
Abstract
Quaternary small mammals are important proxies for paleoenvironmental reconstructions. The Brazilian record, although famous and continuously growing, is hampered by poor chronological control due to the nature of most of the published assemblages. Here, we present a taphonomic analysis and paleoenvironmental reconstruction based on marsupials, bats, and rodents (480 remains) from the Gruta do Presépio archeological site (GPR, hereafter), Santa Catarina State, southern Brazil. The site is a rock shelter located in an Atlantic Forest biome, an important biodiversity hotspot. The GPR sedimentary sequence spans the Early Holocene (8330 ± 30 years 14C BP) to recent Historical time (after the arrival of Europeans). Most of the small mammal remains were deposited by strigiforms, although for the oldest units some strongly digested remains could indicate the activity of diurnal birds or carnivorous mammals. The GPR small mammal assemblages include 28 taxa (five marsupials, nine chiropterans, and 14 rodents). Most of them indicate the occurrence and persistence of forested environments during the Holocene around the site, which is in overall agreement with regional palynological data. A major community change, implying the increment of rodents allied to disturbed ecosystems (e.g. Akodon, Oligoryzomys, Mus), is very recent and reflects human forest clearance as a signature of the Anthropocene.
Introduction
Small mammals are suitable proxies for paleoenvironmental and, to a minor extent, paleoclimatic reconstructions, particularly during the Quaternary (e.g. Andrews, 1990; Avery, 1982; Guillermo et al., 2021; Hadly, 1996; Pardiñas et al., 2022; Thackeray, 1987; Wood and Barnosky, 1994). Taphonomic studies of micromammal remains are important for inferring the main sources of the faunal accumulation, embracing a variety of agents and processes, in order to perform supported reconstructions of the past (e.g. Andrews, 1990; Avery, 1982; Dodson and Wexlar, 1979; Fernández et al., 2019; Guillermo et al., 2021; Montalvo and Fernández, 2019; Pardiñas, 1999b; Stahl, 1996).
For Brazilian cave deposits, foundational studies on the Neotropical Quaternary small mammals and taphonomy have been undertaken. In fact, it was Peter Lund working in the extensive karst system of Lagoa Santa (Minas Gerais) who carried out the first experiments regarding the time implied in the formation of a bony deposit produced by owls and, more precisely, to connect the latter with cave ossuaries (Lund, 1846). During the last two decades, descriptions of numerous Quaternary small mammal assemblages have been published for the country (e.g. de Oliviera et al., 2013; Hadler et al., 2018; Motta et al., 2019; Neves et al., 2017, 2020). However, many of them are limited to taxonomic aspects because they lack, due to the nature of the deposits (e.g. karst systems), a precise chronological framework (e.g. Boroni et al., 2021; Pires et al., 2018). Fortunately, in southern Brazil, several Holocene assemblages from Rio Grande do Sul State have been retrieved from archeological sites with firmly established stratigraphy and absolute dates (Fernández et al., 2019; Hadler et al., 2008, 2009, 2010, 2016; Stutz et al., 2017, 2020a). These findings indicate that both owls and humans were the main accumulating agents of small mammals (Fernández et al., 2019). In addition, the studied sites revealed a clear tendency toward the replacement and regional extinction of grassland species by forest species toward the Late-Holocene (Hadler et al., 2016; Stutz et al., 2017, 2020a).
Recently, a small mammal assemblage from the Holocene of Santa Catarina State, in southern Brazil, was recovered. This state is situated in the Atlantic Forest biome, an important biodiversity hotspot (Marques et al., 2021; Myers et al., 2000). Understanding how this biome will respond to changing conditions in the future is a research priority (Wilson et al., 2021). One way to answer this question is to look at the past, via fossil proxies, to identify the impact of past climate change on the organisms (Fordham et al., 2020; Wilson et al., 2021).
The goal of this paper is to describe this sequence of archeological small mammal assemblages collected from a cave named Gruta do Presépio. Beyond the rich taxonomic sample represented by the studied assemblages, special attention is given to taphonomic analysis and paleoenvironmental reconstruction to assess the effect of past climate change on these micromammal communities.
Archeological context
Gruta do Presépio (GPR, hereafter) is part of an archeological complex comprising three other sites, named Gruta do Tigre, Fenda do Tigre, and Oficina do Tigre, in the municipality of Rio do Oeste, State of Santa Catarina, southern Brazil (27°13′10.50″S, 49°46′ 38.97″W, 442 m asl, Figure 1). GPR is a rock shelter 35 m wide at its mouth and 17 m long that was eroded into a shale outcrop of the Rio do Sul Formation overlain by sandstone of the Rio Bonito Formation (Ballejo et al., 2022; Machado et al., 2019). Eleven archeological units measuring 1 × 1 m and one measuring 1 m × 50 cm were excavated at GPR. The stratigraphy recorded is composed of nine layers differentiated by sedimentological and cultural features (Ballejo et al., 2022; Machado et al., 2019). Layer I was sterile. From layers II to IX a variety of remains were collected, with clear concentrations in layers II and III and VII to IX. The main archeological remains that were detected in all these layers were lithic tools and debris made of sandstone, silicified sandstone, chert, quartzite, and basalt, totaling 238 artifacts (Pereira, 2021). Layers II and III presented an extensive amount of charcoal, ash and burnt sediment, and are rich in fluvial shells, but with a low density of lithic artifacts and other faunal remains (Pereira, 2021).
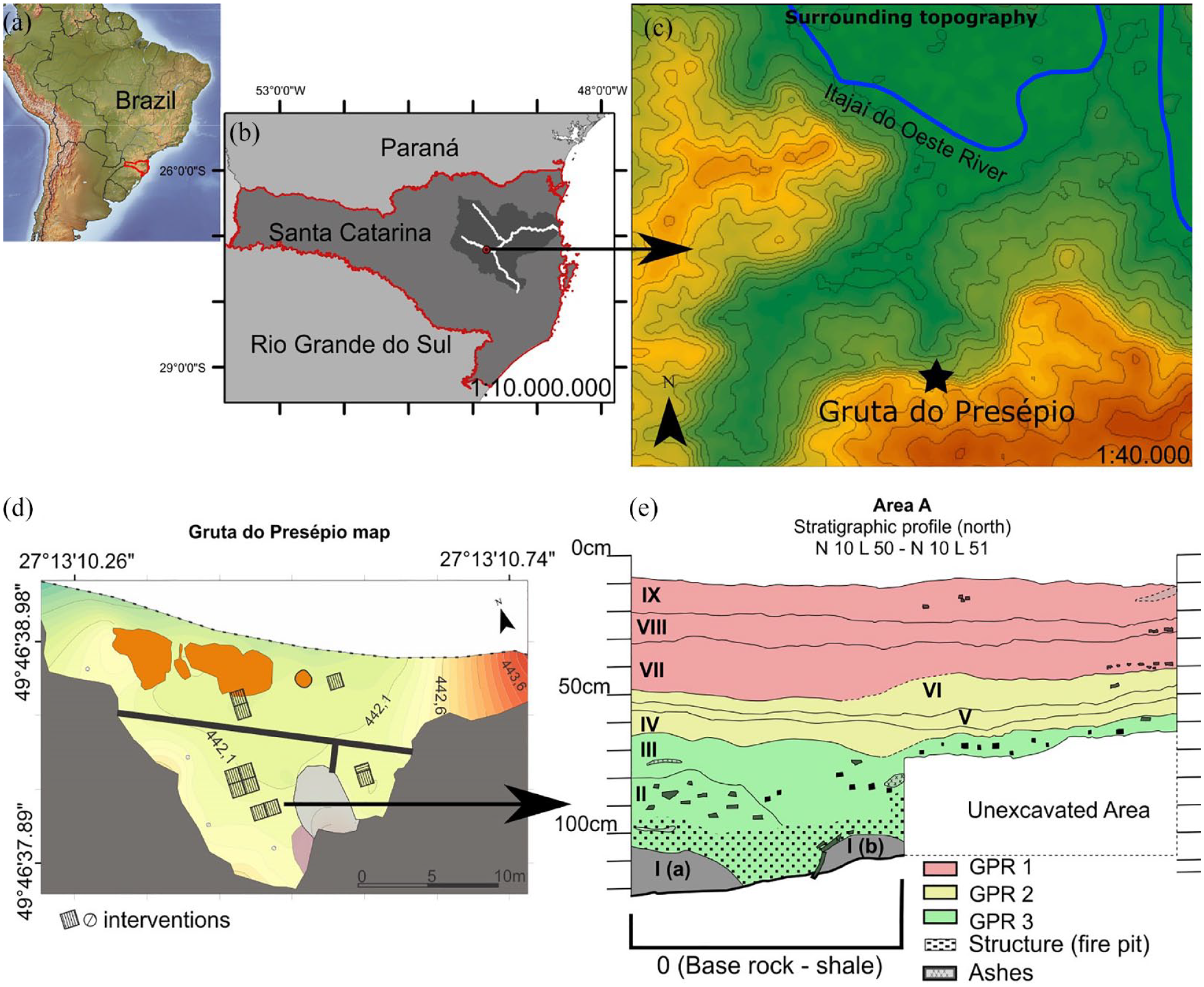
(a) Location of the state of Santa Catarina in South America. (b) Location of Gruta do Presépio in Vale do Itajaí, scale 1:10,000,000. (c) Surrounding topography, scale 1:40,000. (d) Map of the cave and excavation areas, adapted from Machado et al. (2019). (e) Stratigraphic profile of north view of the excavation unit N10 L 50 and N10 L51. Data: ALOS PALSAR, IBGE, ANA, CPRM, Machado et al. (2019) and Pereira (2021).
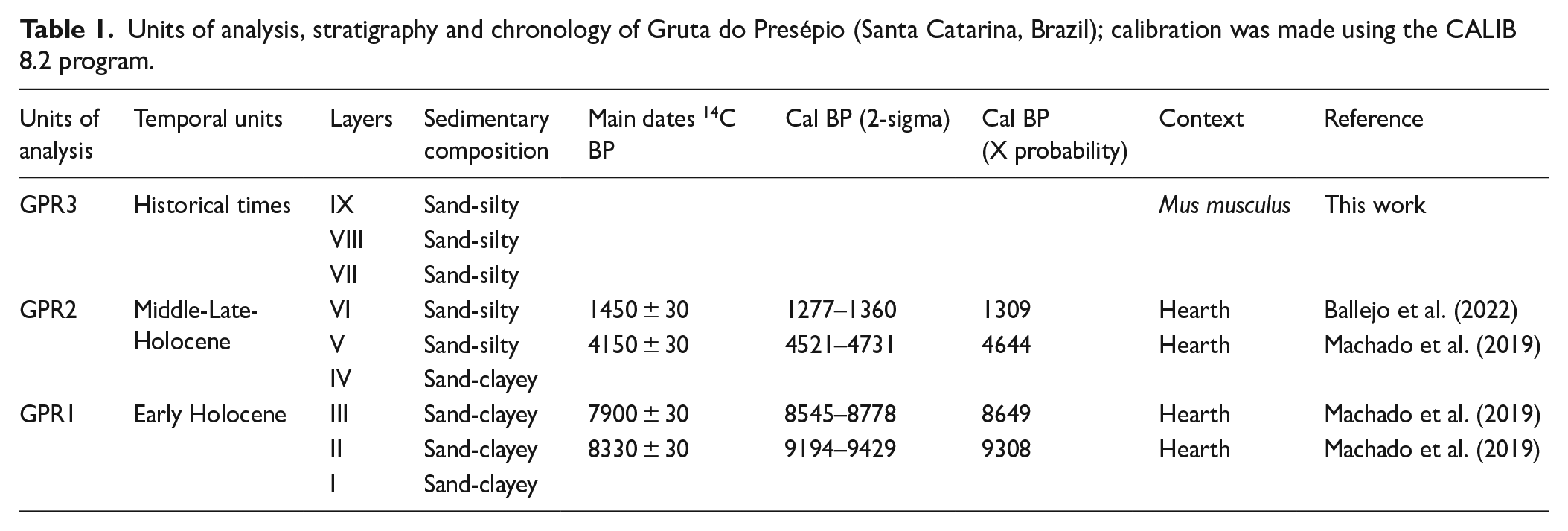
In the archeological complex, dates were obtained only for GPR (Table 1), and point to an extensive, but not continuous, interval of occupation. We defined three main periods of occupation of the site based on stratigraphic and cultural characteristics. The first period, the Early Holocene, presents dates between 8330 and 7750 years BP and covers layers II and III. The second, related to the Mid/Late-Holocene, involves layers IV, V, and VI and has a hiatus between 4150 and 1450 years BP. The third period encompasses layers VII to IX, dating to after the 16th century and being defined based on the occurrence of the exotic rodent Mus musculus starting from layer VII. This chronology, although lacking refinement, inserts GPR into a broader context of occupation in southern Brazil (Dias, 2012; Hadler et al., 2013), adding to the discussion surrounding other areas of the upper Itajaí valley (Reis et al., 2018). These chronological intervals refer to occupations by hunter-gatherer groups from the Early and Mid-Holocene and to the context of occupations linked to southern Jê groups, which began to occupy hillside and plateau areas in southern Brazil at around 3000 years BP (Noelli and Souza, 2017).
Units of analysis, stratigraphy and chronology of Gruta do Presépio (Santa Catarina, Brazil); calibration was made using the CALIB 8.2 program.
The low density of archeological remains associated with the configuration of the fire structures points to an ephemeral use of GPR, with short occupations interspersed with long periods of abandonment. In the Early Holocene, there is more ash, charcoal, and burnt sediment than in other periods, indicating a more intense occupation of the shelter at this time. New excavations are planned to advance the identification of possible areas of activity that could contribute to a better definition of the dynamics of use and formation of the internal space of this site.
Mammal bones were retrieved from layers II to IX. Ballejo et al. (2022) found evidence of scavenger action by New World vultures (Cathartidae) on mammal bone assemblages (NISP = 144 belonging to Dasypus sp., Felidae, Cervidae, and Tayassuidae indet.) recovered from some of the grids located at the entrance of GPR (see Ballejo et al., 2022: fig. 1d and fig. 3). In the same and other grids, medium and large-sized mammals (Cervidae, Tapirus terrestris, Dasypus sp., Puma concolor, and Eira barbara) were also found with evidence of human exploitation (Fernández, personal observation). The mammal assemblage contained in the pellets regurgitated by Cathartidae were found together with other mammal remains discarded by humans that lived in GPR from the Early to Late-Holocene (see Ballejo et al., 2022: fig. 6).
The GPR stratigraphy and chronology are summarized in Table 1. Understanding the gap between the Late-Holocene ages (4150 and 1450 years BP) depends on further dating and excavating more areas.
GPR is located in the Atlantic Forest biome (IBGE, 2019), and the local surrounding vegetation is tropical rainforest of the Alto Vale do Itajaí. This forest is poor in tropical trees, showing great homogeneity in dominant species, such Ocotea spp., Aspidosperma olivaceum, and Copaifera trapezifolia, which present a greater abundance and frequency (Klein, 1978). Currently, only small remnants of this forest type exist in the area, because it has been progressively converted to agricultural land during the last five centuries (Galindo-Leal and Camara, 2003; Vibrans et al., 2012). The climate is mild mesothermal, with average temperatures between 10°C and 15°C, super humid, and without periods of drought (IBGE, 2002).
Material and methods
The sediments extracted from GPR were sieved on site through 2 and 4 mm-sized mesh, and the micromammal remains were separated by manual picking and cleaned with brushes and histology needles. For all layers, constant volume samples were taken, and, in the case of fire structures, full sediment collection was carried out. These samples were floated at the laboratory, and the heavy and light fraction samples were sorted to identify archeological and faunal remains. The studied material, grouped in three chronologically ordered major samples (Table 1), includes 128 fragments of skulls (GPR 1 = 12, GPR 2 = 68, GPR 3 = 48), 77 isolated teeth (GPR 1 = 20, GPR 2 = 41, GPR 3 = 16), and 275 postcranial elements (GPR 1 = 52, GPR 2 = 174, GPR 3 = 49). These remains are housed at the Coleção Arqueológica of the Museu de Arqueologia e Etnologia (MArquE) of the Universidade Federal de Santa Catarina (UFSC), Florianópolis, Brazil.
Only crania and dentaries were used for taxonomy. Identifications were made by comparison with voucher specimens from the Coleção de Mamíferos of the Departamento de Ecologia e Zoologia of the UFSC and of the Seção de Mastozoologia of the Museu de Ciências Naturais da Secretaria Estadual do Meio Ambiente e Infraestrutura, as well as the literature (e.g. Barquez et al., 1999; Cherem et al., 2018; Emmons and Vucetich, 1998; Gardner, 2008; Medellin, 1989; Patton et al., 2015; Stutz et al., 2020a; Voss and Jansa, 2009). The studied material is listed in Supplemental Appendix 1, available online and the specimens used for comparison in Supplemental Appendix 2, available online. The taxonomic framework employed here follows Gardner (2008) for marsupials and bats, and Patton et al. (2015) for rodents. The number of identified specimens (NISP) and the minimum number of individuals (MNI) per taxon were calculated according to Grayson (1984). The NISP was used to access taxonomic abundance (Lyman, 2018). The photographs were taken with the Scanning Electron Microscope in the Laboratório Central de Microscopia Eletrônica of the UFSC.
Paleoenvironmental inferences were supported by comparisons (i.e. presence/absence, relative abundance variation through time) between fossil and recent small mammal assemblages, taking into account the ecological requirements of recorded taxa (e.g. Avery, 1982; Hadly, 1996; Pardiñas, 1999a). The composition of the contemporary community was based on a pellet sample (MNI = 261) produced by the owl Tyto furcata recovered from a nest in the opening of GPR. These materials are housed at the Coleção de Fragmentos Ósseos of the Laboratório de Paleontologia of the UFSC.
Taphonomic analysis was focused on digestive corrosion, breakage, and the relative abundance of elements. The digestive corrosion was considered according to the four categories of damage (light, moderate, heavy, and extreme) described by Andrews (1990), Fernández et al. (2017a), and Montalvo and Fernández (2019) for teeth (isolated and in situ incisors and molars) and postcranial elements (proximal epiphysis of humerus and distal epiphysis of femur). The breakage pattern was identified following the scheme formulated by Andrews (1990). Finally, the relative abundance of each skeletal element was calculated using the formula proposed by Dodson and Wexlar (1979): %Ri = [MNEi/(MNI × Ei)] × 100, where Ri is the relative abundance of the element i; MNE is the minimum number of element i observed in the sample; MNI is the minimum number of individuals; and Ei is the number of the element i in a complete skeleton. Post-depositional features, including weathering, root-marks, and cement crusts, were also analyzed. Weathering was evaluated on a scale from 0 (not weathered) to 3 (extremely weathered) following the stages proposed by Andrews (1990). Root-marks on the bone surface along with cement crusts were classified considering their presence/absence on studied remains. For the cement crust, the petrographic analysis of aggregate sediments (mounted on stubs and coated with gold) was conducted on a JOEL Field Emission Scanning Electron Microscope at the Laboratório Central de Microscopia Eletrônica of the UFSC. An accelerating voltage of 5 kV was used to obtain high-magnification images, and 10 kV was used while carrying out Energy-dispersive X-ray (EDX) analyses.
Results
Taphonomy
Tables 2 to 4 summarize the main taphonomic attributes recorded in the studied samples. Of the analyzed remains (femora, humeri, incisors, and molars = 427; Table 3), 9.1% showed signs of digestive corrosion, with some variations per unit (GPR 1 = 9.4%; GPR 2 = 12.7%; GPR 3 = 3.9%). In all units, most elements were included in the light category, displaying sparse pitting on bone surfaces and enamel reduction or loss of shine (Figure 2a and b). However, some bones and teeth were more damaged (Figure 2c and d), and even a few teeth from GPR 2 and GPR 3 were strongly affected by digestion (Figure 2e–h).
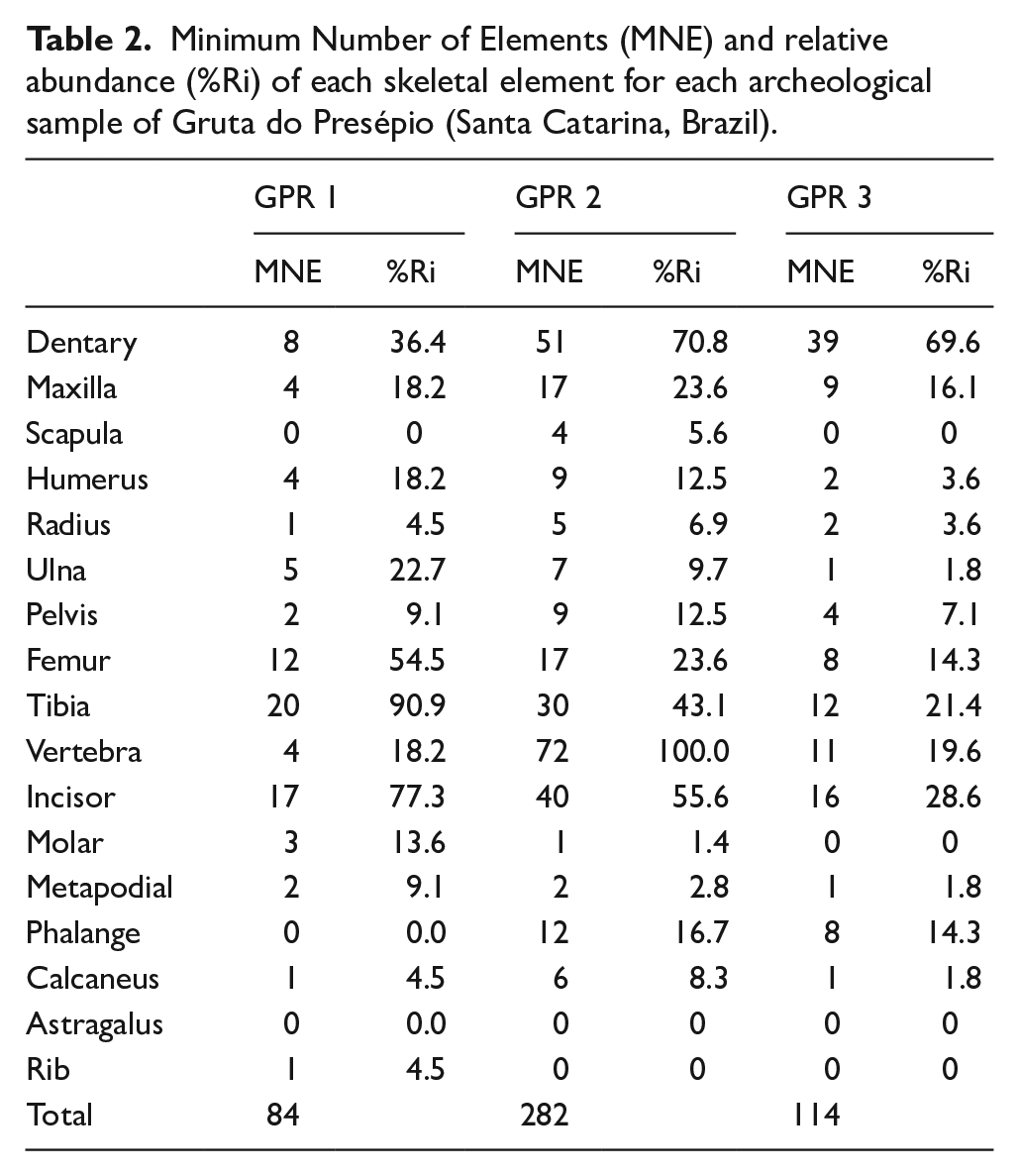
Minimum Number of Elements (MNE) and relative abundance (%Ri) of each skeletal element for each archeological sample of Gruta do Presépio (Santa Catarina, Brazil).
Digestion in teeth (incisors and molars) and postcranial bones (femur and humerus) for each archeological sample of Gruta do Presépio (Santa Catarina, Brazil).
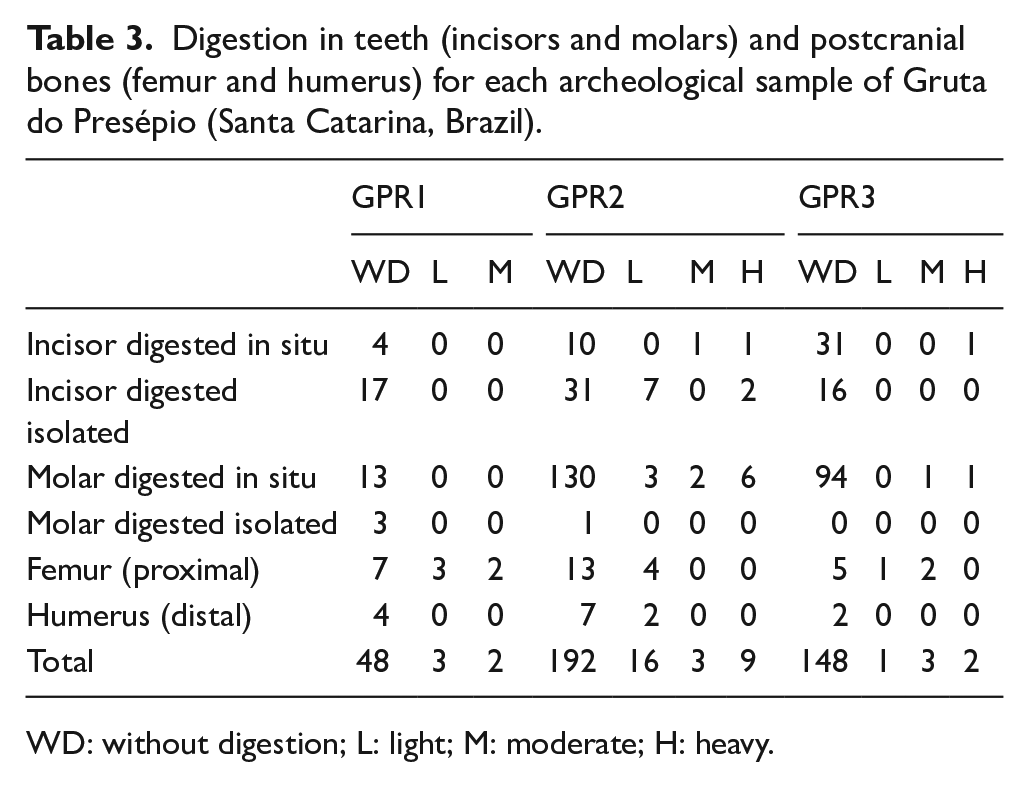
WD: without digestion; L: light; M: moderate; H: heavy.
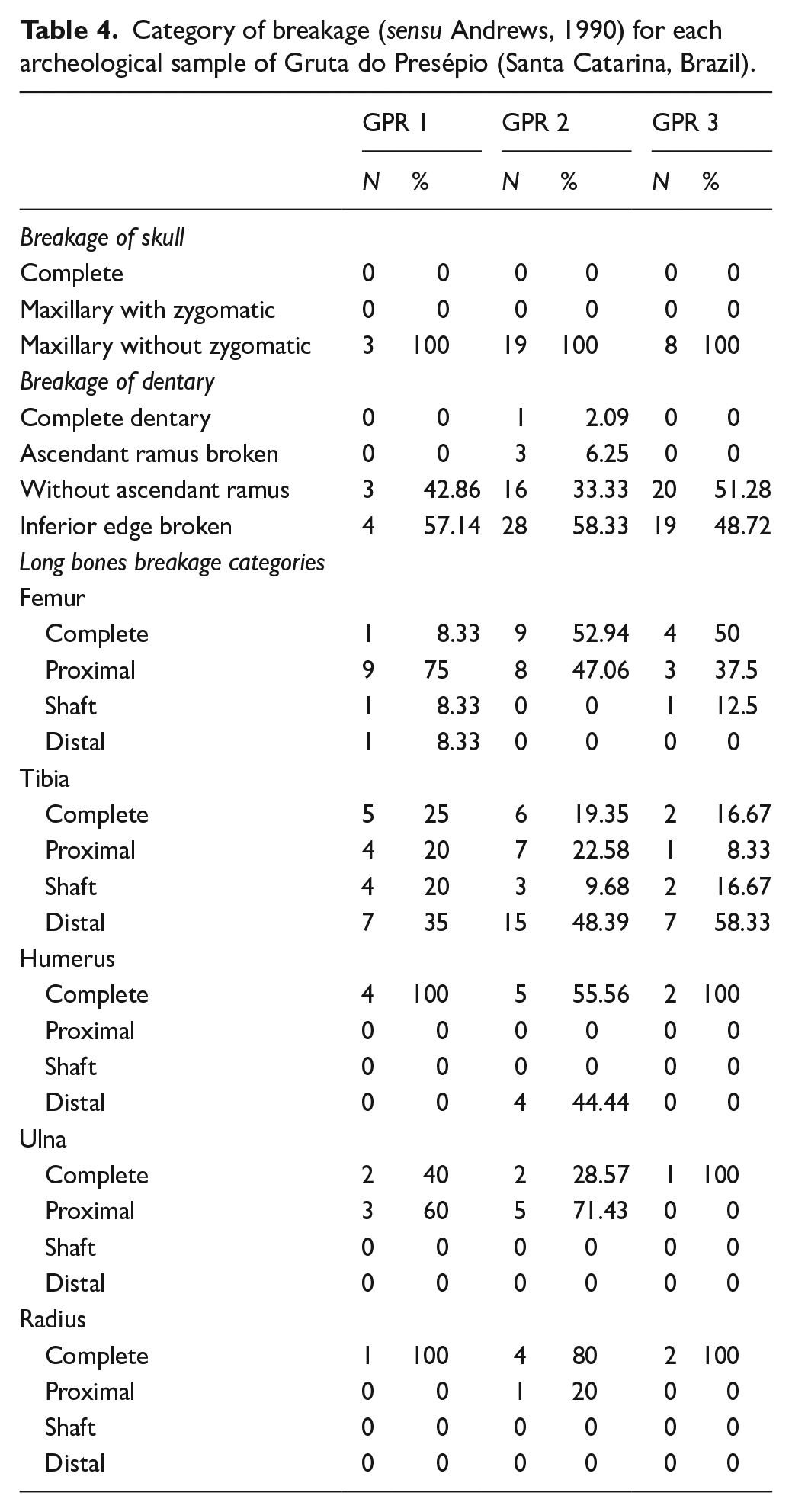
Category of breakage (sensu Andrews, 1990) for each archeological sample of Gruta do Presépio (Santa Catarina, Brazil).

Taphonomic features of the material from Gruta do Presépio (Santa Catarina, Brazil). (a) Incisive of Sigmodontinae (GPR 2). (b) Detail of the light digestion. (c) Femur of Sigmodontinae (GPR 1). (d) Detail of the moderate digestion. (e) Left dentary of Sooretamys angouya (GPR 2). (f) Detail of the heavy digestion on m1. (g) Right dentary of Monodelphis scalops (GPR 3). (h) Detail of the heavy digestion. (i) Right dentary of Gracilinanus microtarsus (GPR 3) with roots marks. (j) Rostral part of the cranium of Juliomys sp. (GPR 2) with cement deposition. (k) Right dentary of G. microtarsus (GPR 2) with cement deposition.
Most of the bones (80.4%) were broken, showing sharp edges and rough surfaces, in all units (Table 4). All crania were heavily fractured (only the maxillae were preserved) and just one dentary was complete, while most lacked the ascending rami. In addition, 63.2% of the analyzed postcranial bones were broken (Table 4).
The relative abundance of skeletal elements reveals a high frequency of dentaries, maxillae, tibiae, and femora (Figure 3). The average of the relative abundance was very low in all units (Table 2).
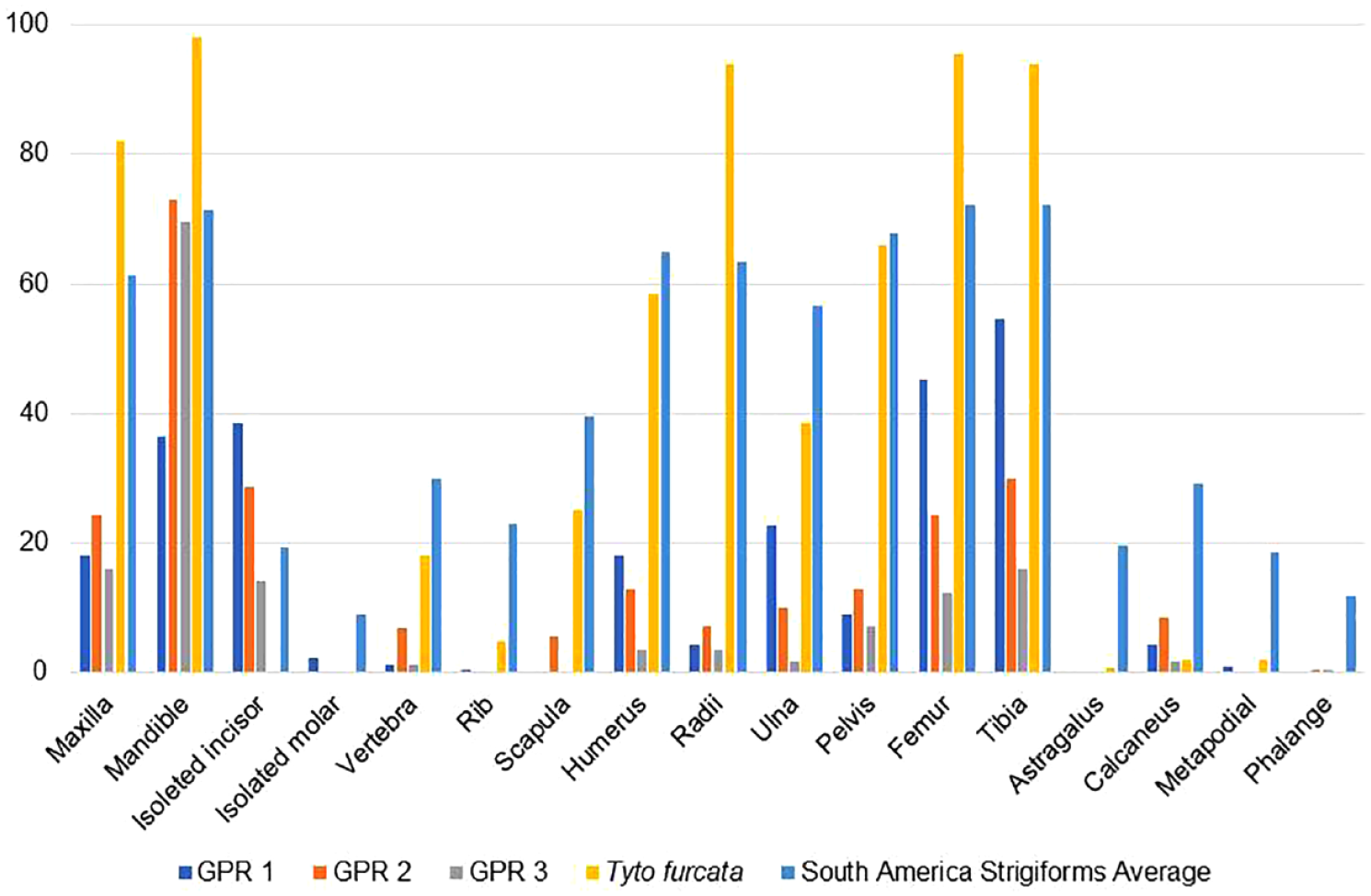
Relative frequency of each skeletal element for each archeological sample of Gruta do Presépio (Santa Catarina, Brazil).
Considering the post-depositional processes, neither weathering nor abrasion produced by water/wind transport were registered in the samples analyzed. Conversely, bones with root etching were found (Figure 2i), mainly in GPR 2 (GPR 1 = 3.1%; GPR 2 = 9.8%; GPR 3 = 0.4%). In addition, bones and teeth with cement crusts were recorded in all units (Figure 2j and k), mostly in the upper one (GPR 1 = 6.9%; GPR 2 = 7.3%; GPR 3 = 12.3%). The analysis by EDX revealed the main constituent minerals of the sediments adhered to the surface of the bones to be gypsum (CaSO4·2H2O), fluorapatite (Ca5(PO4)3F), illite (K, H3O)(Al, Mg, Fe)2 (Si, Al)4O10[(OH)2·(H2O)], feldspars, detrital, and diagenetic quartz (SiO2), and Ti-oxide (TiO2).
Taxonomic diversity and paleoenvironment
The studied GPR small mammal assemblages comprised 28 taxa, including five marsupials, nine bats, and 14 rodents (Table 5, Figure 4). Due to several reasons, including the fragmentary nature of the analyzed material or lack of representation of diagnostic characters, several taxa were determined only to generic level (e.g. Anoura sp., Histiotus sp., Delomys sp., Juliomys sp.).
Taxonomic composition of the small mammal assemblages studied from Gruta do Presépio (Santa Catarina, Brazil).
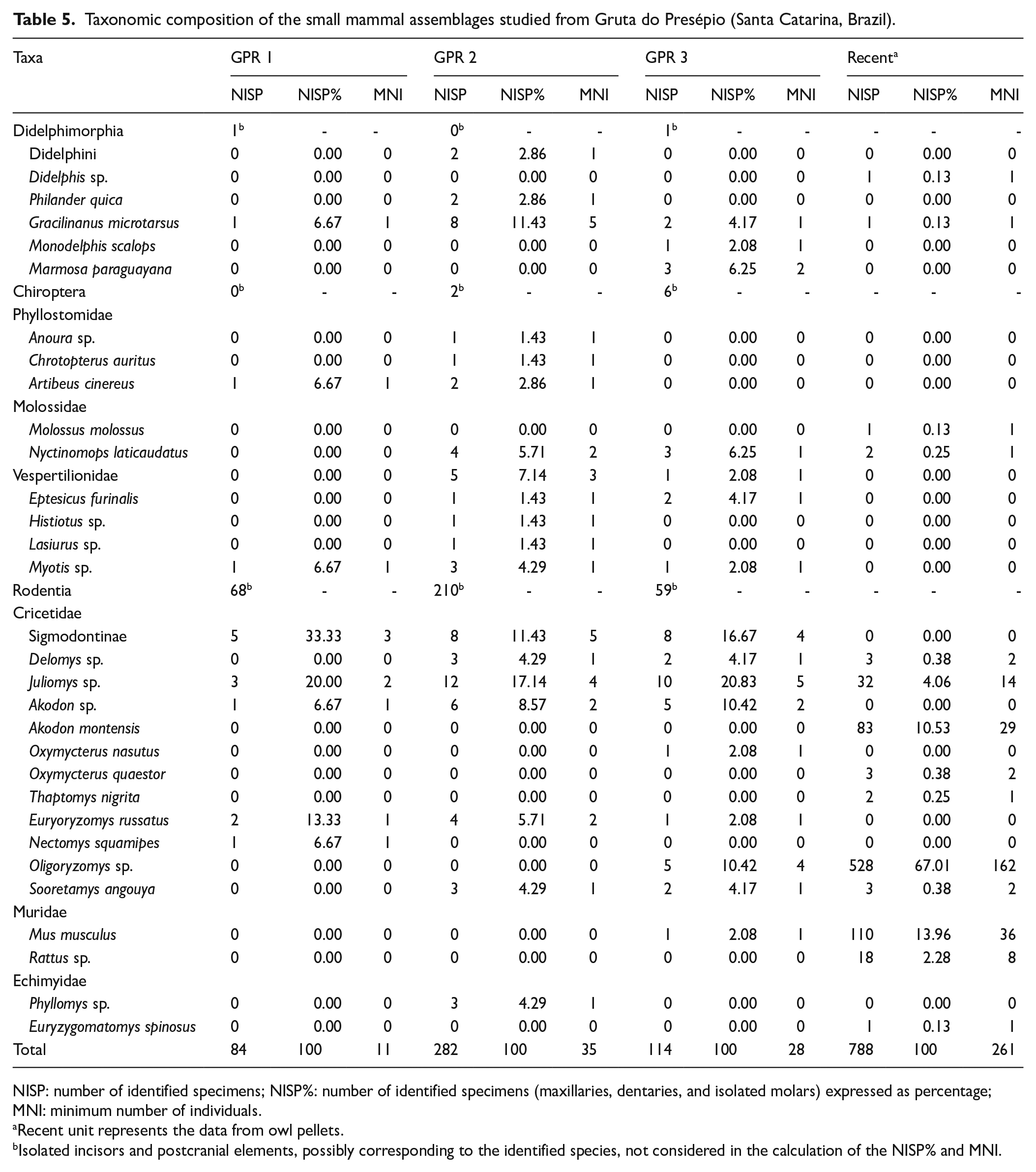
NISP: number of identified specimens; NISP%: number of identified specimens (maxillaries, dentaries, and isolated molars) expressed as percentage; MNI: minimum number of individuals.
Recent unit represents the data from owl pellets.
Isolated incisors and postcranial elements, possibly corresponding to the identified species, not considered in the calculation of the NISP% and MNI.
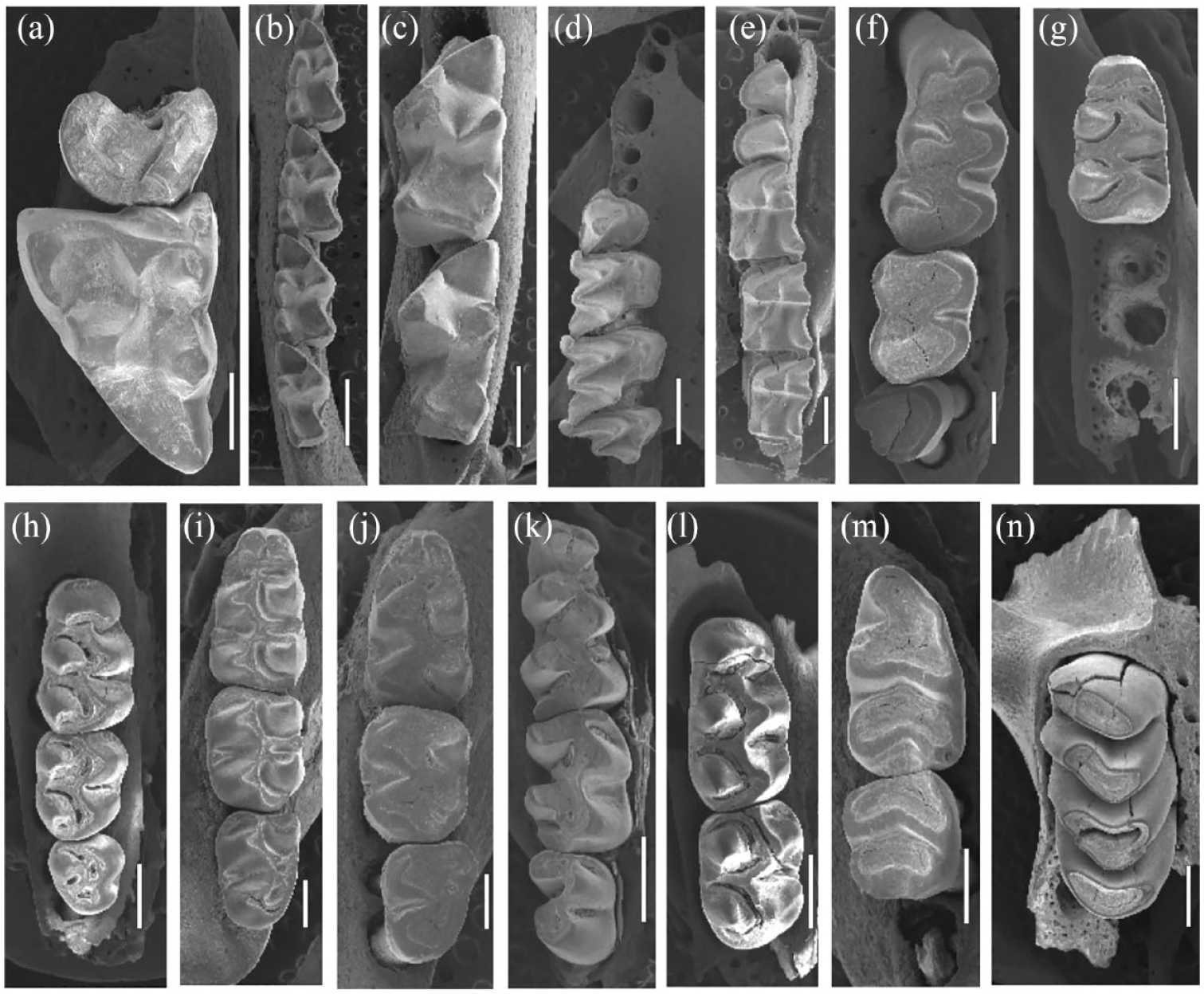
Specimens from Gruta do Presépio (Santa Catarina, Brazil). (a) Philander quica (GPR 2), left maxilla com M1. (b) Gracilinanus microtarsus (GPR 2), right dentary with m1–m4. (c) Marmosa paraguayana (GPR 3), left dentary with m3–m4. (d) Myotis sp. (GPR 2), right maxilla with P3–M3. (e) Nyctinomops laticaudatus (GPR 2), left dentary with p2–m3. (f) Akodon sp. (GPR 3), right maxilla with M1–M3. (g) Delomys sp. (GPR 2), right dentary with m1. (h) Euryoryzomys russatus (GPR 2), right maxilla with M1–M3. (i) Juliomys sp. (GPR 3), left dentary with m1–m3. (j) Oligoryzomys sp. (GPR 3), left dentary with m1–m3. (k) Oxymycterus nasutus (GPR 3), right dentary with m1–m3. (l) Sooretamys angouya (GPR 3), right maxilla with M1–M2. (m) Mus musculus (GPR 3), right dentary with m1. (n) Phyllomys sp. (GPR 2), right maxilla with P4. Scales bars (a–e, g, h, k, l, and n) = 1mm; (f, i, j, and m) = 500 µm.
Rodents are the most frequent taxon in the studied samples, surpassing 90% of NISP in the recent assemblage. Marsupials and chiropterans are more abundant in GPR 2 (representing 17% and 27%, respectively) than in the other units. Marsupials are equally diverse in GPR 3 and GPR 2, while Chiroptera is more diverse in GPR 2. Gracilinanus microtarsus, Akodon, and Juliomys sp. are the only taxa recorded in all units, including the recent sample. Echimyidae is recorded in GPR 2 (Phyllomys sp.) and in the recent assemblage (Euryzygomatomys spinosus) by one individual in each (Table 5). Five genera (Didelphis, Molossus, Thaptomys, Rattus, and Euryzygomatomys) are recorded only in the recent assemblage.
Only seven taxa are registered in GPR 1. It is characterized by a high frequency of forest taxa, such as Juliomys sp. (20% of NISP) and Euryoryzomys russatus (13.3% of NISP). GPR 2 encompasses 18 taxa, and the most frequent are Gracilinanus microtarsus (11.4%) and Juliomys sp. (17.1%). GPR 3 encompasses 15 taxa, and Juliomys sp. (20.8% of NISP) is the most frequent taxon, followed by Oligoryzomys sp. and Akodon sp. (10.4% each). Only one specimen of Mus musculus (2.08%), an exotic species, was recorded in GPR 3. In the recent assemblage, 14 taxa are recorded with Oligoryzomys sp. the most frequent (67.0%), followed by Mus musculus (13.96%) and Akodon montensis (10.5%) (Table 5).
The occurrence of species associated with forest environments, such as Gracilinanus microtarsus, Monodelphis scalops, Marmosa paraguayana, Philander quica, Delomys sp., Juliomys sp., Sooretamys angouya, Euryoryzomys russatus, and Phyllomys sp., is observed thorough the GPR sequence (Table 5). Two sigmodontine rodents that became dominant in human-modified ecosystems, Akodon sp. and Oligoryzomys sp., showed a dramatic increase in very recent times (Figure 5b). Although the studied samples are small, in particular GPR 1, forest dwellers are consistently present (Figure 5a); on the contrary, micromammals indicating grasslands or open environments, such as Thylamys, Bibimys, Calomys, Necromys, etc., lack a record.
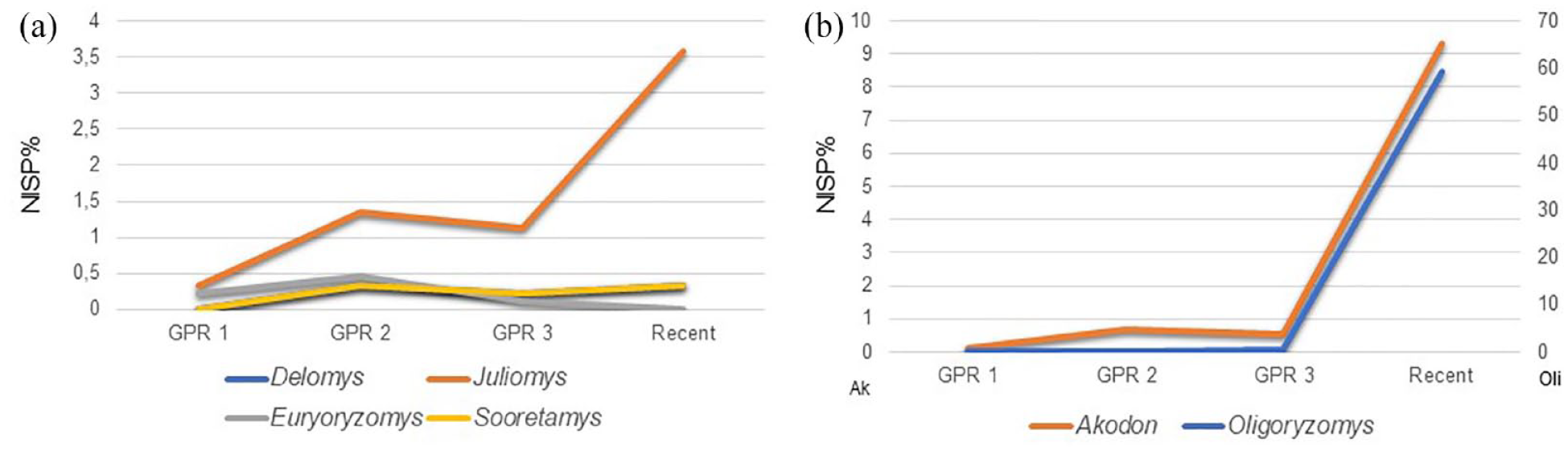
Temporal changes in the relative abundance (NISP%) based on a total NISP = 892 of four forest dwellers rodents (a) and two generalist rodents (b) from Gruta do Presépio (Santa Catarina, Brazil).
Discussion
Taphonomy
The absence of cut-marks and burnt remains suggests non-human accumulation for the analyzed samples in GPR (Fernández et al., 2017a, and references therein). Conversely, the practice of human consumption of small mammals has been recorded in Holocene archeological sites from Rio Grande do Sul State in the extreme south of Brazil (Fernández et al., 2019).
The finding of mostly light digestive corrosion on a low percentage of teeth and postcranial bones is in agreement with an accumulation produced by the nocturnal birds of prey – Strigiformes (e.g. Andrews, 1990; Fernández et al., 2017b; Montalvo and Fernández, 2019). Among the various species of owls that inhabit the study area, the barn owl, Tyto furcata, is the raptor that produces lower levels of digestion and preys on a variety of vertebrates similar to that recorded in GPR (e.g. Cherem et al., 2018; Stutz et al., 2020b). However, the fact that some strongly digested remains have been found in GPR 2 and GPR 3 could also indicate the participation of a different type of predator. Much greater modifications on bones and teeth are exerted by diurnal birds of prey – Accipitriformes – and carnivorous mammals (Montalvo and Fernández, 2019, and references therein). In fact, New World vultures have been documented as accumulators of medium and large mammal remains retrieved from GPR (Ballejo et al., 2022). It is largely known that archeological records are palimpsests on which different taphonomic agents intervene (e.g. Bailey, 2007; Ballejo et al., 2022; Guillermo et al., 2021; Lucas, 2005).
The studied remains from GPR are characterized by the absence of weathering and wind and/or hydraulic transport, suggesting a rapid introduction into the sedimentary deposit. Herbaceous cover, mainly during the deposition of the Mid–Late-Holocene sediments, is suggested by the greater root etching on bones registered in GPR 2 (Andrews, 1990). However, the broken elements with sharp angles and rough edges suggest a trampling action instead of digestion (smooth edges and obtuse fracture angles), a common taphonomic process in caves because they are restricted circulation spaces (Andrews, 1990; Pardiñas, 1999b).
Regarding diagenesis, the cementation of gypsum, a common mineral in extensive evaporitic sedimentary rock beds rarely found in the form of sand, recorded on bones and teeth in GPR, indicates oxidizing conditions. Fluorapatite is a common phosphate mineral that occurs widely as an accessory mineral in calcium-rich igneous or metamorphic rocks (Mange and Maurer, 1992). It can also occur as a detrital or diagenetic mineral, or as a residual product in lateritic soils (Weissbrod and Nachmias, 1986). Illite is a group of non-expandable clay minerals produced from the alteration of muscovite or feldspars in arid or hydrothermal environments. Feldspars are igneous rock-forming minerals that contain cations such as Na2+, Ca2+, and K+, and are very common as detrital or diagenetic phases in deposits or sedimentary rocks (Tucker, 2001).
Community change during the Holocene
All the species recorded in GPR occur, or potentially occur (Cherem et al., 2004), at present in the studied area. GPR 1, GPR 2, and GPR 3 are predominantly composed of forest taxa, such as Gracilinanus microtarsus, Monodelphis scalops, Marmosa paraguayana, Philander quica, Delomys sp., Juliomys sp., Sooretamys angouya, Euryoryzomys russatus, and Phyllomys sp. (Gardner, 2008; Patton et al., 2015; Pavan and Voss, 2016). The frequency of Gracilinanus microtarsus and the diversity of bats in GPR 2 are remarkable. Currently, for southern Brazil there are no cases like this in recent pellet samples (e.g. Cherem et al., 2018; da Silveira et al., 2021; Stutz et al., 2020b). The predominance of Akodon and Oligoryzomys in the contemporary community is clear, plus the exotic murid Mus musculus. These latter taxa first appear in GPR 3, at very low frequencies, especially Mus. Thus, we can observe a change in taxonomic composition from the Early Holocene toward recent Historic time and the present assemblage. The latter is very similar taxonomically to other assemblages produced by barn owls in Santa Catarina State (e.g. Cherem et al., 2018; da Silveira et al., 2021), with Oligoryzomys and Mus being the most frequent mammals preyed upon. This reveals a homogenization of micromammal assemblages, characterized regionally by the dominance of a few opportunistic taxa, a general impoverishment, and the extirpation of specialist species. It is largely known that such kinds of features typify worldwide human-influenced communities characterizing the Anthropocene (e.g. Fusco et al., 2016; Geri et al., 2010; Pardiñas, 1999c; Pardiñas et al., 2000, 2012; Rowe and Terry, 2014; Teta et al., 2014).
For the Holocene of Brazil there are few studied small mammal communities with precise chronological control (Hadler et al., 2013). Against this backdrop, three well-known assemblages from the northwestern Rio Grande do Sul State (i.e. the Garivaldino, Sangão, and Pilger archeological sites) are important. These deposits cover the Holocene from 9430 years BP to 3000 years BP, overlapping with almost the entire sequence of GPR (Fernández et al., 2019; Hadler et al., 2013). A rich pool of species is verified from these deposits, including those from open areas (e.g. Dicolpomys fossor, Proclinodontomys mordax, Ctenomys, Cavia, Calomys), with others common in forests (e.g. Delomys, Sooretamys, Phyllomys; Hadler et al., 2008, 2009, 2013, 2016; Stutz et al., 2017, 2020a). This taxonomic composition and the differential chronological distribution of the taxa recorded suggest an important environmental heterogeneity characterizing the entire Holocene (Hadler et al., 2013). The extinctions in the Rio Grande do Sul assemblages are noteworthy and include regional extirpations (e.g. Kunsia tomentosus and Gyldenstolpia) and biological losses (e.g. Dicolpomys fossor, Proclinodontomys mordax and Clyomys riograndensis; Fernández et al., 2019; Hadler et al., 2016; Stutz et al., 2017, 2020a). On the contrary, the assemblage and environmental evolution reflected by the assemblages recorded in GPR suggest community stability through time. For example, there is no evidence of extinctions or pseudoextinctions at this site. Although the potential taphonomic biases must be considered in interpreting these results (several of the extinguished forms in Rio Grande do Sul are clearly associated with human exploitation; Fernández et al., 2019), the GPR samples point to environmental continuity during the Holocene.
Paleoenvironments in southern Brazil: Combining evidence
Palynological data from Santa Catarina State are concentrated in coastal plains (e.g. Behling and Negrelle, 2001; Cancelli et al., 2012a, 2012b; Kuhn et al., 2023; Val-Peón et al., 2019). For the highlands, there are studies from Northern Santa Catarina State Plateau (Oliveira et al., 2006), Serra Geral (Behling, 1995) and Serra do Tabuleiro State Park (de Oliveira et al., 2012; Jeske-Pieruschka et al., 2013), all of the sites having altitudes over 850 m. Jeske-Pieruschka et al. (2013) pointed out that the dense ombrophilous forest moved from the lowlands over the slopes in the Early Holocene due to warmer and wetter conditions. Herbaceous plants decreased from the Mid–Late-Holocene onward, and arboreal taxa increased due to the wetter conditions, showing the expansion of the forest (de Oliveira et al., 2012). This wetter condition in the Late-Holocene was corroborated by trace-element analysis of a speleothem from Botuverá cave, which showed that the total amount of rainfall was higher when compared to the Early and Mid-Holocene (Bernal et al., 2016). Despite that, grassland was present during the Late-Holocene, indicating a mosaic environment. The dense ombrophilous forest reached its maximum expansion after 1500 cal. years BP. The Araucaria Forest began to develop more recently after 2000 years BP and expanded twice during the Late-Holocene (Jeske-Pieruschka et al., 2013). Thus, the dense ombrophilous forest expanded three times in Santa Catarina State (in the Early Holocene, during the Mid/Late-Holocene, and in the Late-Holocene). It differs from the highlands of Rio Grande do Sul, where the initial development started in the Mid-Holocene, indicating that the dense ombrophilous forest expanded from the north to more southerly regions (Jeske-Pieruschka et al., 2013).
In southern Brazil, the expansion of Araucaria Forest seems to be related to indigenous southern Jê populations (e.g. Bitencourt and Krauspenhar, 2006; Reis et al., 2014; Robinson et al., 2018). On one hand, the general wetter conditions favored the expansion of forests in the Late-Holocene on the south-facing slopes in a mosaic with grasslands (Jeske-Pieruschka et al., 2013). On the other hand, human activities were important in expanding forests beyond their natural habitat distribution boundaries (north-facing slopes) (Robinson et al., 2018). Unfortunately, the role of human populations in the dense ombrophilous forest expansion, if any, is not yet understood.
The palynological data are, overall, in accordance with data furnished by the small mammal assemblages recorded in Rio Grande do Sul and Santa Catarina. In the Garivaldino, Pilger, and Sangão sites, grassland taxa are common throughout the entire sequences, and the forest taxa begin to appear from about 6000 years BP (Fernández et al., 2019; Hadler et al., 2013, 2016; Stutz et al., 2017, 2020a). In GPR, forest small mammal taxa were present in the last 8400 years BP and they dominate the samples until recently. Medium to large-sized mammals recorded in GPR also corroborate this finding, since the tapirid Tapirus terrestris, the mustelid Eira barbara, and the procyonid Nasua nasua occur predominantly in forests, occasionally feeding in or crossing open environments, including agricultural fields and grasslands (Ballejo et al., 2022).
Holocene climate variations, although with lower amplitudes of temperature than during glacial times, were larger and more frequent than is commonly recognized (Mayewski et al., 2004; Urrego et al., 2009; Wanner et al., 2015). The Holocene can roughly be subdivided into four periods: a progressively warming Early Holocene (between about 11.7 and 7–8 ka BP), above all in the Northern Hemisphere; a warm Mid-Holocene (between 7–8 and 4–5 ka BP); a cooler Late-Holocene (between 4 and 5 ka BP and the 19th century AD); and the modern, warm Industrial Era since about AD 1860 (Wanner et al., 2015). For eastern South America the records summarized by Gorenstein et al. (2022) suggest an overall drier Mid-Holocene climate compared to the Late-Holocene. Our data suggest that Holocene climate changes had different effects on the environments of the Atlantic Forest biome. While in Rio Grande do Sul, the meridional limit of the biome, the forests expanded later, in Santa Catarina expansion started earlier, maintaining an environmental stability over the Holocene.
Supplemental Material
sj-docx-1-hol-10.1177_09596836231200440 – Supplemental material for Forest stability during the Holocene in Santa Catarina, southern Brazil revealed by small mammals from Gruta do Presépio
Supplemental material, sj-docx-1-hol-10.1177_09596836231200440 for Forest stability during the Holocene in Santa Catarina, southern Brazil revealed by small mammals from Gruta do Presépio by Patrícia Hadler, Fernando J Fernández, Jorge José Cherem, Juliana Machado, Lucas Bueno, Thiago Umberto Pereira, Marivaldo Nascimento and Ulyses Francisco José Pardiñas in The Holocene
Supplemental Material
sj-docx-2-hol-10.1177_09596836231200440 – Supplemental material for Forest stability during the Holocene in Santa Catarina, southern Brazil revealed by small mammals from Gruta do Presépio
Supplemental material, sj-docx-2-hol-10.1177_09596836231200440 for Forest stability during the Holocene in Santa Catarina, southern Brazil revealed by small mammals from Gruta do Presépio by Patrícia Hadler, Fernando J Fernández, Jorge José Cherem, Juliana Machado, Lucas Bueno, Thiago Umberto Pereira, Marivaldo Nascimento and Ulyses Francisco José Pardiñas in The Holocene
Footnotes
Acknowledgements
To P. C. Simões Lopes and M. E. Graipel for access to the material of the Coleção de Mamíferos of the Departamento de Ecologia e Zoologia of the Universidade Federal de Santa Catarina. To the Laboratório Central de Microscopia Eletrônica of the UFSC for the SEM photographs. To the anonymous reviewers and Francis Mayle for their contributions to improving the manuscript. To Katarzyna Piper for English revision.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Fundação de Cultura Catarinense – Edital Elisabete Anderle 2017; CNPq – Edital Universal 28/2018 (grant number 40953/2018-7); and Escala Docente Program from Asociación de Universidades Grupo Montevideo.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.