
Abstract
Archaeological findings from the Neolithic open-air location Ayakagytma ‘The Site’, situated in the south-eastern part of the Kyzyl-kum Desert in Uzbekistan, can potentially shed new light on the camel domestication process in the Central Asian drylands and help to connect it to regional changes of paleoclimate. Detailed analyses of composition and 13C isotopic ratios of fatty acids performed on potsherds from an archaeological horizon of a Keltaminar culture dated at 3000–4000 cal BC combined with analogical analyses of modern camel and horse milk samples from Uzbekistan indicated a plausible possibility that camels were kept and milked by stockbreeders of Ayakagytma during this time period. The observed herding practices based almost exclusively on camel husbandry, as opposed to earlier more balanced herds of cattle, horses and camels, were probably an adaptation to climate transition from relatively humid to relatively dry, and the following changes in vegetation. Such climatic shift did not correspond with the general trend of Holocene moisture changes over the Westerlies-dominated Central Asia. However, it was in accordance with wet-to-dry climate transitions recorded in sediments of several lakes in the same region at around 4000 cal BP. The observed changes in Neolithic stockbreeding practices, as other inconsistencies in Holocene moisture evolution over a massive area of arid Central Asia, may have resulted from local manifestations of globally-forced climate changes and/or from local hydrographic alterations unrelated to paleoclimate.
Introduction
For millennia camels have been used as multifunctional animals, providing means of transportation and haulage, as well as a source of milk, meat and fleece. In Central Asia, Bactrian camels became the dominant economic factor in the second millennium BC, as they are well-adapted to desert environments and enabled long-distance trade with Europe (e.g. Kuzmina, 2015; Peters and von Den Driesch, 1997). Regardless of their socio-economic importance, our knowledge concerning the process and location of Bactrian camel domestication remains ambiguous, mostly due to the limited occurrence of their remains in well-dated archaeological contexts (e.g. Burger et al., 2019; Kuzmina, 2015; Orlando, 2016). Despite the genetic evidence of a single domestication process (Trinks et al., 2012), two potential regions of Bactrian camel domestication were suggested – northeastern Iran and southwestern Turkmenistan, characterized today by the highest genetic diversity of Bactrian camels (Burger et al., 2019; Kuzmina, 2015); or areas located further to the east, for example, Kazakhstan or northwestern Mongolia, where people lived alongside wild camels for an extended period (Burger et al., 2019).
Fortunately, excavations carried out by the Polish-Uzbek Archaeological Expedition between 1995 and 2004 in the Neolithic open-air location Ayakagytma ‘The Site’, situated in the south-eastern part of the Kyzyl-kum Desert in Uzbekistan, can shed new light on this dilemma. This location is a multilayered, stratified site of the Neolithic entity called Keltaminar culture (Szymczak et al., 2006). The Keltaminar populations were nomadic stockbreeders who settled mainly in desert regions of Central Asia (Lasota-Moskalewska and Szymczak, 2012). In Ayakagytma ‘The Site’ two chronological horizons of the local Neolithic could be distinguished: the early (Dariasai) phase, 14C dated at ca. 6100–5400 cal BC and the middle (Tuskan) phase, 14C dated at ca. 4000–3000 cal BC (Szymczak et al., 2006). The chronological gap between the horizons resulted probably from water level shifts in a paleolake located near Ayakagytma ‘The Site’, that led to its flooding between 5400 and 4000 cal BC (Szymczak and Khudzhanazarov, 2009, 2011). Interestingly, when after 4000 cal BC Keltaminar population returned to Ayakagytma ‘The Site’, a significant change in composition of zooarchaeological assemblages was recorded. During the early settlement phase, they were dominated by cattle and horse remains (30% and 34% of the identified animal remains, respectively), with slightly lower share of camel remains (23%) (Lasota-Moskalewska and Szymczak, 2012). After 4000 cal BC, during the middle phase of Keltaminar culture, the composition of zooarchaeological assemblage changed radically and became almost entirely dominated by camel remains, comprising approximately 85% of the identified remains, with other animals constituting less than 5% each. Thus, we may expect that after 4000 cal BC the process of camel domestication became more advanced in the Keltaminar population. However, the role of camel milk in the Keltaminar diet remained uncertain (Lasota-Moskalewska and Szymczak, 2012). Interestingly, though today consumption of fermented camel milk is widespread in Central Asia, no evidence has been found so far of camel milk used by the Neolithic pastoralists in this region.
Therefore, we studied chemical characteristics of lipid residue preserved in available pottery shreds excavated from the younger horizon at the Ayakagytma ‘The Site’ in search for evidence that camels were milked by the Keltaminar culture at least during the Tuskan phase of the local Neolithic (4000–3000 cal BC). Moreover, as the domination of camels in herds may be viewed as an adaptation to dry climatic conditions, we analysed available literature data on Holocene climate evolution in the Westerlies-dominated arid Central Asia (as defined by Chen et al., 2008; Huang et al., 2014) to explore potential climatic and/or environmental drivers of a drastic change in stockbreeding practices of Keltaminar culture at around 4000 cal BC.
Materials and methods
General characteristics of the study area
The Kyzyl-kum desert covers about 300,000 km2 to the north-east of the Amu-Darya river, over the territory of the Republics of Kazakhstan, Uzbekistan and Turkmenistan. According to the Köppen-Geiger classification (Kottek et al., 2006), the Kyzyl-kum desert is characterized by a cold desert climate, with features of a cold steppe climate noticeable in its south-eastern outskirts. Currently, mean air annual temperatures (MAAT) in the Kyzyl-kum desert are between 11oC and 16oC, and mean annual precipitation (MAP) between 50 and 100 mm (Gintzburger et al., 2003). The highest values of precipitation are recorded in winter and an additional smaller peak may occur in spring, in summer a drought is observed (Gintzburger et al., 2003). The Kyzyl-kum desert is predominantly covered with recent aeolian sand dunes and sandy-loam/clay formations, with saline depressions in between them. On sandy soils typical for the dunes develops psammophytic vegetation, including (1) small trees, shrubs and small shrubs (e.g. Haloxylon spp., Salsola spp., Astragalus spp., Ammodendron spp., Acanthophyllum spp., Halothamnus spp.), making up to 30% of the flora composition; (2) annual summer plants (e.g. Agriophyllum sp., Climacoptera spp. Corispermum spp.), constituting 40% of the flora composition; (3) graminoids (e.g. Aristida pennata and A. karelinii, Carex physodes, Poa bulbosa), making up 20% of the flora composition; and (4) ephemeroids and ephemerals (e.g. Ferula assa-foetida, Eremopyrum spp., Malcolmia spp., Isatis spp.), constituting about 10% of the flora composition (Gintzburger et al., 2003; Kapustina, 2001). Vegetation on sandy dunes provides the major source of fodder for sheep and camels (Gintzburger et al., 2003). On grey-brown soils of sandy-loam and clay formations develops poorly diversified gypseous vegetation, dominated by species from the families Chenopodiaceae and Amaranthaceae, such as Salsola spp., Anabasis salsa, Halocneum strobilaceum and by species from genus Artemisia. This type of vegetation is used as fodder reserves for the autumn-winter period, due to low yields and highly variable productivity, both between years and within seasons (Gintzburger et al., 2003; Kapustina, 2001). In the depressions, due to the shallow ground water level, develop solonchak or solonetz soils. On these soil types establishes halophytic vegetation, which is highly heterogenous and dynamic, due to diversified and unstable habitat characteristics (constant changes in water level). Among the species growing in saline depressions are salt tolerant species from genus Artemisia and succulent halophytes from the families Chenopodiaceae and Amaranthaceae, including many species from genera Salicornia, Halostachys, Kalidium, Halocneum, Salsola, Halimocnemis and Anabasis. In waterlogged places establish various graminoids, for example, Aeluropus litoralis, Pharagmites australis, Erianthus spp. (Gintzburger et al., 2003; Kapustina, 2001). Halophyte vegetation is characterized by low productivity and differentiated palatability. Nevertheless, it is used by camels during autumn and winter, when the excess salts are leached away by precipitation (Gintzburger et al., 2003).
Archaeological site and field research
Ayakagytma ‘The Site’ (N 40°39′05″, E 64°37′’06″) is located in the south-eastern part of the Kyzyl-kum desert, on the territory of the Republic of Uzbekistan, in Bukhara district, some 160 km North of the city of Bukhara, and about 60 km North of the town of Shafrikan (Figure 1).
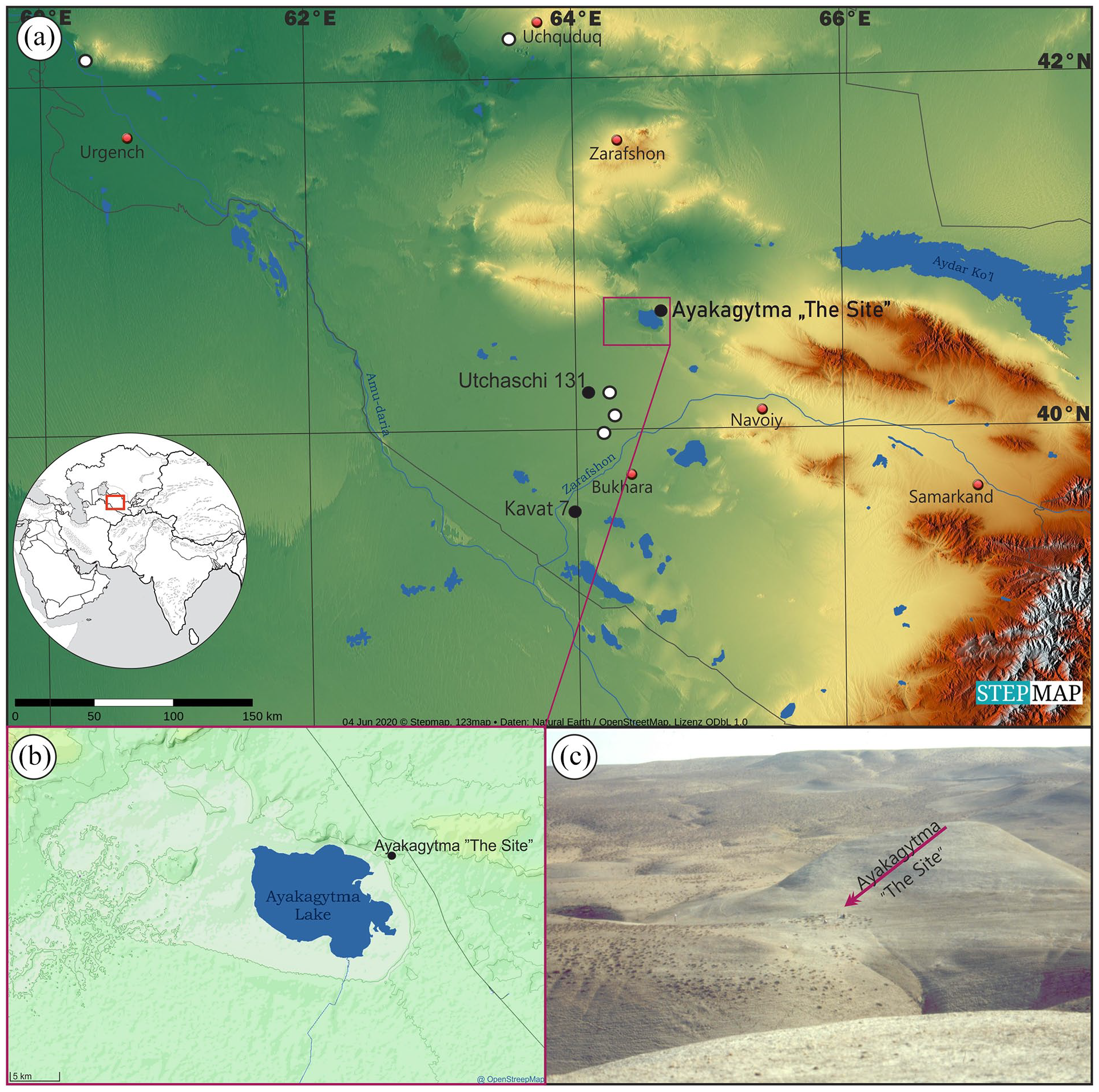
(a) The most important Keltaminar sites in the Kyzul-kum desert. Black dots - stratified sites, white dots - clusters of surface collections. (b) Location of Ayakagytma “The Site”. (c) Landscape of the Ayakagytma “The Site”.
‘The Site’ is situated on a plateau on the East side of a gorge, 350 m from an actual edge of the Ayakagytma Depression – a solonchak endorheic depression located between sandy-loam/clay formations and large areas of sand dunes. ‘The Site’ is separated from the depression not only by a gorge but also by a hill chain. The plateau itself is limited from the West by the main gorge, from the North and the South – by supplementary gorges, while from the East it is limited by a limestone island hill of some 15 m of relative height. At the feet of this hill, surface archaeological material occurred in an extremely dense concentration, on an area of some 1500 m2. Thus, the main trench was located there, in the area of the highest surface artefacts concentration (see Supplemental Figure S1, available online). In total, 124 m2 were excavated, which is probably much less than 10% of the whole area of ‘The Site’. The finds from every square metre were documented separately, after dry sieving of all the material through 4 mm mesh. The same was done with the following mechanical layer, about 10 cm thick. Excavations brought a large collection of various archaeological artefacts (7793 artefacts in total), including flints, stone, pottery and worked and unworked animal bones. All these finds had their position documented and were attributed to a particular stratigraphic/planographic unit (Szymczak et al., 2006). 14C analysis of a series of 13 charcoal samples found on site was performed by β-counting (using conventional method employing CO2 gas proportional counters) at the Laboratoire des Sciences du Climat et de l’Environment (Unite Mixte de Recherche CEA -CNRS) at Gif-sur-Yvette, and the Modane Underground Laboratory (French Alps) (see Supplemental Table S1 and Figure S2, available online). The zooarchaeological analysis was conducted by the Institute of Archaeology, University of Warsaw. The results were presented in Szymczak et al. (2006). Finally, seven main stratigraphic units were differentiated on a combined, general cross-section revealed along the East-West axis of Ayakagytma ‘The Site’; the numbers used in Supplemental Figure S1 correspond with the numbers which mark units described in detail in Szymczak et al. (2006).
Samples for lipid analyses
In the presented study, we performed chemical fingerprinting of interpretable amounts of lipids (i.e. >5 μg/g d.w. according to Evershed, 2008) extracted from potsherds from the site. Detailed analyses of fatty acid composition and of 13C isotopic ratios for C16:0 and C18:0 fatty acids were performed on artefacts from the Tuskan phase of the local Neolithic (4000–3000 cal BC), namely 19 potsherds found scattered on-site and 4 samples of organic residues acquired from a vessel found inside a clay oven, to our knowledge the oldest complete pottery vessel found in Central Asia. The samples of organic residues were taken from smoked parts inside this vessel and from an organic wood tar substance that was probably superimposed on the outside lower part of the vessel for sealing and insulation. These samples were used for comparative measures to exclude potsherds that underwent a similar sealing procedure, which may have influenced their lipid composition and isotopic ratios. Additional analyses were performed on a sediment sample from a horizon dividing the older and the younger phases, and on a modern camel and horse milk obtained from animals living in the nearby Karakalpakstan region (mixed from four animals each).
Extraction and derivatization of FAMEs
Lipid analysis and interpretations followed protocols established in (Sachse et al., 2004) and (Patalano et al., 2020). All reagents and solvents used were of analytical grade (HPLC, >95% of purity). Between 7 and 11 g of cleaned and ground potsherd samples were extracted with an accelerated solvent extractor ASE 350 (DionexTM) at 100°C and 1500 psi for 15 min in three cycles, using a mixture of dichloromethane (DCM) and methanol (MeOH) (9:1, v:v). The obtained 35 mL of the total lipid extract (TLE) were evaporated under nitrogen stream, weighted and re-dissolved in 1 mL n-hexane. TLE was separated into fractions of different polarities on a Pasteur pipette conditioned at 500oC and filled with high-purity silica gel (60 Å pore, Sigma Aldrich) activated for 8 h at 120°C (Patalano et al., 2020). n-Hexane fraction was obtained as the first, DCM fraction as the second and MeOH fraction as the third. The MeOH fraction, containing fatty acids, was transmethylated with a mixture of 0.3 mL of toluene with acetyl chloride/MeOH (95:5, v:v) at 60°C for 12 h. After cooling to room temperature, methyl esters of fatty acids (FAMEs) were extracted with a solution of 1.5 mL of organic free 5% NaCl and 0.5 mL of n-hexane. After phase separation, n-hexane layer containing FAMEs was transferred with a pipette into a new vial.
GC-MS analysis
To assess FAMEs composition of an individual sample 1 µL of n-hexane extract was injected on the Agilent 7890B gas chromatograph equipped with a capillary column HP-5 ms Ultra Inert (30 m × 0.25 mm × 0.25 μm) and coupled with the 5977A MSD mass spectrometer. The carrying gas was He with a constant flow of 2 mL/min. The following temperature programme was used: 50°C for 1 min, heating to 210°C at a rate of 20 °C/min for 18 min, heating to 230°C at a rate of 20 °C/min, and hold for 13 min. The compounds in the studied samples were identified by comparing their retention times with those of the reference compounds from the Supelco 37 Components FAME mix CRM 47,885 standard. A chromatogram for this standard is presented in Figure 2a, showing separation and resolution of peaks. Retention times of the reference compounds from the standard are given in Table 1. For each compound from the Supelco 37 Components FAME mix CRM 47885 standard a mass spectrum was generated and verified against the NIST Mass Spectral Library (an example of C16:0 mass spectrum is presented on Figure 2b). Obtained mass spectra were used in verification of compounds identified in the studied samples.
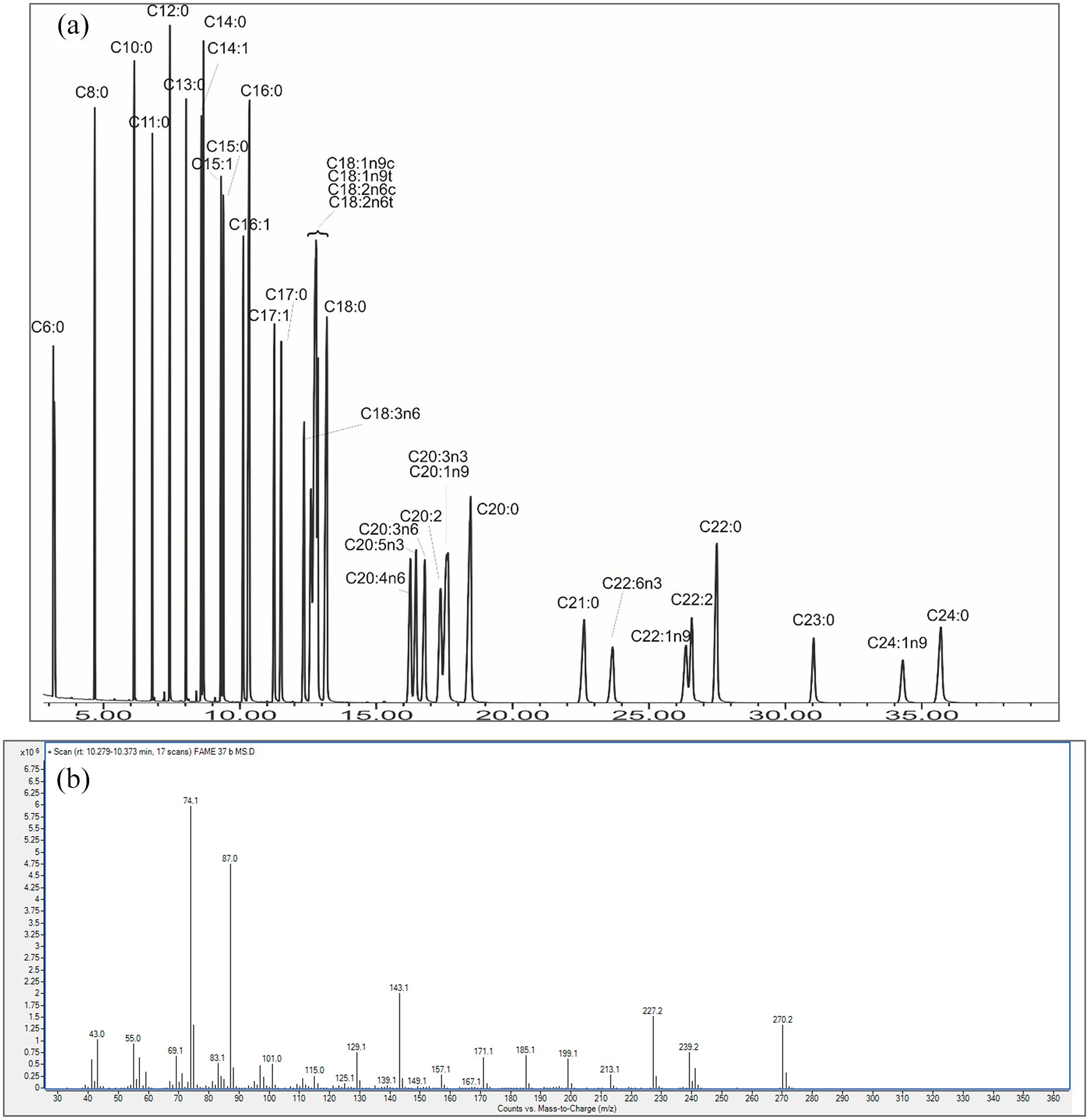
(a) – Chromatogram of the Supelco 37 Components FAME mix CRM 47885 standard, showing separation and resolution of peaks. Retention times are given in Table 1. (b) – An exemplary mass spectra of C16:0 from the Supelco 37 Components FAME mix CRM 47885 standard.
Retention times for FAMEs from the Supelco 37 Components FAME mix CRM 47885 calculated as means out of five runs of the standard.
Results of the performed analyses are presented as relative FA concentrations, calculated as a ratio of each FA area to the sum of all FAME areas, and multiplied by 100, as in (Johansson et al., 2016). Sample chromatograms of the studied samples are shown in Supplemental Figure S3, available online.
GC-IRMS analysis
To provide better identification of potential lipid sources in archaeological artefacts, δ13C isotopic signatures of palmitic (C16:0) and stearic (C18:0) fatty acids in n-hexane extracts were measured with the GC IsoLink + Trace GC Ultra (Thermo) gas chromatograph connected online with the IRMS Delta Plus (Thermo) mass spectrometer in the Continuous Flow system (measurement accuracy ±0.33 ‰). FAMEs were separated on an Rtx5-MS chromatography column and burned in a reactor at 1030°C. The CO2 obtained from combustion was introduced directly by a capillary into the spectrometer source. The isotopic ratios are expressed using the delta (δ) value as follows: δ13C [(13C/12C)sample/(13C/12C)standard-1] × 1000‰. Measurements were calibrated according to a calibration curve using standard F8-3 mix and A7 mix (Schimmelmann, Indiana University). Isotopic ratios were calculated with the use of the internationally accepted standard (V-PDB). A correction for methyl group introduced into each fatty acid during derivatization was performed according to the equation presented, for example, in Regert (2011).
All analyses were performed at the Laboratory of Biogeochemistry and Environmental Conservation at the Biological and Chemical Research Centre of the University of Warsaw. Complete results of lipid and isotopic analyses are presented in the Supplemental Dataset S1, available online.
Results
Fatty acid composition in the studied samples
Among the archaeological samples (i.e. potsherds, organic residue samples and a sediment sample), the highest total lipid content (TLE) was recorded in samples of organic residue from the vessel (between 5.9 and 3.7 mg/g d.w.). The average TLE in the potsherds was 2.5 mg/g d.w. the lowest TLE was observed in the sediment sample (1.1 mg/g d.w.).
In all the studied archaeological samples C16:0 and C18:0 were the dominating fatty acids. C16:0 constituted 28.8% of all the analysed FAs in the sediment, 36.6% on average in potsherds and 52.2% on average in the organic residue samples; and C18:0 comprised 10.0%, 18.2% and 36.1%, respectively (Figure 3). The third most abundant FA present in all the archaeological samples was C14:0, constituting 11.0% of all the analysed FAs in the sediment, 13.8% on average in potsherds and 2.5% on average in the organic residue samples. In comparison with the potsherds and the organic residue samples, the studied sediment was characterized by lack of long chain fatty acids (C20:0 – C24:0) and of most of the analysed unsaturated fatty acids, with the exception of C16:1, C17:1 and C22:1. The last FA was present exclusively in the sediment sample. Short chain fatty acids C4:0 and C6:0 were present only in single potsherd samples. Both the potsherds and the organic residue samples were characterized by the presence of C11:0 (0.2% and 0.3% on average, respectively) and by relatively high percentage participation of the mix of unsaturated C18 FAs (on average 12.3% in the potsherds and 3.7% in organic residue samples), that were absent in the sediment sample.
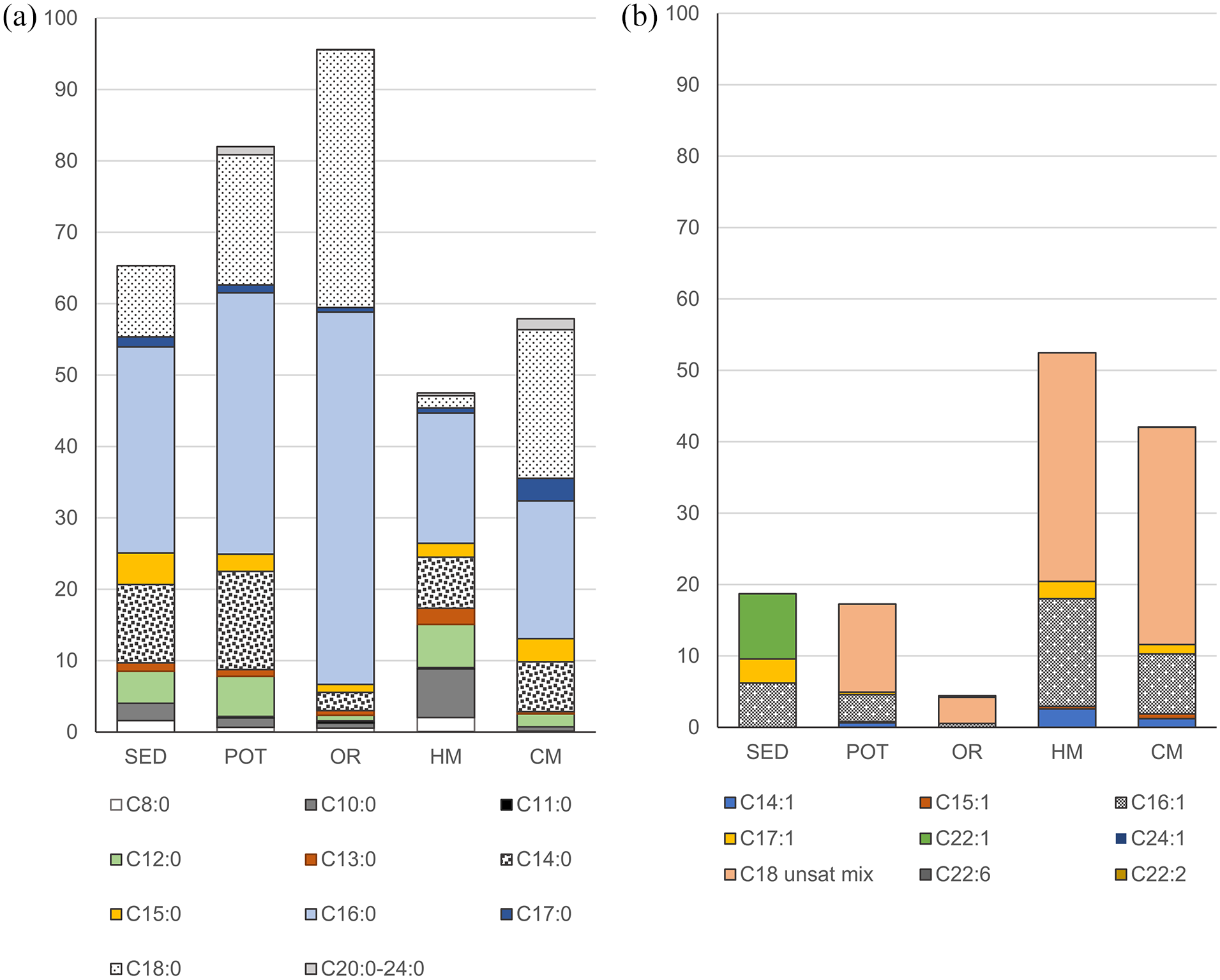
Relative content [%] of the analysed fatty acids in the samples of sediment (SED), pottsherds (POT), organic residue (OR), modern horse milk (HM) and modern camel milk (CM). Values given for the potsherds and the organic residue samples are averages out of 19 and 4 samples, respectively. (a) Saturated fatty acids, (b) unsaturated fatty acids. Sum of both saturated and unsaturated fatty acids constitutes 100%. Detailed values for all the individual samples are provided in the supplementary materials as Dataset S1.
Modern camel milk contained three times more TLE than modern horse milk (1268 mg/g d.w. in comparison to 400 mg/g d.w.). In both milk samples dominated mix of unsaturated C18 FAs (30% in camel milk and 32.1% in horse milk) and C16:0 FA (19.3% in camel milk and 18.2 in horse milk). The third most abundant FA in both milks was C16:1, comprising 8.4% in camel milk and 15.1% in horse milk. In comparison to horse milk, camel milk was characterized by 20 times higher participation of C18:0 FA (20.8% all the analysed FAs vs 1.7%) and four times higher participation of C17:0 FA (3.14% vs 0.7%). However, the relative content of C10:0, C12:0, C13:0 FAs was lower in camel milk than in horse milk, namely 0.02% versus 0.14%; 1.8% versus 6.1% and 0.3% versus 2.3%, respectively.
Fatty acid isotopic ratios in the studied samples
In the case of δ13C isotopic ratio for C16:0 and C18:0 FAs, the potsherd samples covered a wide range from −33.64‰ to −22.00‰ for δ13C 16:0 and from −37.16‰ to −24.35‰ for δ13C 18:0, and thus were dispersed over a scatter plot in Figure 4a. Eight potsherd samples were located near the modern horse milk sample (−26.14‰ for δ13C 16:0 and −28.25‰ for δ13C 18:0), in the range of δ13C 18:0 values established by Outram et al. (2009) for ancient and modern horse milk from Kazakhstan. For the remaining potsherd samples, δ13C 16:0 and δ13C 18:0 values were higher and modern camel milk sample was located inside this group (−23.74‰ for δ13C 16:0 and −24.81‰ for δ13C 18:0). A potsherd sample with the lowest δ13C 16:0 and δ13C 18:0 values, formed a separate cluster with a sediment sample (−33.34‰ for δ13C 16:0 and −36.86‰ for δ13C 18:0) and one of the organic residue samples (−33.01‰ for δ13C 16:0 and −37.06‰ for δ13C 18:0). The remaining organic residue samples formed a separate cluster in between the values of −30‰ and −32‰ for both δ13C 16:0 and δ13C 18:0.
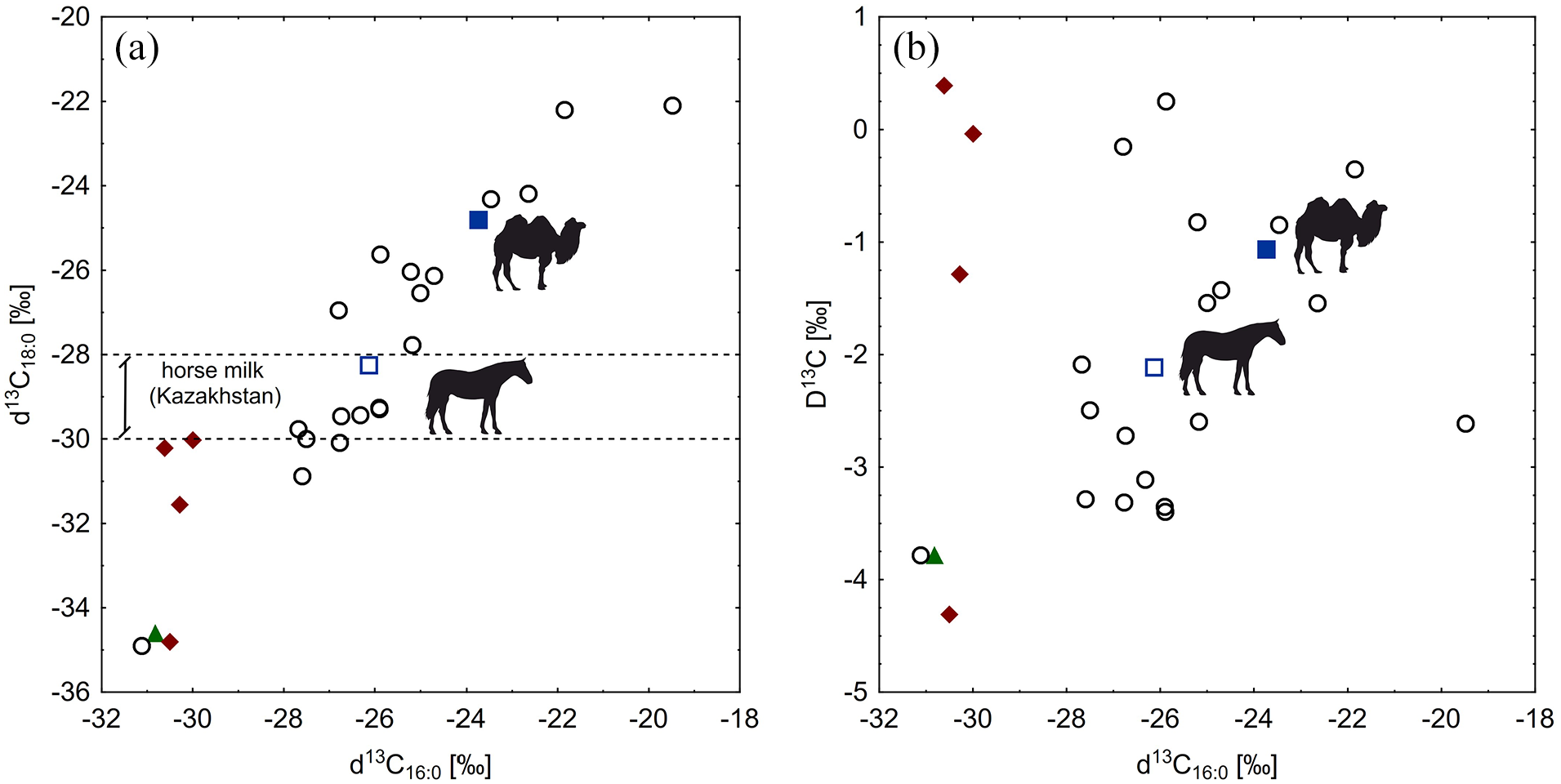
(a) Plot of δ13C16:0 values against δ13C18:0 values. Range of δ13C 18:0 for horse milk from Kazakhstan (ancient and modern) were given after Outram et al. (2009) (dotted lines). (b) Plot of Δ13C values against δ13C16:0 values. Circles – potsherd samples, diamonds - organic residue samples, triangle – a sediment sample, full square – a camel milk sample, empty square – a horse milk sample.
A scatter plot in Figure 4b. showing δ13C 16:0 isotopic ratios against Δ13C values complements Figure 3a. Apart from the sediment sample, one organic residue sample and one potsherd sample, all others were characterized by Δ13C values above −3.3‰. The remaining three organic residue samples formed a cluster with Δ13C values between −1.1‰ and 0.5‰. The best part of the potsherd samples grouped either in the lower-central part of the plot, together with modern horse milk sample (Δ13C value −2.1‰), or in the upper-centre, together with modern camel milk (Δ13C value −1.1‰).
Discussion
Identification of ancient fatty acid sources: a preliminary assessment
Over the last 20 years of studies on lipid residues preserved in archaeological potsherds and vessels, several molecular and isotopic criteria were proposed to distinguish potential sources of animal fats. Though the isotopic reference values for C16:0 and C18:0 fatty acids were determined for modern animals raised in Britain on a diet based on C3 plants (e.g. Evershed, 2008; Konuspayeva et al., 2008), available data from other studied geographical regions, while not identical, are mostly comparable (see Supplemental Table S2, available online). Nevertheless, data obtained from outside of the European region should be interpreted cautiously, as samples from distant locations (different isoscapes, different food sources) may significantly differ chemically (e.g. Bocherens et al., 2006; Roffet-Salque et al., 2017). In modern camel and horse milk samples a huge variance in FAs composition and isotopic ratios was found, depending on geographical location and type of fodder (Barnard et al., 2007; Copley et al., 2003; Doreau and Martuzzi, 2006), stage of lactation and milk production intensity (Barnard et al., 2007; Doreau and Martuzzi, 2006; Zorya et al., 2019), and, in the case of camel, also state of hydration of an individual animal (Barnard et al., 2007). Therefore, we decided to use as an additional reference material modern camel and horse milk sampled in the Karakalpakstan region under climatic conditions similar to these of the Kyzyl-Kum desert, where Ayakagytma ‘The Site’ is located.
The observed characteristics of modern camel and horse milk sampled in the Karakalpakstan region, such as the domination of C16:0 and mix of unsaturated C18 FAs in both milks; significantly higher TLE content and higher relative C18:0 content in camel milk in comparison to horse milk, are in accordance with the available literature (Barnard et al., 2007; Doreau and Martuzzi, 2006). Both sampled milks showed a combination of features typical for fats produced by the Northern-European ruminant and non-ruminant animals (see Regert, 2011) – that is, occurrence of mixed isomers of C18:2 FA and low amount of straight carbon chains with odd carbon number (both typical for ruminant fats), and domination of C16:0 FA over C18:0 FA (typical for porcine fats). In the case of the camel milk, our results of C16:0/C18:0 and C16:1/C18:1 ratios were in good accordance with results given for modern camel milk from Kazakhstan (Konuspayeva et al., 2008). Considering the isotopic values, a δ13C16:0 value of the modern horse milk fell in the range typical for Northern Europe modern reference material of non-ruminant fats. Contrastingly, the modern camel milk was characterized by a higher δ13C16:0 value, that fell in the range established for the North-African archaeological samples of ruminant dairy fats, and was close to δ13C16:0 values in ruminant fats resulting from total subsistence on C4 plants (between −22‰ and −12‰) (Dunne et al., 2012; Evershed, 2008; Gregg et al., 2009; Grillo et al., 2020; Outram et al., 2011; Regert, 2011) (see Supplemental Table S2, available online). As C16:0 FA in milk originates mainly from de novo synthesis of acetate from carbohydrates in the mammary gland, its isotopic composition is directly linked to animal diet (Regert, 2011). Since today Karakalpakstan is among the poorest regions of Uzbekistan, with the Gross Agriculture Output well below the national level (Rudenko and Dosov, 2015), the access to fodder is limited in this region (Rudenko and Dosov, 2015; Zorya et al., 2019). Therefore, both horses and camels still graze mostly in uncultivated rangelands covered with a mix of shrubs and small shrubs, graminoids and flowering non-woody plants (including ephemeral and ephemeroid species as in Gintzburger et al., 2003), mainly following a C3 photosynthetic pathway. C4 plants are represented there by halophytic species developed on saline soils (solonchaks and solonetz) in deflated areas between sand dunes and sandy-loam/clay formations. Such areas cover approximately 3% of the Uzbekistan area (Gintzburger et al., 2003; Kapustina, 2001). Thus, C4 species occur in comparatively lower, yet locally significant numbers. They belong mostly to the Chenopodiaceae and Amaranthaceae families (e.g. Agriophyllum spp., Ceratocarpus spp., Climacopthera spp., Salsola spp.) (Gintzburger et al., 2003). Due to limited water availability, both C3 and C4 plants in the southern part of Central Asia are significantly enriched in 13C isotope, leading to δ13C values ranging from −26.3‰ to −23.6‰ for C3 plants, and from −16.4‰ to −11.3‰ for C4 plants (Bocherens et al., 2006; Ventresca Miller et al., 2020). Thus, higher δ13C16:0 values of the modern camel milk may reflect the natural isotopic characteristics of rangeland vegetation, typical for warm and arid regions of Central Asia (Bocherens et al., 2006; Roffet-Salque et al., 2017; Ventresca Miller et al., 2020). The observed differences in δ13C16:0 value between the camel milk and the horse milk, probably result from dietary differences between these two animals, as camels are able to survive on typical dryland vegetation, including halophytes, most of which are C4 plants (Bocherens et al., 2006). The observed δ13C18:0 value of the horse milk was in accordance with the range established for non-ruminant fats in modern samples from Near East (Gregg et al., 2009) and with values described for the modern horse milk from Kazakhstan (Outram et al., 2009). The observed δ13C18:0 value of the camel milk, just as its δ13C16:0 value, fell into the range established for the North-African archaeological samples of ruminant dairy fats (Dunne et al., 2012). Both the recorded Δ13C values fitted into the range typical for the Northern European modern ruminant adipose fats (Copley et al., 2003; Dunne et al., 2012; Evershed, 2008; Gregg et al., 2009; Outram et al., 2011; Regert, 2011) (see Supplemental Table S2, available online).
Considering isotopic characteristics of our archaeological samples, potsherd profiles, apart from one sample, were distinct from the sediment sample and the organic residue samples. Thus, the probability that the potsherds underwent a sealing process and/or were contaminated with sediment matrix was relatively low. In case of δ13C16:0, 11 out of 19 potsherd samples showed values similar to the Northern-European ruminant and/or non-ruminant reference materials (between −31.0‰ and −25.0‰ according to Copley et al., 2003; Evershed, 2008). For the remaining eight samples (including four samples closest to modern camel milk from the Karakalpakstan region), values of δ13C16:0 fitted into broad ranges between −24.5‰ and −11.0‰ established for ruminant and non-ruminant fats in the North African archaeological samples by Dunne et al. (2012). In case of δ13C18:0, half of our samples showed values similar to modern horse milk from Kazakhstan (according to Outram et al., 2009), and the other half placed roughly in the range given by Evershed (2008) for the Northern-European non-ruminant fats (between −26.5‰ and −24.0‰). Regarding Δ13C values, most of the samples fitted into the range given by Evershed (2008) and Copley et al. (2003) for the Northern-European ruminant adipose fats. The samples that positioned close to modern camel milk from the Karakalpakstan region showed Δ13C values above −1.0‰ Δ13C, which according to Evershed (2008) and Copley et al. (2003) are typical for the Northern-European non-ruminant fats. Interestingly, fatty acid profiles of the potsherd samples that showed isotopic characteristics similar to modern camel milk, simultaneously displayed high relative content of C18:0 (including two the highest values recorded for potsherds − 57.0% and 38.8%) and C17:0 FAs, combined with very low values of C10:0, C12:0 and C13:0 relative content (including the lowest values of these FAs recorded for the potsherds). These characteristics reflect the features typical for camel milk. Thus, though no conclusive evidence for the presence of camel milk in the potsherds was obtained, the results of our study provide some valuable clues considering the process of camel domestication.
Ayakagytma findings in the context of camel domestication process
Hitherto, the earliest evidence for the camel domestication dated to the second half of the fourth millennium and the first half of the third millennium BC came from the Chalcolithic and Early Bronze Age sites of Anau and Geoksyur-5 (Chong-depe) in Southern Turkmenistan, as well as from Shahr-i Sokhta in Sistan and Sialk III located in East-central Iran (Bökönyi and Bartosiewicz, 2000; Compagnoni and Tosi, 1978; Duerst, 1908; Harris, 2010; Kuzmina, 2015; Salvatori and Tosi, 2005; Figure 5).
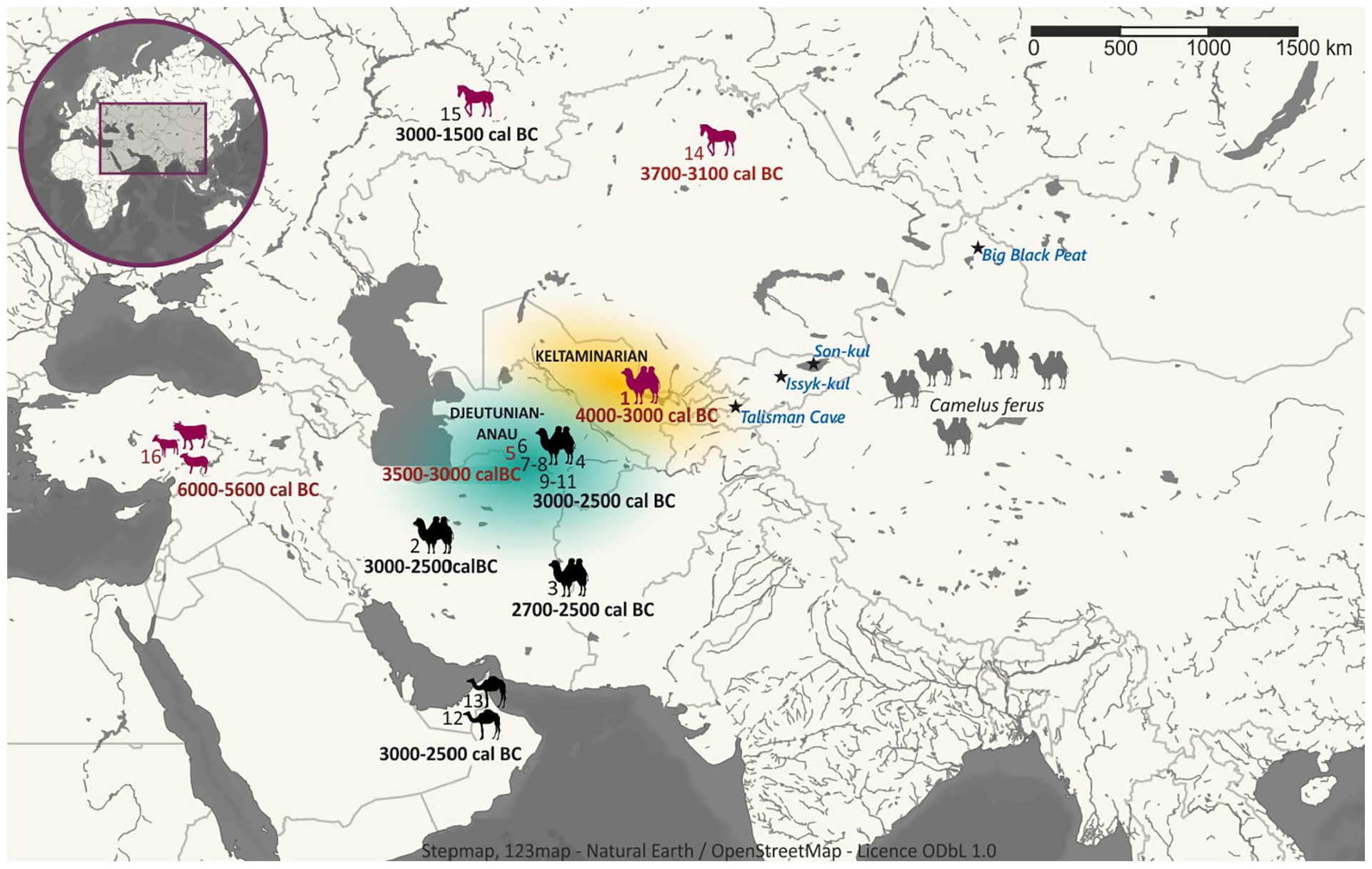
A map presenting the earliest traces of camel domestication and dairy products use.
Dark pink silhouettes represent traces of the earliest dairy product use; grey camel silhouettes represent the modern range of wild camels (Camelus ferus); black camel silhouettes represent archaeological sites with the oldest domesticated camel remains. Sites marked in red are dated to the Neolithic and Chalcolithic periods, whereas sites marked in black are dated to the Early Bronze Age. 1. Ayakagytma ‘The Site’, 2. Sialk III, 3. Shahr-i Sokhta (Bökönyi and Bartosiewicz, 2000,, Salvatori and Tosi, 2005), 4. Gonur Depe, 5. Anau (Duerst, 1908), 6. Shor-depe, 7. Namazga-depe, 8. Ulug-depe, 9. Altyn-depe, 10. Hapuz-depe, 11. Geoksyur-5 (Chong-depe) (Kuzmina, 2015), 12. Umm an-Nar (Hoch, 1979), 13. Ras Ghanada (Uerpmann and Uerpman, 2012), 14. Botai (Wilkin et al., 2020), 15. Bulanovo (Warinner et al., 2014), 16. Çatalhöyük West (Hendy et al., 2018). Extension of sites of nomadic Keltaminarian culture is marked in orange, whereas the Djeitunian and later Anau culture attributed to advanced farming is marked in green. Stars indicate location of paleoenvironmental records compared in Figure 5.
The above mentioned assemblages contained camel bones determined as belonging to domesticated animals, though it must be stressed that the earliest camel remains from Anau were studied over 100 years ago (Duerst, 1908). Additionally, in the Sialk III site a camel was depicted on the pottery vessel (Sala, 2017). Evidence from many sites in Turkmenistan, including terracotta models of wheeled carts drawn by Bactrian camels found at the Altyn-depe site in contexts dating to the Namazga IV period (Kohl, 1992), and golden and silver vessels with representations of Bactrian camels found in situ at the Gonur-depe site (Sarianidi, 2005), established the possible time of camel domestication in the first half of the third millennium BC. Though they cannot be viewed as hard evidence, the preliminary results obtained by us at Ayakagytma ‘The Site’ may be used to additionally support localization of the centre of Bactrian camel domestication in the southern part of Central Asia. Especially, if combined with analysis of zooarchaeological assemblages provided by Lasota-Moskalewska et al. (2009). The authors of this analysis identified 1151 out of 7092 specimen of animal remains collected at Ayakagytma ‘The Site’ to species and/or genus, and compared quantitative structure of zooarchaeological assemblages between the early Dariasai phase (ca. 6100–5400 cal BC) and the middle Tuskan phase (ca. 4000–3000 cal BC) (see Supplemental Figure S4, available online). During both phases the dominating species were cattle (~30% during the Dariasai phase and ~6% during the Tuskan phase), horses (~34% during the Dariasai phase and 6% during the Tuskan phase, potentially including also wild-ass) and camels (~23% during the Dariasai phase and ~85% during the Tuskan phase). Based on well-marked traces of domestication on the collected bones, Lasota-Moskalewska et al. (2009) described cattle as domesticated animals, alongside sheep/goats, pigs and dogs, whose remains were represented in lower numbers in the zooarchaeological assemblages. Though no clearly visible traces of domestication were found on horse and camel remains, Lasota-Moskalewska et al. (2009) described them as animals that were potentially tamed and used, as the percentage participation of hunted animals (i.e. gazelles, aurochs/bison, deer and wild sheep) in the zooarchaeological assemblages was noticeably lower (all the identified hunted species in total constituted ~7% during the Dariasai phase and ~2% during the Tuskan phase). Complete lack of traces of advanced farming practices or permanent dwelling objects at the Ayakagytma ‘The Site’ confirms that the Keltaminar people remained a nomadic population, whose lifestyle and economy was based on breeding dairy animals (cattle and, during the Tuskan phase, possibly camels). As such it is an example that the evolution of animal domestication strategies does not necessarily have to be dependent on the adoption of conventional agriculture. If our further research provides conclusive information on the presence of camel milk in the potsherds from the Ayakagytma ‘The Site’, they will be one of the oldest known evidence of Bactrian camel domestication.
Changes in animal husbandry practices as adaptations to Holocene climate evolution
A comparison between faunal assemblages from the early Dariasai phase (ca. 6100–5400 cal BC) and from the middle Tuskan phase (ca. 4000–3000 cal BC) (see Supplemental Figure S4, available online) provides some clues about local climatic conditions over these periods. Domination of cattle and horses during the early phase, combined with lack of permanent dwelling structures and almost complete lack of pig remains, suggest that Keltaminar tribes were nomads, using relatively rich grazing sites (Lasota-Moskalewska and Szymczak, 2012; Lasota-Moskalewska et al., 2009). Scarce palynological data for this area indicate that before 5000 cal BC vegetation around Ayakagytma ‘The Site’ comprised small trees and shrubs, mainly from genera Tamarix and Haloxylon, perennial species from the Chenopodiaceae and Amaranthaceae families, graminoids and legumes (Khujanazarov et al., 2014). After the hiatus between 5400 and 4000 cal BC, stockbreeding practices of the Keltaminar tribes changed radically to almost exclusive domination of camels as a stock component. This shift can be interpreted as a result of a significant environmental change in the area, leading to a drawdown of the groundwater table and, consequently, a shift from mainly grassland vegetation to domination of significantly less nutritious desert shrubs. As a result, Keltaminar tribes were forced to the breeding of camels that can adapt to extreme environmental conditions and feed on such vegetation (Lasota-Moskalewska and Szymczak, 2012). Thus, we expect noticeably drier climate conditions during the middle Tuskan phase (4000–3000 cal BC) than during the early Dariasai phase (6100–5400 cal BC).
Available data on Holocene moisture variations over Central Asia are miscellaneous and sometimes contradictory. Therefore, three different models of Holocene climate changes were proposed for this region: (1) a trend of increasing moisture throughout Holocene (e.g. Chen et al., 2016; Kang et al., 2020; Wang et al., 2013), (2) maximum moisture in middle Holocene (approximately 6000–2000 cal BC), with dry early Holocene and moderately humid late-Holocene (e.g. Boomer et al., 2000; Chen et al., 2008; Lan et al., 2021) and (3) a trend of decreasing moisture – from humid early Holocene, through moderately humid middle Holocene to dry late-Holocene (e.g. Cheng et al., 2012; Lan et al., 2021). The pattern of changes observed at the Ayakagytma ‘The Site’ do not fit precisely into any of these models. However, some paleoenvironmental records from Westerlies-dominated arid Central Asia (as defined by Chen et al., 2008) give clear indications of a drought around 4000–3000 cal BC. Examples of such records, preserved in different environmental matrices and assessed with different proxies, are given in Figure 6.
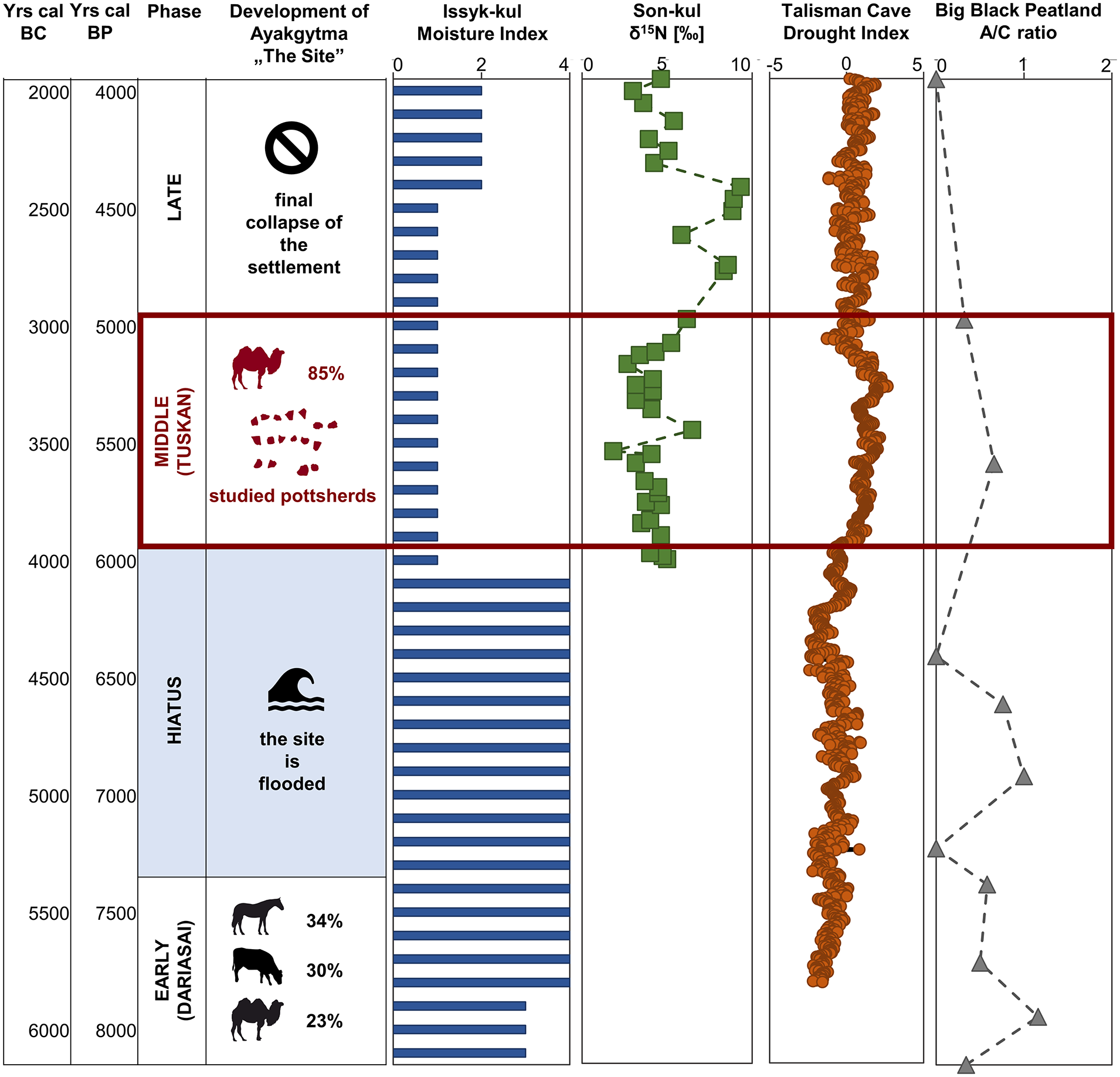
History of Ayakagytma ‘The Site’ in between 6100 and 2000 cal BC compared to the selected records of paleoclimate changes from the area of the Westerlies-dominated arid Central Asia as defined by Chen et al. (2008). Moisture Index for Issuk-kul Lake given after Chen et al. (2008); δ15N [‰] given after Lauterbach et al. (2014); Drought Index for Talisman Cave given after Tan et al. (2021); Artemisia to Chenopodiaceae (A/C) ratio for Big Black Peatland given after Xu et al. (2019). Data on the archaeozoological assemblages discovered in the older and in the younger horizon of Ayakagytma ‘The Site’ given Lasota-Moskalewska et al. (2009). Remains marked with the horse silhouette comprise Equidae family, represented in this study by horse and wild-ass. Percentages were calculated on the collection of 1151 specimen identified to species and/or genus.
Proxies’ fluctuations are shown only for the period of functioning of Keltaminar culture in the Ayakagytma ‘The Site’ (6100–2000 cal BC). In case of Issyk-kul Lake (Kyrgyzstan) at around 4000 cal BC started a phase characterized by lower lake levels and higher salinity combined with lower precipitation and higher evaporation (Chen et al., 2008; Ricketts et al., 2001). This phase is illustrated in Figure 6 by low values of the moisture index, calculated by Chen et al. (2008) on the basis of primary data on δ18O and CaCO3 given in previous publications on this lake (Ferronskii et al., 2003; Jiang et al., 2007; Rasmussen et al., 2001; Ricketts et al., 2001). On the scale used by Chen et al. (2008) (0) indicates the driest interval during Holocene at the given site and (4) – the wettest interval. The phase characterized by the lowest values of the moisture index lasted till approximately 2500 cal BC. Paleoenvironmental data from Son-kul Lake (Kyrgyzstan) comprise a large set of proxies, including isotopic data (δD, δ15N, δ13C and δ18O), salinity index (Sr/Ca), mineralogic data, algal remains and palynological data and show a complex picture of a lake changing its status from closed to open system (Huang et al., 2014; Lauterbach et al., 2014; Schwarz et al., 2017). During the period of 4000–3000 cal BC Son-kul Lake was closed and relatively saline. Moreover, values of δ15N of Son-Kul sediments were noticeably lower in comparison to the following periods, indicating decreased meltwater-derived input of soil material, and thus lower snowfall intensity in Central Kyrgyzstan (Lauterbach et al., 2014) (Figure 5). Interestingly, dry and cold winters during 4000–3000 cal BC were combined with relatively humid summers (according to δD values), resulting in high seasonality of precipitation (Huang et al., 2014; Schwarz et al., 2017).
Stalagmites collected in the Talisman Cave (Kyrgyzstan) provide one of the most precisely dated and highest-resolved mid-Holocene records from Central Asia (Tan et al., 2021). According to the Drought Index showed in Figure 6 and represented by the PCA results of δ18O, δ13C and Sr/Ca records in stalagmite samples, precipitation in Central Asia was relatively high before 4000 cal BC. However, abnormally positive values of the Drought Index, exceeding one standard deviation, were recorded between 3820 and 3180 cal BC, with the highest values at approximately 3280 cal BC. These data indicate a prolonged period of aridity, or even megadrought, lasting over 600 years (Tan et al., 2021). Finally, paleoenvironmental records from Big Black Peatland (China), including peat accumulation rate, palynological data and peat cellulose δ13C, indicate that the period between 4300 cal BC and 1800 cal BC was noticeably drier then both earlier and later stages in peatland development (Xu et al., 2019). Data presented in Figure 6 show that after approximately 4500 cal BC ratio of Artemisia to Chenopodiaceae pollen in peat decreased considerably, indicating a shift to vegetation typical to dry environmental conditions. Mid-Holocene drought signals were observed also in paleoenvironmental records from several other locations in Central Asia e.g. Lake Balkhash in Kazakhstan (Endo et al., 2012), loess sequences in Central Kazakhstan (Ran and Feng, 2014), peat cores from southern Altai Mountains (Zhang et al., 2020) and Dali Lake in China (Liu et al., 2015). Thus, this pattern of climatic changes, with relatively humid early and late-Holocene and dry middle Holocene, was recently listed as the fourth model of moisture variations over Central Asia (Lan et al., 2021).
Precipitation patterns over arid Central Asia depend on the intensity and the location of the westerly zone. The stronger and the more southward oriented Westerlies, the more moisture is transported from the North Atlantic, Mediterranean and Caspian Sea to arid Central Asia. The Westerlies in turn are affected by solar activity – the stronger it is, the higher surface temperature and more northward oriented westerly zone, resulting in lower supply of water vapour to Central Asia (Liu et al., 2015; Tan et al., 2021; Xu et al., 2019). According to Steinhilber et al. (2009), between 4000 cal BC and 3000 cal BC total solar irradiance showed some abrupt changes, that may have affected the westerly zone. Furthermore, Tan et al. (2021) described increased storminess in the North Atlantic during this period that may have resulted in a northward shift of the westerly zone (akin to a present-day positive North Atlantic Oscillation), and thus in reduced water vapour supply to Central Asia. Potential increase in surface temperature could have also enhanced evaporation in this region, which may have also partly contributed to the observed mid-Holocene drought (Tan et al., 2021; Xu et al., 2019). However, global climatic changes can be potentially modified by local conditions, including hydrological factors, and lead to complex global responses (e.g. Wolff et al., 2017). Hence, the observed inconsistencies of paleoenvironmental data. In the context of the Ayakagytma ‘The Site’ these inconsistencies can be illustrated with the records of closely located Aral Lake, which during the period of 4000–3000 cal BC was characterized by the highest water level in history and underwent a transgression phase (Boomer et al., 2000; Ferronskii et al., 2003).
Conclusions
Detailed analyses of composition and 13C isotopic ratios of fatty acids performed on 19 potsherds obtained from the multilayered Neolithic site located in the Ayakagytma ‘The Site’ in Uzbekistan provided preliminary insights into milking practices of nomadic stockbreeders from the Keltaminar culture. Though no conclusive evidence for the presence of camel milk in the potsherds was found, the results of our study, combined with the existing zooarchaeological data and the available paleoenvironmental records from the studied area, provide valuable clues indicating that the possibility that camels were kept and milked by the Keltaminar population during the period of 4000–3000 cal BC (Tuskan Phase) is plausible. Such results potentially support localization of the centre of Bactrian camel domestication in the southern part of Central Asia. If our further research provides conclusive information on the presence of camel milk in the potsherds from the Ayakagytma ‘The Site’, they will be one of the oldest known evidence of Bactrian camel domestication.
As compared to the older Dariasai Phase (6100 and 5400 cal BC), zooarchaeological assemblages changed radically in the Tuskan Phase, suggesting a shift towards almost exclusive domination of camels as a stock component. Such transformation seems to be an adaptation to a drier climate and poorer vegetation, and is in accordance with drought signals preserved in different matrices, that is, lake sediments, loess sequences and stalagmites, in several locations in the Westerlies-dominated Central Asia roughly in between 4000 cal BC and 3000 cal BC.
Supplemental Material
sj-docx-1-hol-10.1177_09596836221114289 – Supplemental material for Potential impact of Holocene climate changes on camel breeding practices of Neolithic pastoralists in the Central Asian drylands: A preliminary assessment
Supplemental material, sj-docx-1-hol-10.1177_09596836221114289 for Potential impact of Holocene climate changes on camel breeding practices of Neolithic pastoralists in the Central Asian drylands: A preliminary assessment by Małgorzata Suska-Malawska, Małgorzata Kot, Anna Gręzak, Monika Mętrak, Mukhiddin Khudjanazarov and Karol Szymczak in The Holocene
Research Data
sj-xlsx-2-hol-10.1177_09596836221114289 – Supplemental material for Potential impact of Holocene climate changes on camel breeding practices of Neolithic pastoralists in the Central Asian drylands: A preliminary assessment
Supplemental material, sj-xlsx-2-hol-10.1177_09596836221114289 for Potential impact of Holocene climate changes on camel breeding practices of Neolithic pastoralists in the Central Asian drylands: A preliminary assessment by Małgorzata Suska-Malawska, Małgorzata Kot, Anna Gręzak, Monika Mętrak, Mukhiddin Khudjanazarov and Karol Szymczak in The Holocene
Footnotes
Acknowledgements
Authors wish to express their gratitude to all participants of the Ayakagytma expedition who took part in the archaeological fieldworks; to the current and former Directors of the Institute of Archaeological Research, Uzbek Academy of Science for their constant support during 25-years of scientific cooperation; to Monika Chmielewska, Agata Trojan and Marcin Sulwiński Ph.D. for their invaluable help in laboratory work; and to Magdalena Malawska for assistance in designing Figures 1 and ![]() .
.
Data availability
All data used in this article is presented in its original form as a Supplemental dataset S1, available online.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This study was supported by the Polish National Science Centre (Grant No 2017/25/B/HS3/00520). The field and laboratory research in Ayakagytma ‘The Site’ was financed by the Polish State Committee for Scientific Research (Grant Nos 1HO1G01112 – Gr/795 and 2HO1H03622 – Gr/1681). Lipid analyses were financed by the statutory research of the Faculty of Biology, University of Warsaw (501-D114-01-1140900).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.