustrine sediments is typically interpreted to infer fires burning within 20–100 km of the study site (Conedera et al., 2009; MacDonald et al., 1991; Tinner et al., 1998). Our interpretation of the microcharcoal record from Freshwater Pond in Barbuda is constrained by the island’s isolated location on the northeastern edge of the Lesser Antilles (Figure 1) and the prevailing northeasterly trade winds. These two factors largely preclude the deposition of small charcoal particles derived from fires burning on surrounding islands. Historical volcanic eruptions of the Soufrière Hills volcano of Montserrat, ~100 km to the southwest, may have contributed to fire events on the island in the past, but we found no evidence of tephra in the sediment record at Freshwater Pond, nor evidence that the recent eruptions of Montserrat (1995 onwards) led to increases in sedimentary charcoal. These factors combined simplify our interpretation of the microcharcoal content of our sediment core as largely produced from fire activity on the island.
Burning on Barbuda during the late Archaic through early Ceramic Ages, a period of relatively aridity across much of the Caribbean (Haug et al., 2001; Higuera-Gundy et al., 1999, Hodell et al., 1991; Lane et al., 2011), was represented by moderately abundant microcharcoal fragments in the basal sediments, relative to the rest of the core (Figure 3). Microcharcoal abundance peaked at ~540 CE and remained high until ~1610 CE (Figure 3), a result in general agreement with the regional-scale fire reconstructions of Dull et al., (2010) and Nevle et al., (2011). The substantial increase in biomass burning at ~540 CE occurred after the first known permanent settlement at the Seaview archaeological site on Barbuda (~100 BCE, Bain et al., 2017) and coincides with the beginning of the Ceramic Age cultural period, characterized by the arrival of sedentary horticulturalists who practiced slash and burn agriculture and lived in large settlements across the Caribbean (Wilson, 2007). On Barbuda, this is consistent with the migration of Ceramic Age agriculturalists from northern South America through the Lesser Antilles between 3000 and 2000 years ago (Fitzpatrick, 2015; Wilson, 2007).
Archaeological investigations on Barbuda dating to the Ceramic Age support our assertion that the Freshwater Pond charcoal record primarily documents burning by indigenous peoples on the island. Increased charcoal from ~540–1610 CE coincides with occupation of the Seaview (100 BCE–650 CE), Indian Town Trail (1130 CE), and Sufferers (550–900 CE) archaeological sites (Figure 1; Bain et al., 2017). Further, large charcoal fragments (>4 mm) of Guaiacum officinale, Eugenia sp., Exostema caribaeum, and Amyris sp. found at the Seaview and Indian Town Trails sites provide evidence of the indigenous use of woody species for timber, fuel, and food (Bain et al., 2017), but also supporting evidence of anthropogenic disturbance to Barbuda’s terrestrial ecosystems. Consistently low concentrations of microcharcoal deposited ~1660 CE through the post-Colonial period likely reflect a change from indigenous to Colonial land use activities the latter of which no longer produced a significant charcoal signature. British managers and a population of enslaved peoples practiced subsistence agriculture in areas near Codrington village, with additional environmental impacts resulting from the removal and export of timber and livestock rearing (Bain et al., 2017; Harris, 1965).
Our contention that the Freshwater Pond charcoal record largely reflects anthropogenic burning, with decreased burning during the Colonial Period linked to changes in land use, is consistent with the population collapse hypothesis of Dull et al. (2010) and Nevle et al. (2011) that attributed substantial decreases in fire activity following European colonization in Central America and Amazonia to the subsequent demise of indigenous populations. Interestingly, the Freshwater Pond charcoal record does not co-vary significantly with the stable isotope record of extended drought episodes during the pre- and post-Colonial periods (Burn et al., 2016) (Figure 3, gray bars). This lack of correlation between the charcoal and paleoclimate records suggests that fire activity on the island resulted primarily from human activities. Similarly, Hooghiemstra et al. (2018) documented the different impacts of pre- and post-Columbian land use on fire regimes in the Yaque River Valley on the island of Hispaniola (northern Dominican Republic) In that study, a significant decrease in sedimentary charcoal by ~1650 CE was linked to the replacement of small-scale agriculture practiced by the indigenous Taino peoples by larger-scale farming activities, including deforestation of landscapes for cattle grazing. Further reductions in sedimentary charcoal in the Yaque Valley occurred after ~1740 CE when large-scale deforestation was replaced by selective timber cutting and removal (Hooghiemstra et al., 2018).
Over the past four centuries the eastern Caribbean region has experienced significant climatic variability including multiple droughts (Berland et al., 2013, Burn et al., 2016). During this period, European settlers introduced subsistence agriculture to Barbuda, but the primary environmental impacts of settlement stemmed from the introduction of goats and cattle and the extensive removal and export of timber that together greatly diminished woody plant recruitment and growth and thus reduced the local availability of fuel for fire (Bain et al., 2017). The pollen record of Freshwater Pond corroborates historical records of natural resource exploitation on the island. We interpret peaks in Piper pollen and the presence of Cecropia pollen as indications of vegetation clearance, probably of forests growing in and near the Highlands, around the time of the first permanent English settlement (Figure 3). While humans can alone greatly reduce biomass, the combination of human impacts and frequent or extended droughts, such as those recorded at Freshwater Pond (Burn et al., 2016), would have further reduced available fuels, the extent and intensity of fires, and consequently the abundance of charcoal in our sediment record.
Pre-Columbian land use near Freshwater Pond
Relatively high concentrations of sedimentary macrocharcoal in Freshwater Pond sediments spanning the period ~150 BCE to ~1250 CE (Figure 3) support the archaeological interpretations of burning by pre-Columbian settlers (Bain et al., 2017; Watters et al., 1992). Most charcoal fragments fell into the >125–250 μm size class suggesting deposition within a few 100 m of a fire, but Whitlock and Millspaugh (1996) showed that in the western USA, macrocharcoal particles in this smallest size class were sometimes wind-transported for several kilometers. While studies of charcoal taphonomy in the Caribbean are lacking, we argue that consistently strong easterly trade winds and onshore sea breezes could have transported macrocharcoal fragments from fires that originated in and around the southern portions of the Highlands (Figure 1), only a few kilometers to the east of Freshwater Pond and upslope. Thus, we interpret our macrocharcoal record to represent a combination of in-situ burning of vegetation within the pond’s immediate catchment, fire activity along the southwestern flanks of the Highlands, and possibly some that may have originated from within the island’s archaeological sites.
Barbuda’s southwestern coast was at least seasonally occupied by Archaic Age (3000–500 BCE) peoples who fished for Queen conch (Lobatus gigas; formerly Strombus gigas) discarding the shells onto a linear midden deposit known today as the Strombus Line (Rousseau et al., 2017; Watters, 1980a; Watters et al., 1992). A recent reconsideration of radiometric dates from the Caribbean region confirms Barbuda’s settlement between 4000 and 3000 CE (Napolitano et al., 2019). Shell midden and stone knapping sites, just ~2 km from Freshwater Pond, would have supplied pre-Columbian settlers with lowland and mangrove forest resources exploited for food, medicine, fuel, and construction materials (Bain et al., 2017; Fitzpatrick, 2015; Newsom and Wing, 2004; Wilson, 2007).
The landscape surrounding Freshwater Pond and other low-lying areas along the southwestern edge of the Highlands contain moist soils throughout much of the year and some of the best available land for cultivation by Ceramic Age peoples, especially during the dry season or extended droughts. Recent archaeobotanical research (Faucher, 2018) did not report evidence of manioc or maize cultivation, but studies of starch grains and phytoliths from the Ceramic Age Seaview and Indian Town Trail sites revealed the presence of Maranta spp. (arrowroot) and Fabaceae. Evidence of manioc cultivation is rare across the Caribbean (Fitzpatrick, 2015). We found only limited sedimentary pollen evidence of edible plants of the Solanaceae family and no evidence of the cultivated plants, maize (Zea mays) and manioc (Manihot esculenta). Less than optimal pollen preservation in sections of the sediment core reduces the likelihood of finding crop pollen grains in the Freshwater Pond core, a problem magnified by other factors related to their specific taphonomies (Campbell, 1999). Due to their large size and/or the insect-pollination syndromes of parent plants, pollen grains of many cultivated plants, including maize, manioc, most cucurbits (squash), and yams, are poorly dispersed and produced only in low abundance rendering them poorly represented in pollen-based reconstructions (Lane et al., 2010; Whitney et al., 2012). In fact, during standard pollen counts, they are nearly always found as single pollen grains in non-contiguous samples. Thus, while crop pollen grains do provide strong evidence of cultivation when present, they rarely fully enclose periods of cultivation (Clement and Horn, 2001; Horn and Kennedy, 2001; Kennedy and Horn, 2008). While our negative finding for pollen of cultivars does not provide any evidence of cultivation, it does not rule out the possibility.
Although there is no evidence within the sediment record of manioc or maize cultivation by indigenous peoples on Barbuda (Jones et al., 2018), pollen signals (Cecropia and Piper) of vegetation disturbance and biomass burning, and pollen of ethnobotanically useful plant taxa (e.g. Solanaceae, Moraceae, Myrtaceae, and all mangrove taxa: Fitzpatrick, 2015; Newsom and Wing, 2004; Siegel et al., 2015) support archaeological evidence (Bain et al., 2017) of pre-Colombian human activities on the island. Taken together, the paleoenvironmental and archaeological data yield a clear record of selective removal of trees for building materials, burning of wood for fuel, and the exploitation of edible wild plants by indigenous peoples.
A sharp decline in local fire activity recorded in the Freshwater Pond macrocharcoal record after ~1250 CE preceded the transition from shallow mangrove wetland to lacustrine environment that occurred around the onset of the LIA (Burn et al., 2016; Figures 2 and 3). We attribute this decline to a sudden decrease in anthropogenic ignitions associated with the abandonment or near abandonment of the island by indigenous peoples, rather than a response to climatic change associated with the onset of the LIA. Supporting evidence from the Indian Town Trail archaeological site suggests that this site was likely abandoned at some point after 1130 CE, (Bain et al., 2017; Harris, 1965). This is supported by an observed decrease in Freshwater Pond’s sedimentary microcharcoal and is consistent with Wilson’s (2007) hypothesis of abandonment in several Leeward Islands by ~1300 CE.
Low concentrations of monotypic macrocharcoal deposited in the centuries following island abandonment suggest that burning of the local landscape was infrequent after human activities declined. The sedimentary charcoal, very low in abundance during that period, may represent the remobilization of secondary charcoal already present within the catchment (Enache and Cumming, 2007) rather than local fire activity. Historical records indicate that the small-scale agriculture introduced by the British in the 17th century was mostly limited to lands adjacent to Codrington Village (Harris, 1965), though cultivation of wetland soils near Freshwater Pond may have continued.
Vegetation history, precipitation variability, and human impacts on Barbuda’s landscapes
Poor pollen preservation within the basal iron-oxide mottled blue-green clays (51–84 cm) of Freshwater Pond indicates that surface sediments oxidized repeatedly as the wetland water table fluctuated during the early Archaic and Ceramic Ages. We interpret these basal clays to reflect fluctuating levels of effective moisture in the northeastern Caribbean prior to the onset of the LIA (Burn et al., 2016). While poor preservation precluded identification of most pollen and spores in the basal sediment intervals, the presence of a few better-preserved grains and spores suggests the establishment of a shallow mangrove wetland that supported an herbaceous cover of ferns, sedges, and aroids ~150 BCE (Figure 3). A mixed evergreen-deciduous xeric woodland of low canopy height (3–10 m; described by Francis et al., 1994 and Lindsay and Horwith, 1997) likely grew near the pond, based on the presence of Fabaceae and Moraceae pollen (probably Ficus citrifolia), with Cecropia pollen indicating disturbance. Pollen of Bursera (likely B. simaruba), a common constituent of the mixed deciduous-evergreen forest that characterizes the Highlands today, is generally poorly represented in the Freshwater Pond sediments, but its presence at ~550 CE (50–60 cm) may reflect that this forest type was well-established at that time (Figure 3). By ~1460–1640 CE, during the early phase of the LIA, Freshwater Pond transitioned from a shallow wetland to a lacustrine environment. Though pollen and spores are better preserved in these upper marl sediments (0–50 cm), the continued prevalence of oxidized pollen grains suggests that the shallow pond dried out intermittently during this period. We interpret a peak in pollen percentages of red mangrove (Rhizophora mangle) and buttonwood (Conocarpus erectus) as evidence of wetland establishment and mangrove expansion in response to increased precipitation (Burn et al., 2016). Despite the variability in effective moisture since that time period, the sediment record indicates that Freshwater Pond has remained a mangrove lagoon to the present.
The probable abandonment of the island after 1250 CE (Bain et al., 2017; Harris, 1965) and the concomitant decreases in anthropogenic fire activity, which occurred sometime before ~1550 CE (the precise timing of which is poorly constrained by the radiocarbon chronology of the sediment record), may have facilitated vegetation succession within the mixed xeric forest communities that were previously disturbed by human activities. The colonizers, Cecropia (Smith et al., 2004) and evergreen shrub, Dodonaea, (likely Dodonaea viscosa, Fern, 2014; Francis et al., 1994), were represented in the pollen record ~1550 CE (1460–1640 CE), along with tropical forest trees Celtis and Vitex, and Eugenia, a common understory constituent that typifies the mixed deciduous-evergreen forests of the Highlands today. Notably, macrocharcoal fragments of Eugenia were identified at all archaeological sites studied by Bain et al. (2017). These taxa, useful for construction, fuel, medicinal purposes and food (see Newsom and Wing, 2004; Siegel et al., 2015; Smith et al., 2004), were extensively harvested by indigenous peoples for fuel and timber, and were later exploited during the Colonial period (Bain et al., 2017; Harris, 1965; Newsom and Wing, 2004). Interestingly, pollen of these tropical dry forest taxa appeared only during post-abandonment succession, possibly reflecting that the greatest forest development on the island occurred at that time. By ~1610 CE (42 cm), we infer an extended period of drought (Figure 3, gray bars) at Freshwater Pond based on declines in pollen of mangroves and moist habitat indicators (aroids, fern spores), and increases in pollen of taxa characteristic of open environments (e.g. Scrophulariaceae and Asteraceae), fungal spores, and damaged pollen grains and spores. This extended dry period is corroborated by concomitant decreases in the abundance of fossil shells of Pyrgophorus parvulus (a freshwater gastropod) and positive excursions of stable oxygen isotope data recovered from ostracod valves of the same sediment core (Burn et al., 2016).
Mangrove forest resources adjacent to Freshwater Pond may have been exploited by settlers during the Colonial Period. While historical sources indicate that Barbuda was not permanently settled by the English until the 1660s, it was occupied intermittently by British settlers. Archival sources mention the occupation of the southern part of the island before the arrival of hundreds of enslaved people as the Codrington family of Antigua took over management of the island in 1681. Following that event, the island supplied Antigua with livestock, charcoal, timber, and subsistence crops for almost 200 years (Bain et al., 2017). Pollen, not identifiable to genus/species, of the yam family (Dioscoreaceae) in sediments dating to roughly 1610 and 1660 CE and pollen of the squash family (Cucurbitaceae) from ~1710 to 1780 CE may reflect cultivation or may represent wild relatives in these families. The peak in Piper pollen dating to ~1660 CE, coinciding with the permanent settlement of the island, signals forest disturbance that may have been related to subsistence agriculture by the few early colonists and/or timber extraction for construction. Freshwater Pond may have provided a rare source of drinking water for the colonists along with its adjacent arable soils. During the following two centuries, an extensive system of wells and animal pens were established across the island in order to ensure a year-round supply of fresh water (Boger et al., 2014; Boger and Perdikaris, 2023). The surrounding mangrove and highland forests would have provided hardwood trees for construction and export.
Increased pollen of Fabaceae (legumes) and Piper along with declines in Moraceae pollen (probably Ficus citrifolia) during a second period of extended drought ~1710 CE (30 cm) (Burn et al., 2016) may reflect short-term expansion of the xeric woodland surrounding Freshwater Pond at the expense of the mixed evergreen forest of the Highlands in response to drought conditions. Concomitant increases in the concentrations of fungal spores, which we found to be excellent predictors of changes in effective moisture at Freshwater Pond (Burn et al., 2016), may indicate greater use of the site by open-range grazers (European cattle or goats) during the drought or may be associated with exposed and decaying aquatic vegetation.
Conclusions
The Freshwater Pond sediment record documents vegetation shifts, fires, and land-use activities on the island of Barbuda across more than two millennia that examines, supports, and expands interpretations from other records of climate and vegetation change (Burn et al., 2016; Jones et al., 2018) and human history (Bain et al., 2017; Faucher, 2018) on the island. The paleoecological record presented here documents the presence of ethnobotanically useful and disturbance taxa contemporaneous with burning by indigenous inhabitants of the island during the Ceramic Age (500 BCE–1500 CE), with sedimentary charcoal recording possibly earlier burning. Past excavations of Archaic Age (3000–500 BCE) sites document the seasonal exploitation of the island for conch fishing (Rousseau et al., 2017; Watters et al., 1992), a finding supported by relatively high sedimentary charcoal from a period below our earliest calibrated radiocarbon date in the range of 42–354 BCE (2σ; Table 1).
Plants used for food, medicine, and building materials, and disturbance indicators are represented in the Freshwater Pond pollen record including several mangrove genera, Cecropia, Celtis, Dodonaea, Eugenia, Piper, Vitex, Asteraceae, Cucurbitaceae, Dioscoreaceae, Euphorbiaceae, and Solananceae. Our pollen record supports archaeological and historical records of vegetation clearance for fuel and construction and medicinal purposes and adds possible evidence (the presence of Solanaceae pollen grains) that some areas of the island, probably near the pond, were exploited by indigenous groups for small-scale plant cultivation (gardens). To date, no archaeological or paleoecological studies from the island have revealed evidence of large-scale agriculture by indigenous peoples (Bain et al., 2017; Jones et al., 2018). However, our charcoal record indicates that early occupants of Barbuda regularly burned vegetation near Freshwater Pond until ~1250 CE, after which archaeological evidence may suggest temporary abandonment (Bain et al., 2017), an interpretation supported by a dramatic decline in sedimentary charcoal. During this period, our sediment record indicates the transition of Freshwater Pond from a wetland to a permanent waterbody, with evidence of vegetation regrowth as sedimentary pollen of rare tropical dry forest types (i.e. Celtis, Vitex, Eugenia) that were only present in sediments dating to ~1550 CE (1460–1640 CE; 2σ error ranges), a period in which the island’s forests may have been the most abundant and diverse of the past 2000+ years. The much-reduced number of macrocharcoal fragments deposited in Freshwater Pond during the Colonial and Post-Colonial Ages may reflect redeposition of charcoal rather than burning within the watershed. Microcharcoal production remained consistent during this period, perhaps reflecting in part activities at the lime kilns noted during archaeological surveys near the western and southern coasts.
Our interpretation of sediment record as low fire activity combined with a high relative degree of vegetation disturbance by ~1660 CE reflected changes in land use patterns associated with the permanent settlement of the island. The pre-Columbian slash and burn agriculture of the Ceramic Age was replaced by the selective cutting and removal of valuable timber species growing in forests of the Highlands and elsewhere, and livestock rearing (Bain et al., 2017). Wetlands near the pond may have been exploited for subsistence agriculture as suggested by the presence of gourd and yam pollen grains, but those grains might also represent wild relatives. While subsequent human populations altered the island landscape for more than 2000 years, the Little Ice Age (1400–1850 CE) brought repeated droughts to the region (Berland et al., 2013; Burn et al., 2016) that would have further reduced fuel conditions and vegetation succession against a legacy of vegetation burning, timber removal, and livestock grazing. The paleoecological record here adds new evidence and support to prior studies of Barbuda’s dynamic landscape history.
Footnotes
Acknowledgements
We are immensely grateful to Dr. Reg Murphy, and members of the Barbuda Council for their support of this research.We appreciate laboratory assistance from Virginia Tech undergraduate researchers, MaryAnna Garifo and Rylan Liddy.
Author contributions
All authors contributed to the interpretation of results, fieldwork, and writing of the final paper.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project is based upon work supported by the National Science Foundation, Grant No. 0851727 to S. Perdikaris, and funds from the Social Science and Humanities Research Council of Canada via a Research Development Initiatives Grant to Bain (PI), Kennedy, and Burn (820-2010-0189), and doctoral funding to A. LeBlanc. Additional funds from the Association of American Geographers Paleoenvironmental Change Specialty Group to A. LeBlanc supported this research.