
Abstract
This study presents the results of a systematic survey of La Pastosa Islet, located 700 m off the west coast of Río Negro, Argentina. This small islet has the particularity of hosting the northernmost colony of Magellanic penguins (Spheniscus magellanicus) to date and contains evidence of its use by hunter-gatherer groups during the late Holocene. This growing colony started around 2002 and has had a significant impact on the subsurface archeological record of the islet by increasing erosion and exposing buried materials through nest excavation, particularly lithic artifacts. The spatial analysis of the distribution of artifact and penguin burrows indicates that the actions of the latter generated a “snapshot” of the subsurface distribution of the lithic record. The observed distribution suggested higher discard rates at the center of the islet. Furthermore, this study shows the synergy between both types of records in the formation dynamics of the lithic landscape and the agency of penguins as active landscape engineers. Finally, the obtained results have important implications for the management of the protected sector of the recently established Islote Lobos National Park, within which the study area is located.
Introduction
Studying the presence of penguin colonies in coastal areas with evidence of past human occupation is crucial to better understanding the complex relationship between penguins and their environment. By fully appreciating the importance of their presence in shaping coastal landscapes and conserving archeological heritage, a deeper understanding can be gained. Although the literature on this subject is scarce, there has been interest in economic, taphonomic, and landscape modification aspects. The economic aspect involves studying the potential utilization and exploitation of these species by human groups in the coastal regions of Patagonia (Cruz et al., 2014). Taphonomic interest focuses on how this species incorporates into archeological sites and contributes to archeological deposits (Borella and Cruz, 2012; Cruz, 2017; Massigoge et al., 2015). However, our interest in this species lies in its taphonomic role, as it significantly modifies the landscapes in which it settles. These alterations can include vegetation removal, accumulation of feathers and guano, soil compaction, and creation of trails or paths (Butler, 1995). Several studies have addressed the effects of this species on the coastal areas of the Argentinian Patagonian Atlantic coast (Cardillo et al., 2015; Cruz, 2004, 2007; Ercolano et al., 2016; Hammond et al., 2013; Pearson et al., 2010). The particularity of the case study presented here is that we will examine how the presence of an extensive colony of Magellanic penguins (Spheniscus magellanicus) can influence the lithic record in a delimited and restricted space currently subject to intense faunal disturbance. Through the use of indicators of spatial and morphological transformations of the lithic record and the intersection of spatial variables, the main purpose of this study is to characterize the spatial distribution of penguin nests and nest remnants, along with lithic scatters, to evaluate their association using statistical methods. Furthermore, through taphonomic and technological analyses, we aimed to characterize the formational history of lithic assemblages and strategies of islet use by hunter-gatherers. In addition, this area is part of a National Park and is subject to protection and management regulations. Therefore, we hope that the information generated here will be useful for decision-making in the management of places of high biological and cultural sensitivity.
Background
The San Matías Gulf (SMG) extends from Punta Bermeja in the north (Río Negro Province) to Punta Norte in the south, in the Valdés Peninsula (Chubut Province) (Figure 1). The coastline of Río Negro extends for approximately 380 km. The region is characterized by a semi-arid temperate climate, with an average temperature of 12°C and annual precipitation ranging between 100 and 350 mm (Labraga and Villalba, 2009). It has a semi-diurnal tidal regime with amplitudes between 3.5 and 7.6 m (Naval Hydrography Service, 2009). The prevailing winds from the west (NW, W, and SW) reach maximum speeds of ca. 90–100 km/h, with the strongest winds occurring between September and January (National Meteorological Service, 2018). The western sector of the SMG coast runs in a north-south direction and extends from the city of Las Grutas to Puerto Lobos, on the border with Chubut Province.
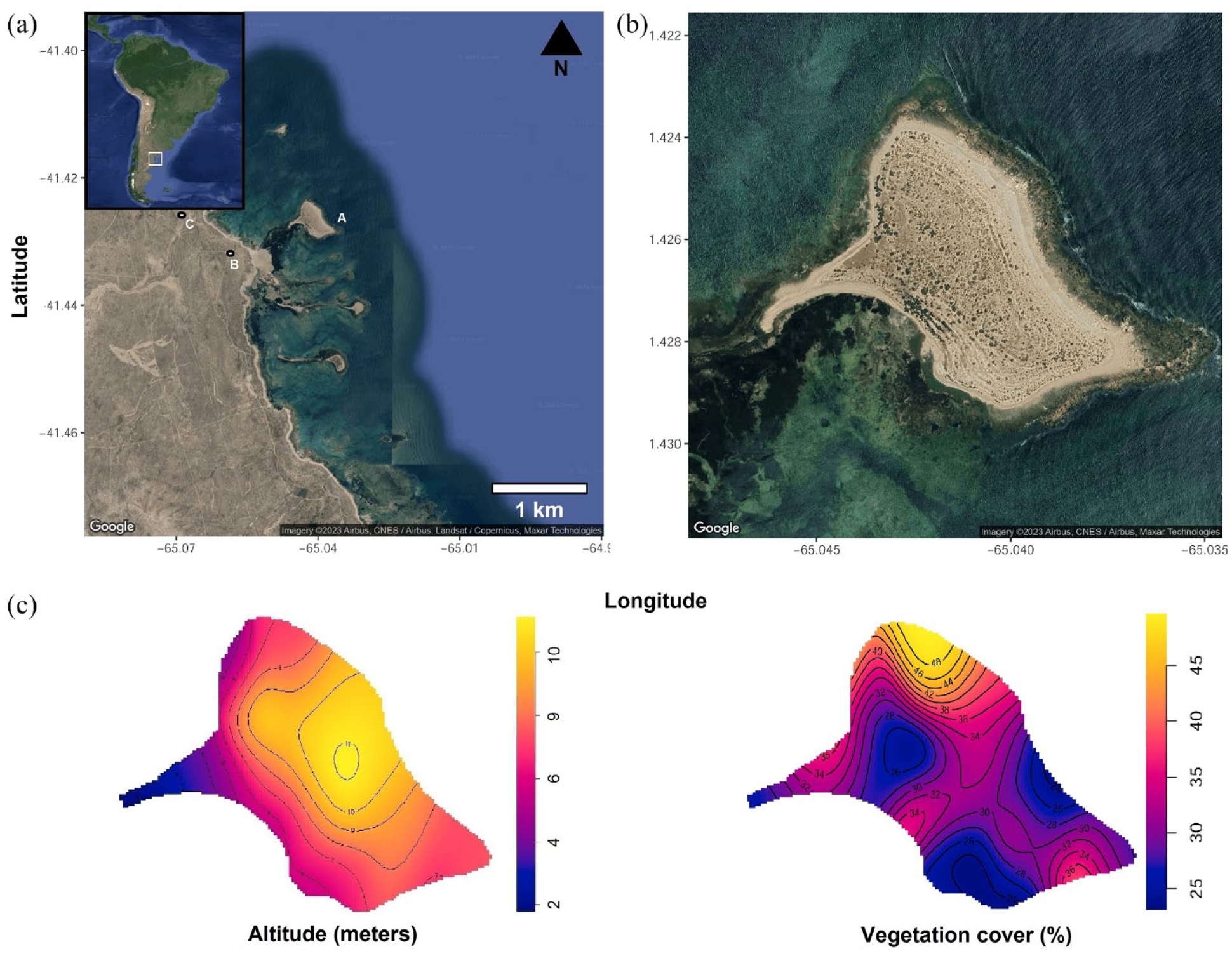
1. Map of Islote Lobos complex: (a) La Pastosa Islet (left), (b) Parador Q-B site, and (c) Cañadón del Puma site. 2. Detailed view of La Pastosa Islet. 3. Topography (left), and vegetation cover (right).
The Islote Lobos National Park is composed of several loci along the coastal strip and on the La Pastosa islet, connected to the coast only at low tide (Figure 1) (Borella et al., 2020). Currently, this area is subject to environmental protection regulations owing to the high biodiversity of marine and terrestrial species. La Pastosa islet (LP) has the largest surface area (approximately 18 Ha) among those that make up the complex. From north to south, the islets are named Lobos, Ortiz Norte, Ortiz Sur, Redondo, and Los Pájaros (Sánchez, 1973). They are mainly composed of granite outcrops, which are an extension of the Sierra Grande Plutonic Complex that reaches the shoreline. These granite outcrops are interspersed with solid limestone boulders, providing a foundation for semi-stable dunes adorned with semi-shrub vegetation such as Jarilla (Larrea divaricata) and Jume (Suaeda divaricata) present in patches of medium-sized shrub between 1 and 3 m tall, adapted to coastal and saline conditions and offer shelter for penguins (Cardillo et al., 2015; Sánchez, 1973; Figure 2). La Pastosa is located 768 m from the mainland and is connected through a tombolo during low-tide periods (Kokot and Pisani, 2011). The intertidal zone is formed by contact between the two outcrops, granite and limestone, partially covering it and forming tidal pools of varying depth and variability, where a marsh composed of Spartina densiflora and Salicronia ambigua develops (Cardillo et al., 2015; Sánchez, 1973). Conversely, the opposite process occurs along the coastal perimeter of the islet, where the accretion of sandy sediments, pebbles of various sizes, and accumulation of pinniped bones from the reproductive sea lion (Otaria flavescens) colony that exists there occur (Borella and Borrero, 2010, 2019; Borella and L’Heureux, 2019; Cardillo et al., 2015). From an environmental perspective, this area hosts a high biodiversity of birds and is home to one of the northernmost penguin colonies and a significant colony of fur seals. This fur seal settlement extends approximately 5 m from the rocky coast toward the interior of the islet and consists of about 722 individuals (data from the 2016 census; Albornoz and Núñez, 2016). Given its limited extent on the islet and the naturalistic observations made, we believe that the fur seals are not significant agents in the faunalturbation (Wood and Johnson, 1978). However, we do not rule out their possible influence. Islote Lobos National Park is also a nesting and resting site for numerous species of coastal and marine birds, including various migratory birds (Canevari et al., 1998). In the nesting area, birds such as the royal tern (Sterna maxima), kelp gull (Larus dominicanus), Neotropic cormorant (Phalacrocorax brasilianus), various types of herons like the black-crowned night heron (Nycticorax nycticorax), American oystercatcher (Haematopus palliatus), spectacled duck (Lophonetta specularioides), among many others, can be found. As for terrestrial species, only the small South American gray fox (Pseudalopex griseus) has been observed on the La Pastosa islet. This area represents a biologically important marine reservoir on the northern coast of Argentine Patagonia as well as a testament to the continuity of human use of this marine space over time. The islet La Pastosa, along with other islets, is considered of intangible nature for most of its extension, and access is restricted. Therefore, in the year 2022, it is created the Islote Lobos National Park, and through archeological works in the area, participation took place in the management plan, focusing on developing conservation strategies and recommendations for effective and efficient treatment of the cultural landscape and the heritage in this coastal sector.

Magellanic penguin (Spheniscus magellanicus) in the islet La Pastosa. (a) Penguins on the rocky coast of the islet, where they go for feeding. (b–d) Different kinds of nests were observed in the islet. Note the impact of nesting on the sediment and vegetation.
From an archeological point of view, the area has relatively low visibility given the preservation conditions. The low density and diversity of recovered materials suggest the sporadic and relatively punctual use of this space (Borella et al., 2020; Cardillo et al., 2015). Accumulations of mollusks, bone remains, charcoal, and lithic material appear discreetly buried in eolian mantles with little development and semi-vegetation. Moreover, along the continental coastal strip, excavations and surface archeological research have been conducted at sites with high-artifact density (Borella et al., 2020). The recovery of bioarcheological remains on the La Pastosa islet and the continental sector allowed the dating and study of paleodiets and delimited the temporal range of occupations in this coastal sector (Table 1) (Borella et al., 2016, 2020; Favier Dubois et al., 2009). The chronological range obtained for the islet is showing at least two chronologically different moments of occupation or use of space in the Late Holocene, one at ca. 3700 and another ca. 2700 BP. The remains were recovered in aeolian contexts, with evidence of recent exposure.
Radiocarbon chronology obtained from the study area.

The dates were calibrated using OxCal v4.3.2, considering a local reservoir effect (266 + 51) at two standard deviations with the SH Cal 13 curve (Borella et al., 2020; Favier Dubois, 2009; Favier Dubois and Jull, 2018; Stuiver et al., 1986).
In the study area, the archeological site Parador QB is located in front of the islet, a few meters from the current coastline, and approximately 800 m away from it (Borella et al., 2020). Surface collections and systematic excavations were carried out at this site; establishing a temporal range between 1533 and 656 calibrated years BP (see Table 1). The faunal evidence for Islote Lobos area indicates the exploitation of a wide diversity of marine resources, such as otariids (Borella and L’Heureux, 2019), fish, mollusks, and terrestrial resources, such as guanaco (Lama guanicoe) (Borella et al., 2020). The lithic materials recovered on the surface and during excavation indicate the use of local rocks (such as chalcedony, silex, and chert) and non-local high-quality obsidian, which come from distances greater than 300 km from the study area (Alberti, 2016). Among the recovered tools, those related to resource processing, such as scrapers and grinding stones, stand out. Although no chronological overlap has been detected, until now, between the occupations of the islet and the immediate coastal area (see Table 1), this site is of comparative relevance because of its proximity and spatial relationship with the surrounding environment and its resources; therefore, we will use it as a reference for the interpretation of the results.
In La Pastosa islet, transects were previously carried out, suggesting that the lithic record tends to accumulate in the highest sector of the islet (Cardillo et al., 2015). In most cases, their exposure seems to be directly associated with faunal disturbances related to the recent installation and increasing extension of penguin colonies. The results indicate that the record is preferentially presented as small clusters. Although the sample size was small, a non-random distribution of materials in space was observed. These results suggest that unequal distribution could be linked to hunter-gatherers’ persistent use of specific sectors of the islet to the detriment of others. Furthermore, evidence suggests that the action of penguins plays a fundamental role in geomorphological dynamics, vegetation cover, and in shaping the archeological record.
Faunalturbation by penguins
From a taphonomic point of view, animals are agents capable of generating both morphological and distributional effects as a result of two main types of activity on the substrate: transit and excavation (Eren et al., 2010; Gifford-Gonzalez et al., 1985; Lopinot and Ray, 2007; Schoville, 2019; Weitzel et al., 2014; Wood and Johnson, 1978). These fall under the category of distributional modifications, which are processes that affect the horizontal and vertical distributions of artifacts (Nielsen, 1991). The former can occur through trampling of artifacts on the surface, and its potential for modification depends on its size. The distance traveled depends on both the size of the acting agent and artifacts. It is possible that the agent’s range of action involves only changes in the position and orientation of the major axis without displacement (Schoville, 2019). Penguins are one of the largest and most numerous of the burrow-nesting colonial seabirds (Butler, 1995). As a consequence, taphonomic effects resulting from trampling by animals, can include horizontal and vertical displacements, inversions, burial as a result of vertical movements, exposure of artifact pieces due to the movement of the matrix that contains the record, and denudation, which involves the removal of vegetation cover that facilitates sediment erosion, favoring the rapid action of other agents (wind, water, animals) (Cardillo et al., 2015; Cruz, 2004, 2007; Schoville, 2019).
The Magellanic penguin (Figure 2) is a flightless bird of 35 and 45 cm in height and approximately 3 kg in weight that breeds in a geographic range spanning from approximately 55°S to 40°S, including islands and coasts of South America, such as the Falkland/Malvinas Islands. In the Atlantic region of South America, their breeding range extends from Cape Horn to the Islote Lobos Complex (41°33′S), with the latter being the northernmost colony on the Atlantic coast and therefore the northern distribution limit for breeding. Magellanic penguins are colonial breeders, which means that they typically breed and nest in large groups or colonies. They are commonly monogamous, and pairs meet repeatedly in the same colony to breed (Boersma et al., 2013; García Borboroglu and Boersma, 2015). Once they reach maturity at around 2–4 years of age, they return to their natal colony to find a mate. In the Atlantic Ocean, colonies show mixed growth trends. Two of the largest colonies, Punta Tombo and Isla Leones in southern Chubut, have experienced a decline, whereas settlements in Península Valdés are increasing. Additionally, new colonies have emerged, such as those in the La Pastosa and Redondo islets, which expanded their distribution range northward. The breeding of Magellanic penguins in La Pastosa was first recorded in 2002, with 22 pairs on the Redondo islet, which grew to 4748 pairs in La Pastosa, Redondo, and Pájaros in 2011, with a population growth rate of 1.91% (Pozzi et al., 2015; Schiavini et al., 2005). Therefore, since 2002, the estimated time of settlement, growth, and impact of the colony in the area of La Pastosa islet has been configured. Magellanic penguins build nests using rocks, twigs, and feathers on the surface, under the shelter of available vegetation, or digging burrows below the surface (see Figure 2). The recorded dimensions of the burrows and nets can reach 1 m in length and up to 1–2 m in depth, removing substrate sediments of at least 0.05 m3 (Stokes and Boersma, 1991) (Figure 2). The repeated use of these nests and space triggers erosive processes in the terrain, removing vegetation and displacing redeposited material and sediments (Cardillo et al., 2015). The impact of penguins as disturbing agents on the archeological record along the Argentinian Patagonian coast has been recognized (Cruz, 1999, 2004, 2007; Zubimendi et al., 2016); therefore, their impact on this sector is not minor and requires a detailed analysis. This is how the lithic materials currently observed on the surface are redeposited and have a high displacement probability, depending on their location (Cardillo et al., 2015).
Methods
Fieldwork
In situ records without collection were carried out using oriented transects to assess distributional information for the entire islet, aiming to integrate and discuss the significance of this sector in the study of archeological record evolution and its relevance within the management plan of the area. The distribution of penguin nests and lithic assemblages was mapped, including taphonomic and technological variables such as artifact type and raw material. This approach aimed to minimize the impact on the archeological record, allow longitudinal tracking of surface findings, and adhere to the management plan of the protected area (see Background). Thirteen double transects (26 segments in total), spaced 50 m apart and oriented north–south, were conducted by two researchers separated by 5 m (Belardi, 1992; Borrero et al., 1992; Dunnell, 1992; Ebert, 1992; Foley, 1981; for the San Matías gulf see Cardillo, 2009; Manzi et al., 2011). The starting and ending points of transects were the current coastlines. Owing to the irregular shape of the islet, transects varied in length, ranging from 40 to 528 m, with an average of 292 m. Considering that each individual covered 5 m in total, approximately 76000 m2 was covered, which corresponds to approximately 42% of the estimated area of the entire islet (183000 m2). Each finding was georeferenced and assigned the same GPS coordinates for the specimens found in close proximity (penguins, nest or artifacts; within a radius of approximately 1 m). Non-collected findings were recorded on predesigned sheets for subsequent quantification and monitoring.
Lithic materials were analyzed following Aschero’s (1975, 1983) lithic typology, considering broader techno-morphological types, such as flakes and debris (débitage), cores, and tools. Taphonomic analysis includes the assessment of morphological and distributional taphonomic effects. Morphological taphonomic effects refer to the post-depositional modifications suffered by artifacts that imply changes in their formal attributes (Borrazzo, 2016). Wind abrasion (corrasion) (Breed et al., 1997) and its intensity (Borrazzo, 2006) were recorded in four stages (W0, W1, W2, and W3) and carbonation (coating or rock coating). This deposit of accretionary minerals mainly carbonates, of varying thicknesses, alters the appearance of the artifact surface (Dorn, 2009). The area covered by rock coating was recorded at the following levels: slight 1, moderate 2, and intense 3. Spatial taphonomic effects or transformations are changes in the artifact distribution, such as its displacement (i.e. vertical and horizontal). These displacements were registered in previous work (Cardillo et al., 2015) and it is currently being evaluated too. The formulation and application of a taphonomic analysis on surface lithic materials, prior to the technological analysis, are conducted to explain and understand the formation processes that affected the assemblages after their deposition (Borrazzo, 2016). This enables the construction of a formational history of the assemblages and a characterization in terms of their resolution and integrity (Binford, 1981; Schiffer, 1987).
In order to display the relationships and interactions between various taphonomic variables, lithics raw material and artifacts categories we use the alluvial plot (Figure 4). Alluvial plots allow for the comparison of multiple taphonomic agents and their effects on lithic assemblages, aiding in the identification of dominant processes and their impacts on the archeological record.
Based on the calibrated dates (see Table 1), the rate of deposition of lithic artifacts in La Pastosa was estimated using two different approaches, taking into account the minimum and maximum temporal distance within the dating ranges. Additionally, during the survey, the altitude (measured with GPS), landform, vegetation coverage (%), visibility (poor, fair, well), slope (%), artifact condition (complete, fragmented), and raw material were documented.
Spatial analysis
For the spatial study of the recorded variables, we followed the methodology of point pattern analysis (Baddeley and Turner, 2006). The point pattern approach in spatial analysis is employed to examine the organization and distribution of points within a spatial context. This entails the analysis of spatial patterns and interactions among individual point locations, aiming to gain insights into the underlying processes and structures. A commonly used model to study point patterns, the Poisson process, is a stochastic model used to describe the occurrence of events in time or space. In the case of spatial Poisson processes, events are modeled as points distributed in a continuous or discrete space. The Poisson process is characterized by the fact that the number of events occurring in the non-overlapping regions of space is independent and follows a Poisson distribution.
In a homogeneous spatial Poisson process, the intensity or rate of an event occurrence is constant throughout the entire spatial domain. This implies that the probability of an event occurring at any point in space is the same. In other words, event occurrence is uniform and does not vary with location. Mathematically, the intensity function of a homogeneous spatial Poisson process is constant and denoted by λ, where λ > 0. The intensity function represents the average number of events occurring per unit area or volume in space (Baddeley and Turner, 2006; Diggle and Cox, 1983).
To measure the degree of clustering or dispersion of points in a spatial domain, we used Ripley’s K function. Ripley’s K-function is defined as the expected number of points within a certain distance, r, of each point in the point pattern, normalized by the density of points in the study area. The interpretation of Ripley’s K-function depends on its value in relation to a reference value; in this case, a null model was generated via random simulation. If K(r) >0, this indicates clustering or spatial aggregation of points, meaning that points tend to be closer to each other than expected under a random distribution. If K(r) <0, this indicates dispersion or spatial regularity of points, meaning that points tend to be farther apart from each other than expected under a random distribution. K(r) =0 indicates a random spatial pattern, where points are randomly distributed with no clustering or dispersion. The results are represented by a K-function plot, where the calculated K(r) values for the empirical distribution are plotted against the distance r, along with the random simulation. The significance of the results was achieved by resampling the data 100 times.
We also wanted to explore the extent to which both spatial processes, the penguin nest, and lithic artifacts are correlated in space. Our expectation is that if the distribution of lithic artifacts on the surface is a function of nest density, their frequency will increase in sectors where more nests are recorded. On the other hand, if there were differential artifact discard behavior within the islet by human groups, there would be no relationship between the two. To assess the degree of separation of different spatial processes within the islet, we used a spatial segregation test (Diggle et al., 2005; Kelsall and Diggle, 1995). This method estimates the intensity function using the kernel smoothing technique for each variable separately, which represents the spatial density of each variable based on the observed data. The segregation index measures the dissimilarity between the observed cross-type intensity function and the expected intensity function under complete spatial randomness. The complete spatial randomness intensity function represents the expected spatial distribution of events in the absence of spatial segregation. The confidence intervals and statistical significance of the results were assessed by randomly permuting the empirical distributions 100 times (see Supplemental Information). Prior to spatial analysis, geospatial coordinates with the same values were jittered to avoid the existence of an overlap between them, as some points had the same GPS value because of their spatial proximity.
For the analyses, the R program was used (R Core Team, 2023) and the packages ggmap (Kahle and Wickham, 2013), geoR (Ribeiro et al., 2020), maptools (Bivand and Lewin-Koh, 2021) were used mainly for the visualization and management of maps. Statistical analysis was performed using spatstat and sp packages (Baddeley et al., 2013; Pebesma and Bivand, 2005); see Supplemental Materials.
Results
Lithic analysis
A total of 81 artifacts were recorded in transects, of which 70% were complete and 28% were fragmented. Among these, 90% were débitage, 0.02% were tools, and 0.07% were cores. This contrasts with the surface material recovered at the nearest site, Parador QB, with a sample of 37 artifacts, which had 46% intact and 54% fragmented, with 83% being débitage, 0.02% tools, and 0.06% cores (Borella et al., 2020). In the islet, artifacts without cortex represented 94% of the sample, whereas only 6.17% had cortex on their dorsal face, while in the continental site, this represented 65% and 35%, respectively. The most frequent raw material in the islet is biogenic chert (90%), followed by chalcedony (0.8%) and silex (0.1%). The surface record of Parador QB is composed of 45% local biogenic chert, 40% chalcedony, 8% silex, and 5% obsidian from external sources (Alberti, 2016; Borella et al., 2020). This suggests that mainland sites have a more diverse use of rocks and relatively high proportions of high-quality chalcedonies that were not immediately available in the area, whereas on the islet, the diversity was lower and readily available chert from the rocky shore was preferred. Both sites have similar artifact components and environmental conditions that expose the lithic material (Carranza, 2022) and some level of planning for fishing activities is evident (Borella et al., 2020; Cardillo and Scartascini, 2016). Parador-QB is located immediately behind the line of semi-stabilized sand dunes in front of La Pastosa islet on the continental coastal fringe, there wouldn’t be intense exposure to the wind with a sedimentary load. Thus, the presence of vegetation could be acting as protection for the artifacts on the surface. While in La Pastosa the evidence obtained suggests that the lithic record is continuously forming, in this case, due to the action of penguins’ work. The main morphological effect recorded was carbonation (coating or rock coating) for 40% of the sample (Figure 3). When it occurs, it is mostly at level 1 of less intensity (63%), then in equal percentages of levels 2 and 3 (19%). Second, corrasion (wind abrasion) occurred in 2.46% of the sample, with a tendency toward the W1 stage. This is a very low frequency compared to that of the continental Parador QB site, where corrasion is the main morphological alteration above carbonation, and 32% of the samples exhibited this alteration (see Table 2 Supplemental Material). Also, it occurs in low-intensity stage W1 (54%). Finally, other characteristics, such as excretion and thermal disturbance, were recorded (Figure 3). While the former occurred in 14% of the sample, the latter was recorded in only 5% of the artifacts.

Artifacts recorded in the Pastosa islet transect: (a) a complete flake of chert without morphological alterations, (b) complete chert flake with penguin excrement on its surface, (c) chert flake with thermal alteration, (d) possible fragmented chalcedony tool with carbonation, and (e) chert flake with carbonates.
As seen in the alluvial plot (Figure 4, see also Table 2 Supplemental Material), the recorded variables are proportionally represented in the different raw materials. However, greater variability is observed in biogenic chert, which is the most abundant rock in La Pastosa, particularly in low-proportion registered modifications, such as thermal alteration or wind abrasion. The common pattern observed (36% of the sample) is that artifacts are mainly represented as complete debris of chert, without carbonation or other surface modifications (Figure 4). The less frequent pathway (12% of the sample) is represented by those chert artifacts that are broken or unbroken with carbonation. Thermal alteration, wind abrasion, or penguin excrements on the surface are observed, along with carbonation, in relative frequencies below 5% of the entire sample.
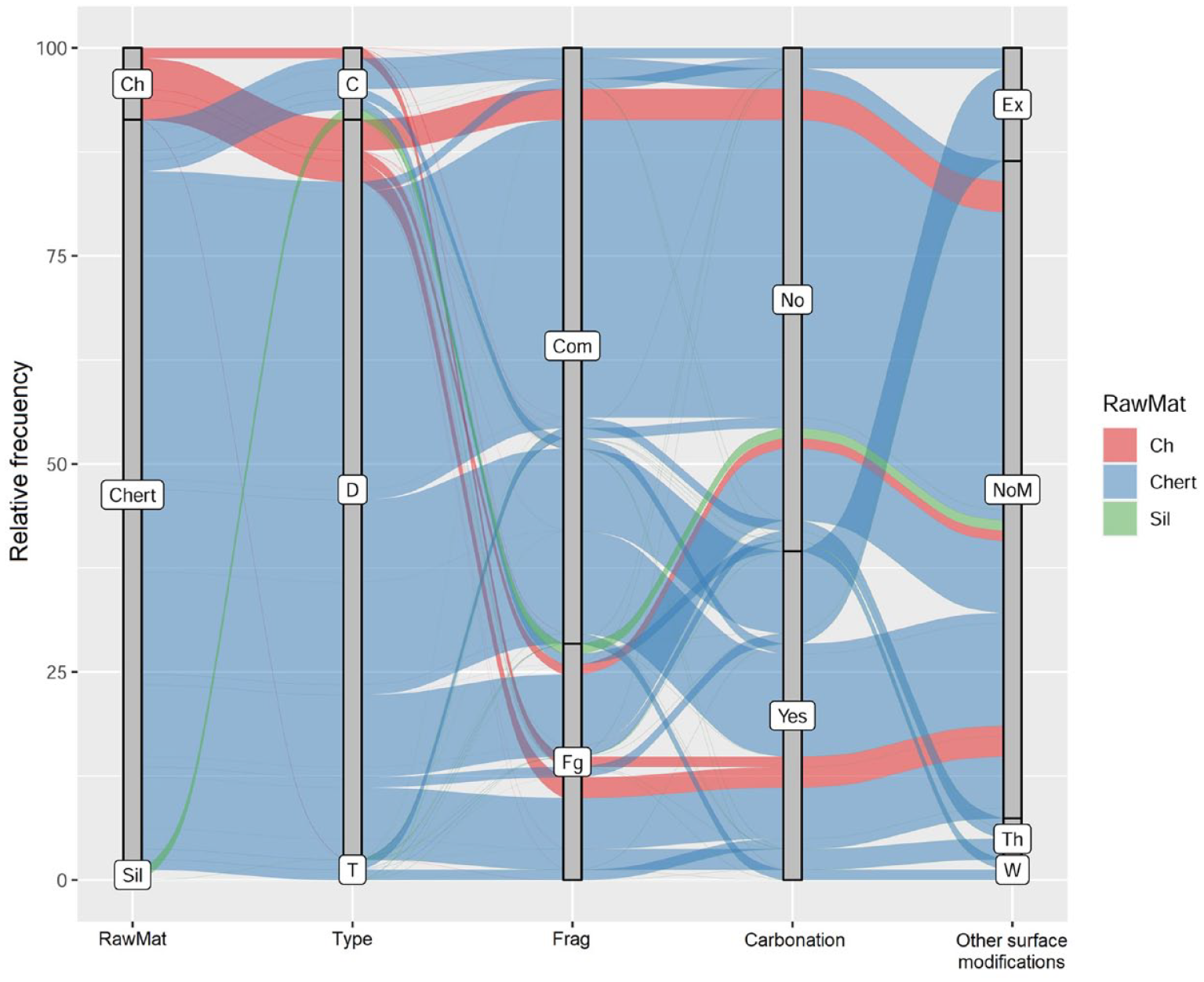
Alluvial plot of percentage distribution of main characteristics recorded on lithics. Ch: chalcedony; Sil: silex; D: debitage; T: tools; C: cores; Com: complete; Fg: fragmented; Th: thermal alteration; W: wind abrasion; NoM: No other surface modifications; Ex: penguin excrement.
Taking into account the minimum temporal distance of 557 years between calibrated dates, the deposition rate was approximately 0.14 artifacts per year. On the other hand, when considering the maximum temporal distance of 968 years between dates, the deposition rate was 0.08 artifacts per year. Finally, considering that approximately 76000 square meters were covered, the artifact density on the islet is about 0.01 artifacts per square meter.
Spatial patterns
Measuring the impact of penguin colonies on the environment is challenging owing to the lack of historical data on the topography and vegetation coverage of the islet. Nevertheless, it is evident that penguins have become significant agents in shaping landscapes over the course of decades. Transit spaces (Figure 5a) within the islet are predominantly devoid of vegetation and surrounding shrub patches where various types of organic and inorganic waste accumulate (Figure 5b). Underground nests (Figure 5c) were found in both vegetation patches and transit spaces. These three contexts are part of the same taphonomic process, where erosion in transit spaces and nest excavation expose materials that are then transported along transit paths (Figure 5b), redeposited in vegetation patches, or trapped in collapsed nests (Figure 5c).

In the main left image are marked the different penguin-made contexts for lithic findings observed during the survey: (a) “marginal” zones: with low circulation were artifacts and different natural debris patches tend to accumulate, (b) transit zones: where the frequent transit occurs along the artifacts, and (c) active and collapsed nests: where lithic material is unearthed, but which also act as taphonomic traps. The arrows indicate the position of the artifacts.
Regarding slope, no significant changes were observed along the routes. In general terms, 81% of the slope is gentle, and in 18.27% of the cases, it is flat, while only 0.8% represents a moderate slope. The predominant vegetation cover was 25%, accounting for 59.01% of the vegetation cover, while areas with higher vegetation cover ranged from 31% to 0.20% with greater density (Figure 1).
The density and quadrat plots (Figure 6) show a higher density in the center of the islet for lithic scatters that are not spatially related to penguin nests. The distribution of nests, on the other hand, although it shows a greater concentration toward the center of the islet, is also distributed toward the south and southwest. Likewise, in almost all the sampled sectors, there was a positive record of nests, with the exception of quadrats that corresponded to beach sectors at the borders of the islet or with more restricted access to the ocean, which was observed at the northern border.
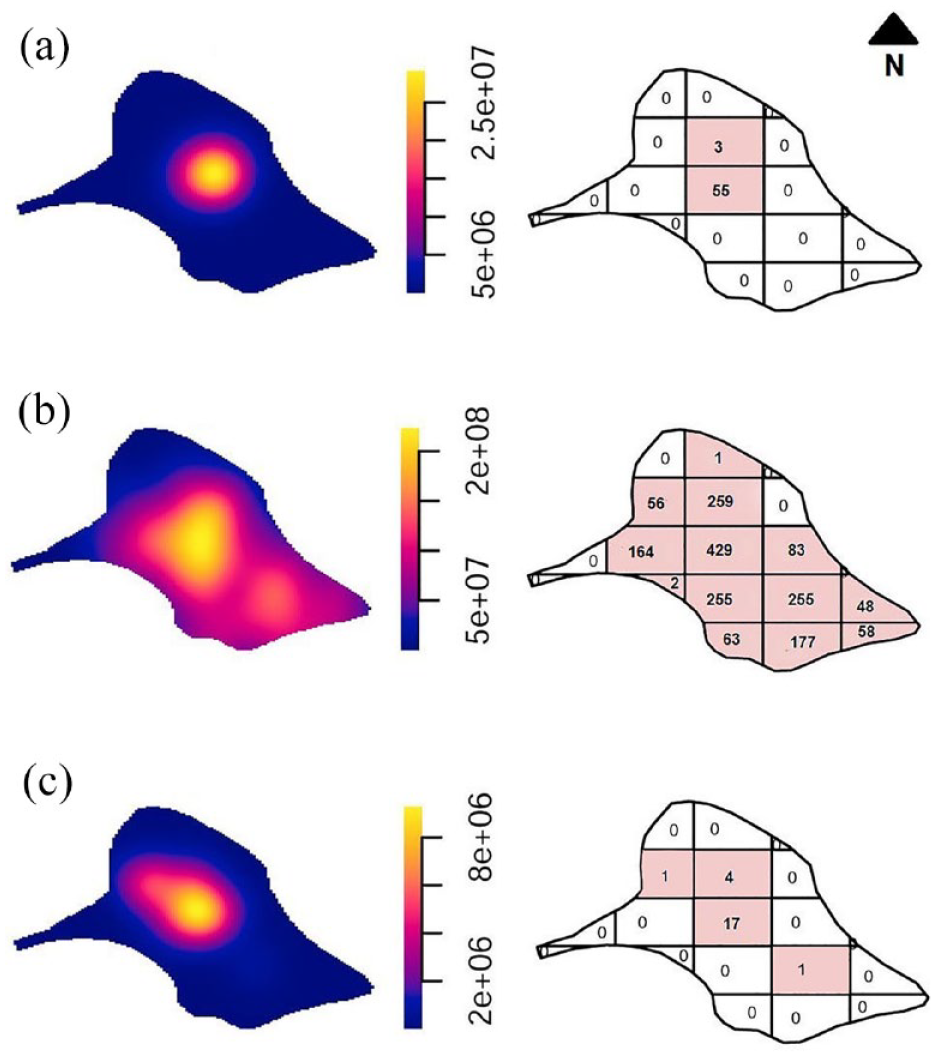
Density and quadrat count for: (a) only artifact scatters, (b) only nests, and (c) artifact scatters and nests. Positive counts are depicted in colored quadrats. Each complete quadrant has 145 × 130 m.
The third distribution (Figure 6c) shows the co-occurrence of lithic scatters and penguin nests, with a higher intensity toward the center of the islet, and low-frequency finds toward the south and west. This indicates a partial overlap between the two distributions but with a greater intensity or concentration for the lithic record.
Also, the K test over the adjusted model of complete spatial randomness suggests significant clustering for the distribution of nests and artifacts (Monte Carlo test: 2/100 = 0.02; Figure 7).
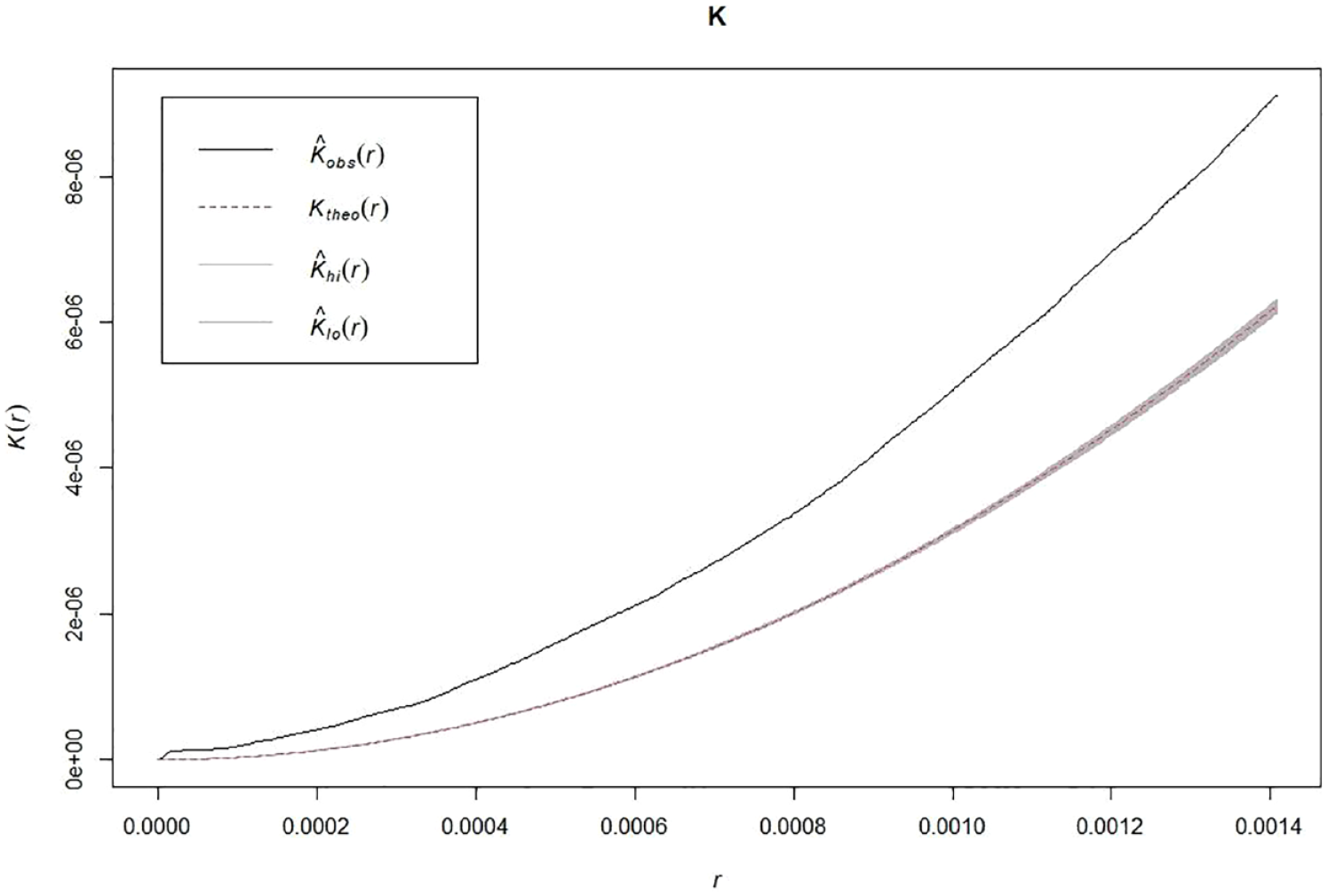
Ripley’s k values for the distribution of nests and lithic scatters. The shaded curve below shows the expected distribution under complete spatial randomness. The upper curve shows the empirical distribution. Both distributions are clearly separated for all r values.
The segregation test that compared the distribution of penguin nests and artifacts scatters showed significant results (T = 57.671, p-value = 0.009), indicating that there are differences in the distribution of both categories, as observed in the density and quadrat plots.
Likewise, a spatial segregation test was carried out between the presence of nests and vegetation coverage (see also Figure 1), indicating that there was significant segregation between them (T = 928.97, p-value = 0.01). This is of utmost importance because burrowing activity and increased substrate removal in some areas could negatively affect vegetation coverage and increase erosion, as suggested by Ercolano et al. (2016). This suggests that penguin activity may have a relevant influence on this aspect of the landscape.
Discussion
The retrieved evidence suggests that penguins are active agents that modify their environments, generating continuous changes in lithic record. However, spatial analyses indicate that this activity is not random in space but varies in intensity and location, being more prominent in the center of the islet, which also constitutes the sector with the highest relative altitude. However, the lithic artifact record appears to be more restricted to the central sector of the islet, where the highest frequency of findings occurred, while it was absent in the lower sectors. Spatial analyses showed a co-occurrence between both processes, as the accumulation of lithic artifacts occurred in the area with the greatest impact on the landscape by penguins. However, the analysis of spatial segregation reveals that the density of the lithic artifact record is not solely explained by the density of nests, as the latter are distributed more widely on the islet. Conversely, the spatial fingerprints of both species were different, as discarded artifacts were concentrated in a more restricted area. Furthermore, the low density of the lithic artifact record suggests a low discard rate over time (estimated to be around 0.14 and 0.08 artifacts per year). Regarding the correlation between vegetation cover and penguin burrow density, the significant results suggest that either penguins negatively affect vegetation cover through nest excavation or increased erosion from transit or that the observed vegetation cover pattern is linked to other factors not considered in this study.
In addition to the low artifact density, the technological analysis indicated that there is a prevalence of knapping debris and internal flakes, some of which are associated with the preparation and rejuvenation of core platforms. The small proportion of cortex and flakes linked to the early reduction stages suggests that these stages did not take place on the islet. This was also supported by the low occurrence of fractures. Moreover, the overall diversity is comparably low, and the design of the tools is expedient. Although the most common rock in the sample, locally available chert is not found on the islet but sporadically on the coastal strip and rocky outcrops. This suggests that there was a limited but planned use of this area, with the transportation of cores or locally available good-quality rock blanks for producing natural edges and, to a lesser extent, tools, which also showed no signs of maintenance or recycling. The overall patterns in lithic technology differ from those observed at the nearest site located on the coast. Parador QB, in addition to its small sample, has a more diversified representation of raw materials, particularly those with very good knapping quality, such as chalcedony. Likewise, this sector shows clear indications that all stages of core reduction were carried out on-site for all raw materials, except for obsidian, which comes from an extra-areal source (Alberti, 2016; Borella et al., 2020).
Another significant aspect that sheds light on the use of this space is the presence of human burials in the highest sector of the islet (Favier Dubois et al., 2008), some of which have been dated to 2700 BP. The activity of penguins has also aided in identifying possible buried archeological deposits with faunal remains, and further studies are expected to provide more insights into the range of human activities that took place over time in this space.
Regarding morphological alterations, the collected variables accounted for the existence of different moments in the post-depositional history of the sample. The high proportion of materials with carbonate coatings is a useful indicator in surface contexts of recent exposure and the depositional history of the artifacts, as it indicates the subsurface origin of the pieces (Alberti and Carranza, 2014; Carranza, 2022). The substrate that contained the pieces prior to exposure had physicochemical conditions that allowed the formation of precipitates on the surface of the pieces, while they were contained in the sedimentary matrix. In turn, the data obtained from the abrasion profiles show a predominant record of fresh pieces without abrasion, and to a lesser extent, mild abrasion corresponding to stage W1. This inversely proportional presence is consistent with a set that refers to sub-surface conditions characteristic of these aeolian contexts and that has recently been exposed to sub-aerial conditions as a result of the faunal disturbance produced by penguins and the subsequent erosion that triggers this dynamic. The conditions in which the assemblage is found at the time of its recovery indicate that insufficient time has elapsed for the sediment-laden wind to act on the artifacts, first removing the rock coating and subsequently generating rounding and polishing of the edges and surfaces. Therefore, the stability conditions (Borerro, 2004) recorded in this assemblage as a result of the sum of morphological alterations is recent in nature.
The presence of organic coatings, such as excrement, is useful because they present a higher friability alteration on the surface of the rock. Because of their characteristics, they are reversible and indicate a limited and immediate moment at the time of artifact recovery. In this regard, future studies are needed to evaluate whether their presence favors the weathering of pieces and the time involved in this process.
The exposure of lithic artifacts as a result of this dynamic of nesting, unintentional transport “kicking,” trampling and displacement increases visibility but potentially decreases the integrity of the exposed assemblages. Furthermore, it is necessary to increase the sample size both on the islet and along the coast to gain a more detailed understanding of technological variability and formation processes. This includes studying other nearby islets, for which there is no evidence of utilization by hunter-gatherer groups or penguins, but which can be highly useful for naturalistic observations and studying landscape dynamics in particular environments, such as wind erosion and vegetation cover. The distinctive features of limited accessibility and the small size of the Pastosa islet, combined with the role played by hunter-gatherers and penguins in landscape formation, make this area a natural laboratory for investigating archeological landscape dynamics from a taphonomic perspective. Our research highlights the agency of penguins in revealing human spatial usage patterns and transforming lithic landscapes, thus emphasizing the unique nature of this space for archeological research. Finally, the cultural evidence in this area reflects the continuity of using a resource-rich space over time. This is why delving into the knowledge of post-depositional processes and how they can potentially affect the properties of the archeological record allows us to make better decisions in areas protected by environmental and cultural regulations. This area is a clear example of spaces dedicated to the conservation of natural and cultural heritage, research, sustainable tourism, and local development.
Supplemental Material
sj-kml-6-hol-10.1177_09596836231200439 – Supplemental material for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina
Supplemental material, sj-kml-6-hol-10.1177_09596836231200439 for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina by Eugenia Carranza and Marcelo Cardillo in The Holocene
Supplemental Material
sj-txt-2-hol-10.1177_09596836231200439 – Supplemental material for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina
Supplemental material, sj-txt-2-hol-10.1177_09596836231200439 for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina by Eugenia Carranza and Marcelo Cardillo in The Holocene
Supplemental Material
sj-txt-3-hol-10.1177_09596836231200439 – Supplemental material for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina
Supplemental material, sj-txt-3-hol-10.1177_09596836231200439 for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina by Eugenia Carranza and Marcelo Cardillo in The Holocene
Supplemental Material
sj-txt-4-hol-10.1177_09596836231200439 – Supplemental material for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina
Supplemental material, sj-txt-4-hol-10.1177_09596836231200439 for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina by Eugenia Carranza and Marcelo Cardillo in The Holocene
Supplemental Material
sj-txt-5-hol-10.1177_09596836231200439 – Supplemental material for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina
Supplemental material, sj-txt-5-hol-10.1177_09596836231200439 for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina by Eugenia Carranza and Marcelo Cardillo in The Holocene
Supplemental Material
sj-xlsx-1-hol-10.1177_09596836231200439 – Supplemental material for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina
Supplemental material, sj-xlsx-1-hol-10.1177_09596836231200439 for The role of Magellanic penguins (Spheniscus magellanicus) on landscape dynamics and lithic taphonomy in La Pastosa Islet, North Patagonia, Argentina by Eugenia Carranza and Marcelo Cardillo in The Holocene
Footnotes
Acknowledgements
We thank the national park ranger Esteban Tazzioli for field assistance. Our thanks go to Florencia Borella, whose support and reading of this article is very valuable. The National Parks Administration and the Río Negro Province Culture Secretary for granted permission. We would also like to thank the reviewers for the commentaries and suggestions made to the previous version of this paper.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Scientific and Technical Research Council - Argentina [PIP CONICET 112-202101-00908 CO] and National Agency for Scientific and Technological Promotion [PICT ANPCyT 2021-I-A-00436].
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.