
Abstract
The question of the origin of Alpine farming and pastoral activities associated with seasonal vertical transhumance and dairy production in the Silvretta Alps (Eastern Switzerland) has recently benefitted from renewed interest. There, pastoral practises began during the Late Neolithic (2300 BC), but alpine dairy farming was directly evidenced so far only since the Late Bronze Age/Early Iron Age (1300–500 BC). The vegetation development, timberline shifts at 2280 m a.s.l. and environmental conditions of the subalpine Urschai Valley (Canton of Grisons, Switzerland) were reconstructed for the small (8 m2) Plan da Mattun fen based on palynological and geochemical analyses for the last six millennia. The X-ray fluorescence (XRF) analyses are among the first ones performed on a European peatland in such altitudes. A high Rb/Sr ratio in the fen peat sediments revealed an increase in catchment erosion during the time when the forests of the Upper Urschai Valley were steadily diminished probably by fire and livestock impact (2300–1700 BC). These landscape openings were paralleled by increasing micro-charcoal influx values, suggesting that prehistoric people actively set fire on purpose. Simultaneously, palynological evidence for pastoralism was revealed, such as pollen from typical herbs indicating livestock trampling, and abundant spores from coprophilous fungi. Since then, vertical transhumance and pastoral activities remained responsible for the open subalpine landscape above 2000 m a.s.l., most probably also in the context of milk and dairy production since 1300 BC, which is characteristic for the European Alps until today.
Keywords
Introduction
Past land-use practices have shaped today’s vegetation and landscape of the European Alps (Fischer et al., 2008). Since the mid-Holocene, human activities affected the position of the timber- and treeline, for example, through fires or pastoralism (Dietre et al., 2017, 2014; Staffler et al., 2011). In the Swiss Alps, vertical transhumance of livestock (sheep, goat and cattle) might have occurred since the Neolithic 7000 years ago (Hafner and Schwörer, 2018; Reitmaier et al., 2018). One of the purposes of leading herds to subalpine and Alpine meadows above 2000 m a.s.l. during summer was to save low-altitude arable lands for crop production (Akeret and Jacomet, 1997; Brombacher et al., 2007; Jacomet et al., 1999) and later for hay making (probably since the Iron Age), and also to produce storable and durable dairy products for supply during winter, and for trade. This characteristic land-use strategy implied prehistoric work force, livestock management and long absence from settlements in the valleys, which in turn required ways to treat and/or to store the milk or dairy products obtained during seasonal stays in the Alpine highlands. Lipid residues within ceramics (pottery) excavated in Alpine archaeological sites above 2000 m a.s.l. in the Silvretta Alps (Eastern Switzerland) revealed the start of dairy production to date back to at least the Late Bronze Age/Early Iron Age, c. 1300–500 BC (Carrer et al., 2016). Here, the oldest semi-permanent habitation structures attributed to pastoral activities were concomitantly also dated to these periods (Reitmaier, 2017). However, palaeoecological evidence suggests that pastoral activities above 2000 m a.s.l. may have started earlier (4250 BC; Dietre et al., 2014), but no corresponding habitation sites were identified so far from this period and altitude. Thus, those early pastoral activities had a different economic background (animal primary products from small ruminants). To further address the question of the onset of high-mountain pastoralism in the Eastern European Alps, we investigated its impact on the past vegetation and landscape of the subalpine Upper Urschai Valley (Silvretta Alps, Canton of Grisons, Switzerland). Based on palynological quantifications of microfossils from the well-dated Plan da Mattun fen peat sequence (2280 m a.s.l.), we reconstructed the vegetation history of the Upper Urschai Valley over the last 5900 years. We paid particular attention to spores from coprophilous fungi as local markers of past pastoralism, that is, prehistoric livestock presence (Cugny et al., 2010), as well as to micro-charcoal particles to investigate the role of fire in the opening of the formerly densely forested landscape. Furthermore, we considered palaeoecological and archaeological data together with geochemical proxies (X-ray fluorescence (XRF) analyses) from a parallel fen peat sequence in order to detect changes in the environmental settings in the fen’s catchment (and particularly erosion processes; Gadd et al., 2015) possibly linked to past livestock and anthropogenic activities by Late Neolithic herders. We discuss the potential synchronicity of these changes with general Central European climatic change. In addition, we discuss why fundamental changes in pastoral sustainability did probably happen since the Bronze Age (2200–800 BC), whereas the periods before were based – according to archaeological and archaeozoological evidences – on ephemeral campsites (such as abris or rock shelters or open air sites) with pasturing of small ruminants (sheep and goat) primarily for meat production, and eventually for wool (Kothieringer et al., 2015; Reitmaier, 2017).
Materials and methods
Study area
The Silvretta Alps connect Switzerland with Austria (Figure 1). Geologically, it is a crystalline area over the Penninic Bündner schists and is mostly composed of metamorphic mica-schists, gneiss, quartzite and marble, and with sedimentary rocks consisting of carbonates such as chalk, shale and dolomites (Frei et al., 1995; Gürler and Schmutz, 1995). The high valleys of the Silvretta Alps were formed during the Quaternary (the last 2.5 Mio years), and especially during the last Würmian glaciation (Felber, 2011). From the southeastern to northwestern oriented Lower Engadine Valley (Switzerland), the Tasna Valley stretches northwards near the village of Ardez and splits thereafter at 2000 m a.s.l. westwards into the Urezzas Valley, and northeastwards into our research area, the Urschai Valley (Figure 1). The Urschai Valley ranges from 2050 to 2400 m a.s.l. and features passes up to 2800 m a.s.l.
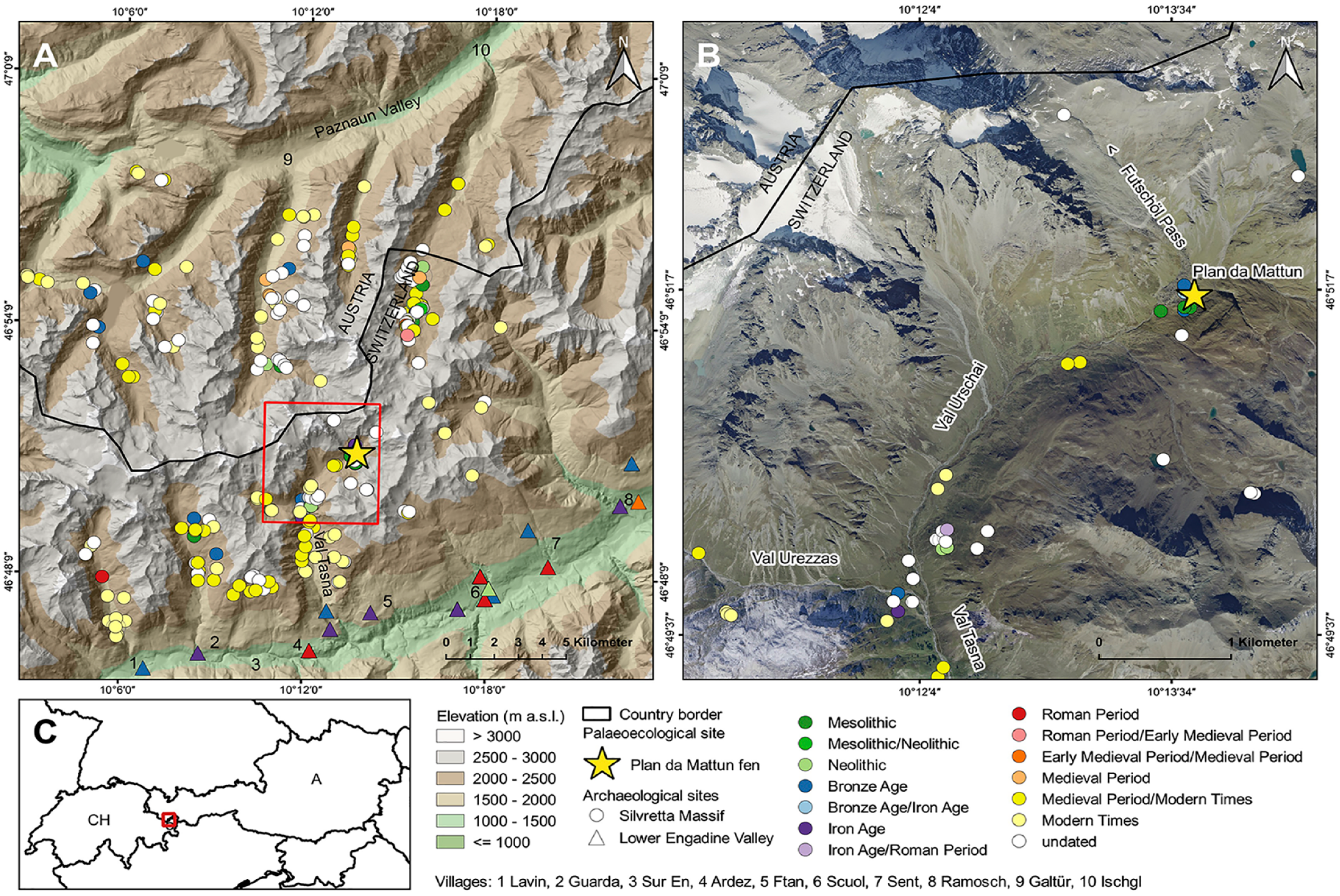
Location of the Plan da Mattun fen (yellow star; 2280 m a.s.l.) with known upland and lowland archaeological sites (from the Mesolithic to the Medieval period; effective 2017) in the Silvretta Alps and Lower Engadine Valley (Canton of Grisons, Switzerland). Source of the Digital Terrain Model: ASTER GDEM (ASTER GDEM is a product of METI and NASA); source of ortho-imagery: Federal Office of Topography swisstopo and Canton of Grisons; source of country-borders: http://diva-gis.org/gdata. Graphic by Christoph Walser, March 2018.
The current vegetation in the Upper Urschai Valley is mainly composed of alpine meadows, livestock pasture and a pioneer flora on rocky slopes. Some few dwarf shrubs are present up to 2600 m a.s.l., such as the dwarf willows Salix reticulata, S. retusa, Daphne striata (striated daphne) and Juniperus communis (mountain juniper). The area is treeless today, and the nearest trees (such as arolla pine – Pinus cembra, or European larch – Larix decidua) are growing up to an altitude of 2100 m a.s.l. in the Lower Urschai Valley. However, closed forests are found in the Tasna Valley up to 2100 m a.s.l. (with single trees up to 2200 m a.s.l). Nearby, in the Val Laver Valley, the closed forest, that is, timberline, stretches to 2200 m a.s.l., and the treeline can reach up to 2300 m a.s.l.
The Plan da Mattun site is located in the subalpine Upper Urschai Valley, at 2280 m a.s.l. (Figures 1 and 2). It is a small (8 m2), but 2-m-deep, fen fed by a small rivulet and underground waters and is located just outside a large Late Glacial rock fall complex dated to the Egesen stadial (10,700–9600 BC, that is, Younger Dryas) originating from below the nearby Piz Futschöl mountain peak (3174 m a.s.l.; Figure 2). These rock structures belong to the oldest subalpine sites in the European Alps used by prehistoric hunter-gatherers as abris (rock shelters) since 8600 BC (Early Holocene/Mesolithic; Cornelissen and Reitmaier, 2016), about 1000 years after the retreat of the local glaciers (Felber, 2011). There, at the Plan da Mattun rock shelter complex, flint tools dating to the Mesolithic and Neolithic periods, pottery fragments of the Laugen-Melaun-type – typical for the Inner Alpine Late Bronze and Early Iron Age (1300–500 BC) – and calcined bones from animals were excavated. The latter may represent a small alpine burnt offering place. Sites of this type generally emerged during the Late Bronze Age and were linked to alpine livestock farming (Reitmaier, 2017). Another archaeological site nearby, the Abri Urschai rock shelter (2180 m a.s.l.; Figure 1) at the entrance of the Urschai Valley, is also noteworthy. It contained several fire places left behind by Neolithic hunter-gatherer societies, as well as animal bones (only from wild animals), arrowheads and other flint tools from hunters, and pottery dating mainly to the late fifth (and up to the third) millennium BC. In addition, charred remains from collected and eatable plants, such as arolla pine seed shells, Cotoneaster seeds, as well as Galium cf. G. pumilum seeds (pastures bedstraw) – all growing locally today in the Urschai Valley – were found at Abri Urschai and point to the use of these plants for consumption or ethnobotanical purposes (Reidl, 2015). Charred oak acorn fruits (Quercus) were found as well, which suggest the use of staple food by these prehistoric societies, as oak trees were likely absent from our research area at that time (Reidl, 2015). Those acorns would have been gathered in the according oak distribution areas such as in Vorarlberg (Austria, 20 km northwest), or in the Vinschgau Valley (South Tyrol, Italy, 25 km southeast), and would then have been transported by people to the Urschai Valley as staple food. The overall composition of all these plant and animal remains and archaeological objects, and the topographical situation of the rock shelter argue for a seasonal camp of Neolithic hunter-gatherers of wild animal and plant resources. In any case, there are no clear local archaeological indications of a systematic seasonal use of high pastures in the Urschai Valley already in the Neolithic.

Photograph of the small (4 × 2 m2) Plan da Mattun fen (Urschai Valley, Canton of Grisons, Switzerland, 2280 m a.s.l.) with the coring location (yellow star) right besides the Mesolithic to Medieval archaeological complex of Plan da Mattun (between the large rock boulders on the left). View northwards towards the Futschöl Pass. Photograph by Jean Nicolas Haas, 2010.
Finally, a large livestock enclosure with a small dry stone shelter, fireplaces and prehistoric pottery excavated at the junction of Plan d’Agl between the Tasna, Urezzas and Urschai Valleys (2060 m a.s.l.; Figure 1) dates to the Late Bronze Age/Early Iron Age. It is recognised as one of the oldest pastoral infrastructures left behind by prehistoric shepherds in the area (Reitmaier, 2017). Analyses of several potsherds provided direct evidence of dairy farming for this alpine building (Carrer et al., 2016).
Field work
On 8 July 2010, a 195-cm-long sediment core (PMM-2010-C) was extracted from the Plan da Mattun fen using a Russian-type peat corer (Belokopytov and Beresnevich, 1955) with a 5-cm-wide and 50-cm-long coring chamber. The obtained stratigraphy revealed Cyperaceous (sedge) fen peat partly interrupted by minerogenic erosional features and wood particles. The sediment cores transported to the University of Innsbruck (Austria) were stored at 4°C before sampling and subsequent palynological preparations. On 2 August 2012, two parallel sediment cores were retrieved from the same Plan da Mattun fen (cores PMM-2A and PMM-2B) using a rod-operated Usinger system piston corer (Mingram et al., 2007). With a total length of 175 cm, PMM-2 comprises four overlapping segments of maximal 1 m in length. These sediment cores were transported to the University of Kiel (Germany) in densely packed plastic in-liners, where they were split lengthwise in the lab and stored in a core repository at 4°C for further use.
Chronology
Plant macrofossils from terrestrial plants were extracted by wet-sieving of fen peat samples from the PMM-2010-C core using a mesh size of >125 µm, and subsequently 10 AMS radiocarbon dates were obtained at the ETH Zürich (Switzerland) using the dedicated MICADAS system (Synal et al., 2007; Table 1). Potential contaminations with carbonates or humic acids were removed using an acid and base treatment as described by Hajdas (2008). The 14C ages were calculated according to Stuiver and Polach (1977) and transformed into calibrated years BC/AD according to the IntCal13 calibration curve (Reimer et al., 2013) with the package rbacon (Blaauw et al., 2007) within the software R (R Core Team, 2017; Figure 3a) in order to produce an age-depth model. The chronology of the PMM-2A/B core was obtained by stratigraphical matching to the PMM-2010-C core, using easily recognisable, fine, inorganic layers, and was supplemented by two additional AMS radiocarbon dates on wood particles obtained at the AMS facility in Poznan (Poland; Table 1). An age-depth model was produced by smooth spline interpolation of these dates using the software clam (Blaauw, 2010; Figure 3b) and a degree of smoothing set to 0.4.
AMS radiocarbon dates performed on macrofossils from terrestrial plants extracted from the fen peat stratigraphies of Plan da Mattun (Canton of Grisons, Switzerland, 2280 m a.s.l, cores PMM-2010-C and PMM-2A/B).
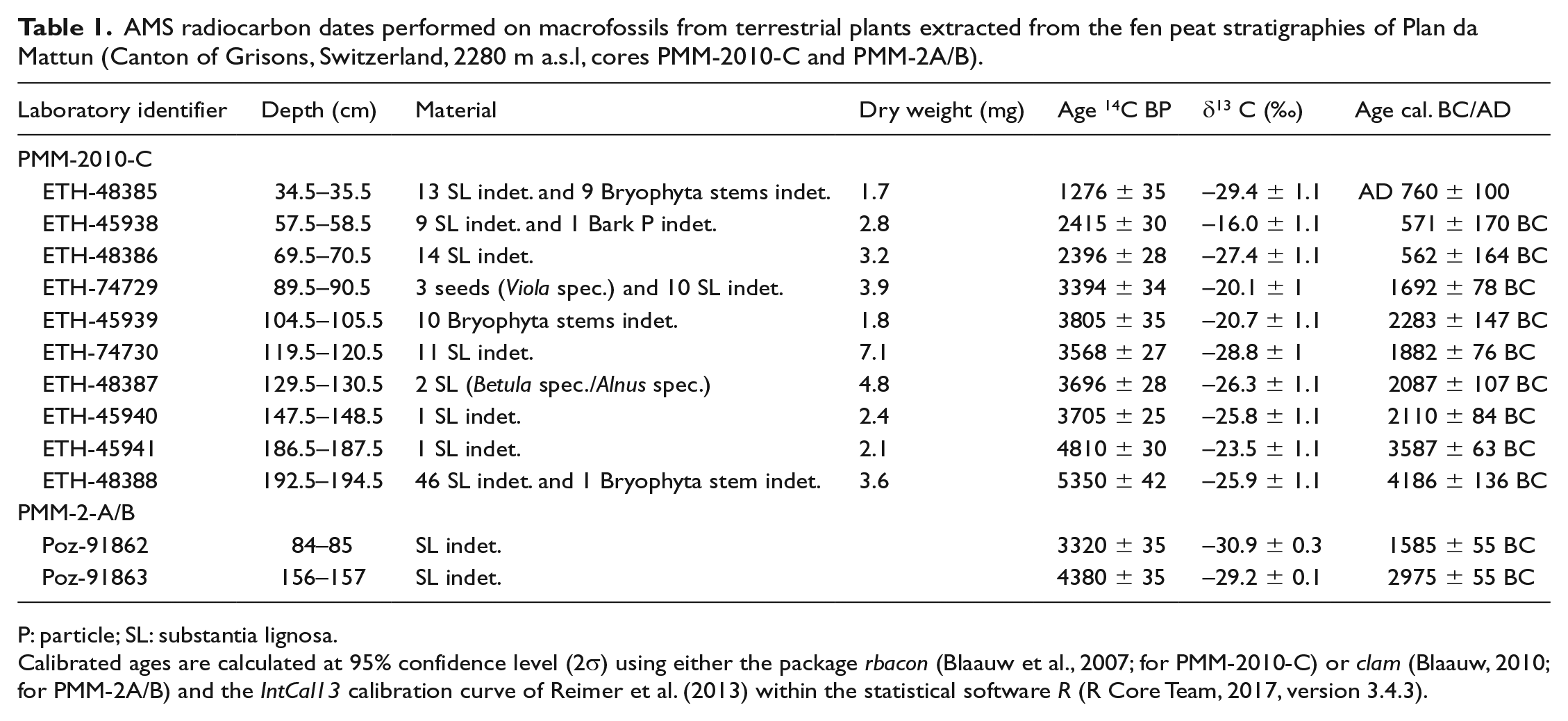
P: particle; SL: substantia lignosa.
Calibrated ages are calculated at 95% confidence level (2σ) using either the package rbacon (Blaauw et al., 2007; for PMM-2010-C) or clam (Blaauw, 2010; for PMM-2A/B) and the IntCal13 calibration curve of Reimer et al. (2013) within the statistical software R (R Core Team, 2017, version 3.4.3).
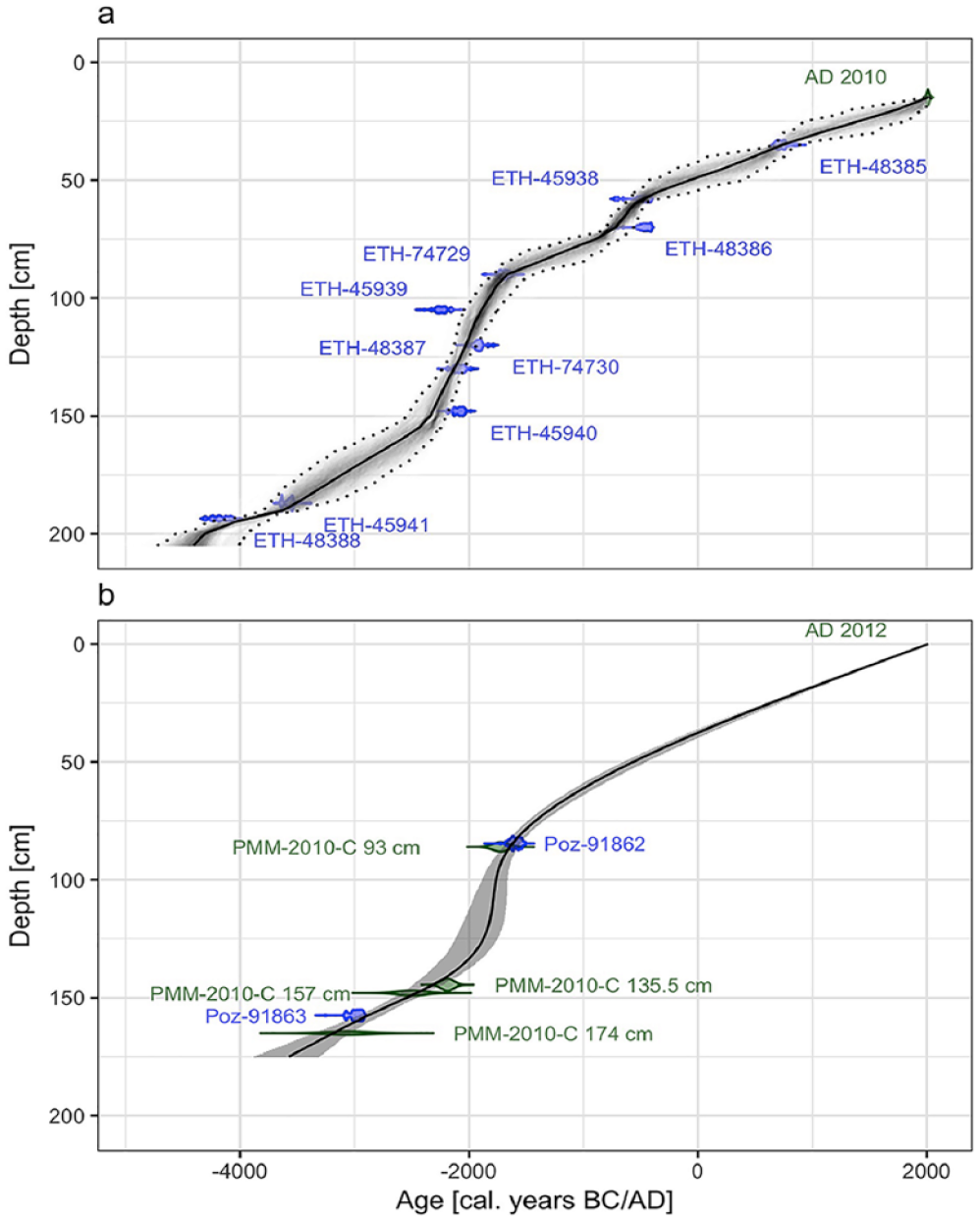
Age-depth models of the Plan da Mattun fen stratigraphies (Urschai Valley, Canton of Grisons, Switzerland, 2280 m a.s.l.). The age-depth model of the core PMM-2010-C (a) is based on the Bayesian interpolation of 10 AMS radiocarbon dates (see Table 1) and one surface date (AD 2010) with the R package rbacon (Blaauw et al., 2007). The age-depth model of the cores PMM-2A/B (b) is based on smooth spline interpolation of two AMS radiocarbon dates (see Table 1), the age of four stratigraphical layers matched with the core PMM-2010-C, and one surface date (AD 2012) with the R package clam (Blaauw, 2010). The solid lines represent the weighted mean modelled age retained; the dotted lines and the grey envelops represent the 95% confidence interval limits. Graphic by Benjamin Dietre, August 2019.
Palynological preparations and analyses
A total of 53 sediment samples of 1 cm3 volume were extracted from the 195-cm-long PMM-2010-C stratigraphy every 3–4 cm. A known number of Lycopodium clavatum (club moss) spores were added to each sample to allow calculation of concentrations and influx values (Maher, 1981; Stockmarr, 1971). Samples were first sieved using a 150-µm and a 7-µm mesh sieve, and the remaining 7–150 µm fraction was chemically treated following the procedure described by Seiwald (1980). This included a 1-min chlorification (1 mL of concentrated sodium chlorate and 1 mL of concentrated hydrochloric acid) at 95°C, a 1-min acetolysis at 95°C and a final staining in glycerine with fuchsine. An additional step with hydrofluoric acid (10%) was used for silty samples. Pollen and fern spores were identified using determination keys (Beug, 2004; Fægri and Iversen, 1989; Moe, 1974; Moore et al., 1991) and the palynological reference collection of the Institute of Botany at the University of Innsbruck, Austria. For each sample, all microfossils were identified and quantified until a minimum sum of 500 determined tree pollen was achieved. Pinus undiff. comprises pollen from Pinus cembra and Pinus sylvestris-type (Scots pine) which includes Pinus mugo (mountain pine). Cereal-type pollen (Cerealia) refers to Poaceae (grasses) pollen featuring the typical characteristics of cereal pollen (i.e. larger than 37 µm, annulus width and pore diameter each larger than 2.7 µm, and annulus thickness larger than 3 µm; Beug, 2004). The definition of pastoral pollen indicators follows Behre (1981), Oeggl (1994) and Festi (2012). Spores of coprophilous fungi and other non-pollen palynomorphs (NPPs) were identified according to van Geel and Andersen (1988), van Geel et al. (2003), van Geel and Aptroot (2006), Cugny et al. (2010), Gelorini et al. (2011) and Dietre et al. (2014). The most abundant and important coprophilous fungi (Cercophora, Podospora-type and Sporormiella) are presented in the ‘Results’ section, whereas other less important taxa – Arnium-type, Chaetomium, Delitschia, Neurospora (syn. Gelasinospora) – are included in the sum of coprophilous fungi. Black, opaque and sharp particles (7–150 µm) were identified as micro-charcoals and quantified on the same palynological slides. All palynological data are available on request and will be stored in the European Pollen Database (EPD) at Aix-en-Provence (France) in due time.
Geochemical analyses
One half of each of the four segments of the PMM-2A/B cores was scanned at 5-mm resolution for major elements with an Avaatech X-Ray Fluorescence (XRF) core scanner equipped with a rhodium tube (Richter et al., 2006). For low-mass elements (Al, Si, S, Cl, K, Ca, Ti, Mn, Fe), a measurement setting of 10 kV and 500 µA was used. For elements with higher mass (Br, Rb, Sr, Zr), 30 kV and 2000 µA was chosen. Exposition time was set to 20 s, and element counts per second (cps) were measured. The humification degree (HD) of the fen peat layers was determined photometrically (Zhong et al., 2010) using a UV 1800 Shimadzu spectrometer at 400 nm. Fen peat samples were dried, powdered and sieved with a mesh size of 63 µm. For each sample, 50 mg of powder was diluted in 50 mL of sodium hydroxide solution and then boiled at 80°C for 1 h. After cooling, the samples were centrifuged for 10 min at 3500 r/min, and 5 mL of each sample was pipetted into a new tube, filled up to 50 mL with distilled water, and measured. These relative values were corrected for the organic matter content determined by loss on ignition according to the formula
Illustration and statistical methods
The diagrams of pollen, cryptogam spores and NPPs were drawn using the software Tilia (Grimm, 2015). Pollen taxa were expressed as percentages of the 100%-pollen sum, which was defined by the total of terrestrial vascular plants, but excluding Cyperaceae and wetland/fen species. Other microfossils (such as spores from coprophilous fungi) were expressed as influx values (number cm–2 yr–1). Clusters of similar pollen assemblages (local pollen assemblage zones (LPAZs)) were assessed by a stratigraphically constrained incremental sum of square clustering based on the square-root transformed percentage values of terrestrial pollen (excluding unidentified pollen, Cyperaceae and wetland/fen species) using the package rioja (Juggins, 2012) in the software R (R Core Team, 2017). The relevant number of LPAZs (i.e. clusters) was ascertained using the broken stick model within the same package. Major shifts in vegetation were also reconstructed by a correspondence analysis (CA) using the package vegan (Oksanen et al., 2018).
Chemical element cps were expressed as a proportion of the total cps (Cuven et al., 2011), or converted as specific element ratios. Pearson’s correlation coefficients were calculated for pairs of the major geochemical proxies using the corrgram package (Wright, 2014).
Results
Palynological analyses
The clustering analysis of the 53 pollen assemblages from the Plan da Mattun fen identified two distinct clusters, the LPAZ 1 from 3900 to 1950 BC and the LPAZ 2 from 1950 BC to AD 1750, with three and four sub-zones, respectively (Figure 4). High arboreal pollen values (>65%) characterise the assemblages of LPAZ 1, dominated by pine (Pinus undiff.), spruce (Picea abies) and green alder (Alnus viridis, syn. Alnus alnobetula). However, herbs such as Poaceae and Cichorioideae (chicory subfamily within the Asteraceae – daisy family) are the dominant taxa (33–54%) in the pollen assemblages of LPAZ 2.
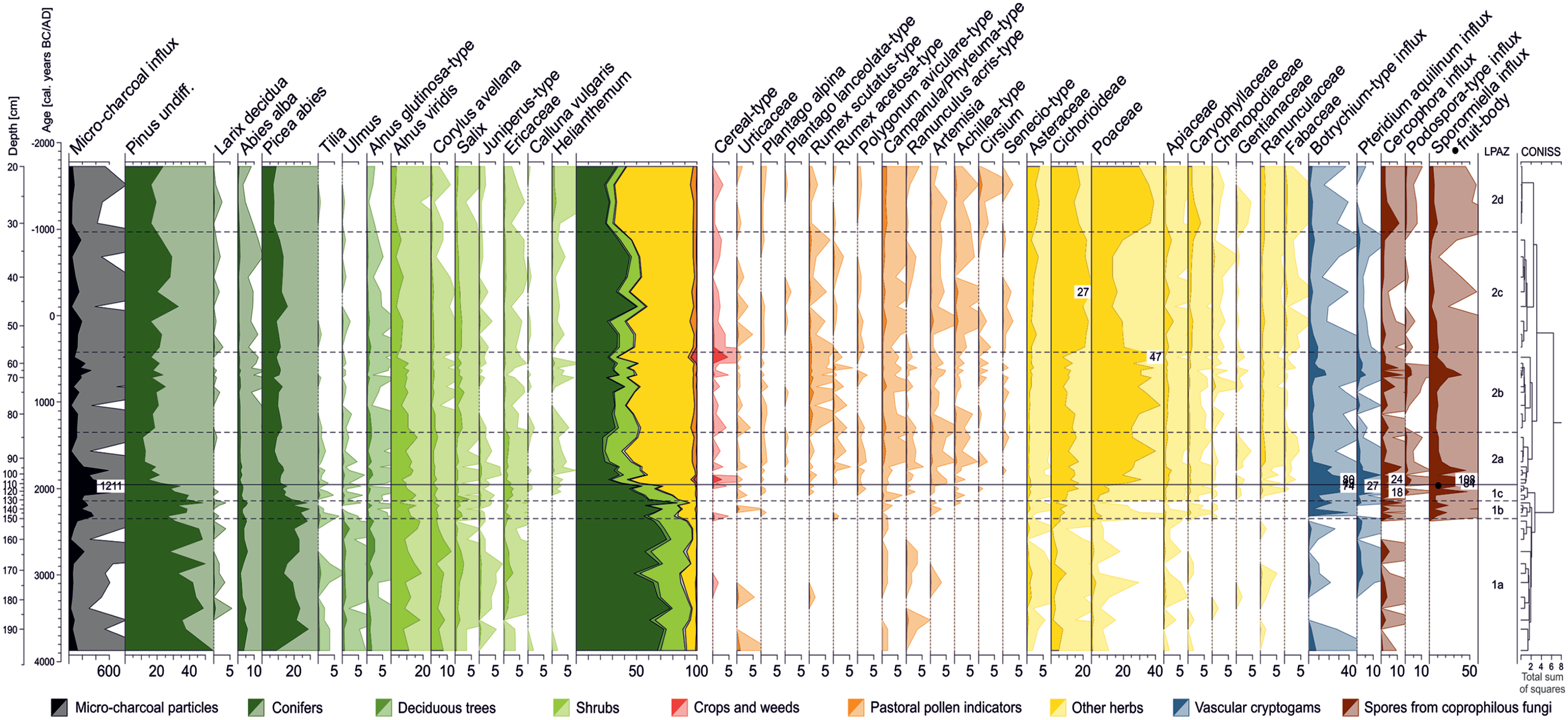
Palynological diagram for selected pollen taxa, spores from vascular cryptogams and coprophilous fungi, as well as micro-charcoal particles found in 53 peat samples from the Plan da Mattun fen (Urschai Valley, Canton of Grisons, Switzerland, core PMM-2010-C, 2280 m a.s.l.). Pollen taxa are expressed as percentage values of the 100%-pollen sum (excluding Cyperaceae and wetland/fen species). Other taxa and micro-charcoal particles are expressed as influx values (number cm−2 yr−1). Light-colour silhouettes represent a 10-fold exaggeration of the percentage values. Graphic by Benjamin Dietre, August 2019.
LPAZ 1a (3900–2350 BC; 193–150 cm)
The pollen record from the PMM-2010-C core starts at 3900 BC. Arboreal pollen dominate (Pinus undiff., Picea abies, Abies alba – fir, Larix decidua, Alnus viridis, Corylus avellana – hazel, Tilia – lime, and Ulmus – elm) and account on average for 92% of the pollen sum (Figure 4). A limited amount of herb taxa was recorded: Cichorioideae, Asteraceae, Poaceae and Ranunculus acris-type (common buttercup). A few pollen grains of Artemisia (sagewort), Campanula/Phyteuma-type (bellflower/rampion) and Apiaceae (umbellifers) were present, as well as cereal-type pollen (0.2%) at 3100 BC. Spores of Pteridium aquilinum (bracken fern) appeared around this time. Spores of the coprophilous fungus Cercophora were also present, as well as some fungal spores of Neurospora (data not shown), and a modest amount of micro-charcoal particles (83 particles cm–2 yr–1 on average).
LPAZ 1b (2350–2150 BC; 150–131 cm)
In this subzone (Figure 4), arboreal pollen values were slightly lower than those of LPAZ 1a (86% on average), while an increase in micro-charcoals was recorded (285 particles cm–2 yr–1 on average). Cichorioideae (up to 10%) and Poaceae (6.8%) also increased, and cereal-type pollen grains (0.5%) were found at 2300 BC. Some pollen from plants typical for pasture are present, such as Plantago alpina (alpine plantain), Rumex acetosa-type (sorrel), Artemisia, Achillea-type (yarrow), as well as fern spores of Botrychium-type (moonwort). In addition, pollen from the herb families of Caryophyllaceae (pinks) and Chenopodiaceae (chenopods) start to increase, together with high spore values of the coprophilous fungi Sporormiella and Cercophora.
LPAZ 1c (2150–1950 BC; 131–111 cm)
The last subzone of LPAZ 1 records a rapid decrease in arboreal pollen within 200 years with values falling from 89% (transition LPAZ 1b–1c) to 61% (transition LPAZ 1c–2a) mainly due to the reduction in Pinus undiff., Picea abies, Alnus glutinosa-type (European black alder) and Abies alba (Figure 4). Concomitantly, the number of micro-charcoal particles rapidly rose from 192 to 1211 particles cm–2 yr–1 (i.e. the highest amounts found in the whole stratigraphy). In parallel, only shrubs and subshrubs – Alnus viridis, Salix, Juniperus-type, Helianthemum (rockroses), Calluna vulgaris (common heather) and the Ericaceae in general (heath family) – benefitted from the reduction in the abundance of pollen of the above-mentioned subalpine trees. Apart from a massive rise in Cichorioideae (up to 17%) and Poaceae (up to 11%) and steady increases of Achillea-type, Campanula/Phyteuma-type, Apiaceae, Botrychium-type and Pteridium aquilinum, first pollen from other plants typical for pastoral areas appeared, namely Cirsium (thistle) and Polygonum aviculare-type (knotgrass). Spores from the coprophilous fungi Cercophora (up to 18 cm–2 yr–1) and Sporormiella (up to 47 cm–2 yr–1) reached their first maximum at 2050 BC, and spores of Podospora-type were found for the first time. A fruit-body of Sporormiella (containing about 20 spores) was also recorded around 1950 BC, as well as cereal-type pollen grains (0.4%).
LPAZ 2a (1950–1350 BC; 111–84 cm)
The LPAZ 2a introduces a 3700-year-long palynological record dominated by Cichorioideae and Poaceae. In this first subzone, they accounted on average for 14% and 27% of the total assemblages, respectively. Tree pollen amounts decreased to their lowermost values of 24% for the whole record, at 1550 BC. Pollen of pastoral plants showed higher values than during LPAZ 1, and some new types were recorded, namely Plantago lanceolata-type (ribwort plantain), Senecio-type (ragwort), Fabaceae (legume family) and Gentianaceae (gentians). At the beginning of LPAZ 2a, an important peak of cereal-type pollen was recorded (2.8%). At 1900 BC, amounts of spores from coprophilous fungi were maximal, including Cercophora (24 spores cm–2 yr–1), Sporormiella (108 spores cm–2 yr–1), Podospora-type and Chaetomium (each 3 spores cm–2 yr–1; the latter not shown). The amounts of micro-charcoals progressively decreased to 100 particles cm–2 yr–1, except for a high value at 1800 BC (586 particles cm–2 yr–1). The lowest value for Pinus undiff. (11%) was found at the end of this subzone (1400 BC).
LPAZ 2b (1350–400 BC; 84–56 cm)
During LPAZ 2b, herbs such as Cichorioideae and Poaceae remained on high levels (11% and 34% on average, respectively). Trees and shrubs levelled at 46% on average, but pollen from Pinus undiff. and Salix were slightly more frequent than previously (19% and 1.6%, respectively). Pollen from plants growing in livestock pastures were regularly present, and particularly those from Campanula/Phyteuma-type and of Rumex scutatus-type (buckler-leaved sorrel). The maximum amounts of spores of coprophilous fungi were found at 700 BC (40 Sporormiella spores cm–2 yr–1, 13 Cercophora spores cm–2 yr–1 and 4 Podospora-type spores cm–2 yr–1). This happened at the beginning of a short phase (750–500 BC) where micro-charcoal particles showed values higher than 100 particles cm–2 yr–1. Right after, a second peak of cereal-type pollen was registered at 500 BC (4.9%).
LPAZ 2c (400 BC–AD 950; 56–32 cm)
During this subzone, Poaceae pollen decreased down to 12%, giving space for Cichorioideae to become the dominant taxa (up to 27%). Arboreal pollen values showed a slight recovery towards 50%, with Pinus undiff. (24%) being the main contributor. Castanea sativa (chestnut) pollen appeared for the first time at 200 BC (data not shown) and accounted for up to 0.8% of the pollen assemblage at AD 700. Concomitantly, Achillea-type, Artemisia, Campanula/Phyteuma-type and Fabaceae were present in increasing amounts, whereas spores of coprophilous fungi (Cercophora, Podospora and Sporormiella) were less frequent.
LPAZ 2d (AD 950–1750; 32–20 cm)
The final subzone LPAZ 2d revealed the vegetation development of the last millennium. Arboreal pollen values decreased compared with the LPAZ 2c towards 34% on average, mainly due to the reduction in Pinus undiff., Picea abies and Abies alba pollen amounts. Inversely, Helianthemum reached maximum pollen values (1.3%), and Poaceae became again the most dominant taxa (as during LPAZ 2b) accounting for 30–39% of the pollen sum. Pollen of Campanula/Phyteuma-type were regularly present (1.5%) as well as of Cirsium, which reached its maximum value for the entire sequence (1.2% at AD 1500). Spores of coprophilous fungi were more frequent than during LPAZ 2c, specifically Cercophora and Sporormiella (up to 11 and 10 spores cm–2 yr–1 at AD 1050). Pollen of Asteraceae accounted for up to 4.1% at the very end of the record (AD 1750).
Major changes in pollen assemblages
We used a CA to detect the main changes in pollen data from the Plan da Mattun fen. The first axis of the ordination gathered 43.3% of the total inertia of the pollen assemblages. Combined with the second axis, they gathered 54.5% of the inertia. The first axis marks a clear distinction between two clusters: assemblages older than 1900 BC show site scores lower than −0.2 on this axis, and younger assemblages show greater site scores (Figure 5a). Assemblages from 2100 to 1950 BC (scores from −0.5 to −0.2) can be seen as a transition towards the next cluster of assemblages. These two clusters correspond well to the age delimitations of the LPAZ 1 and 2 based on a clustering of the same data as described above. A further distinction could be set around 400 BC based on the second axis of the ordination, corresponding to the limit between LPAZ 2b and 2c.
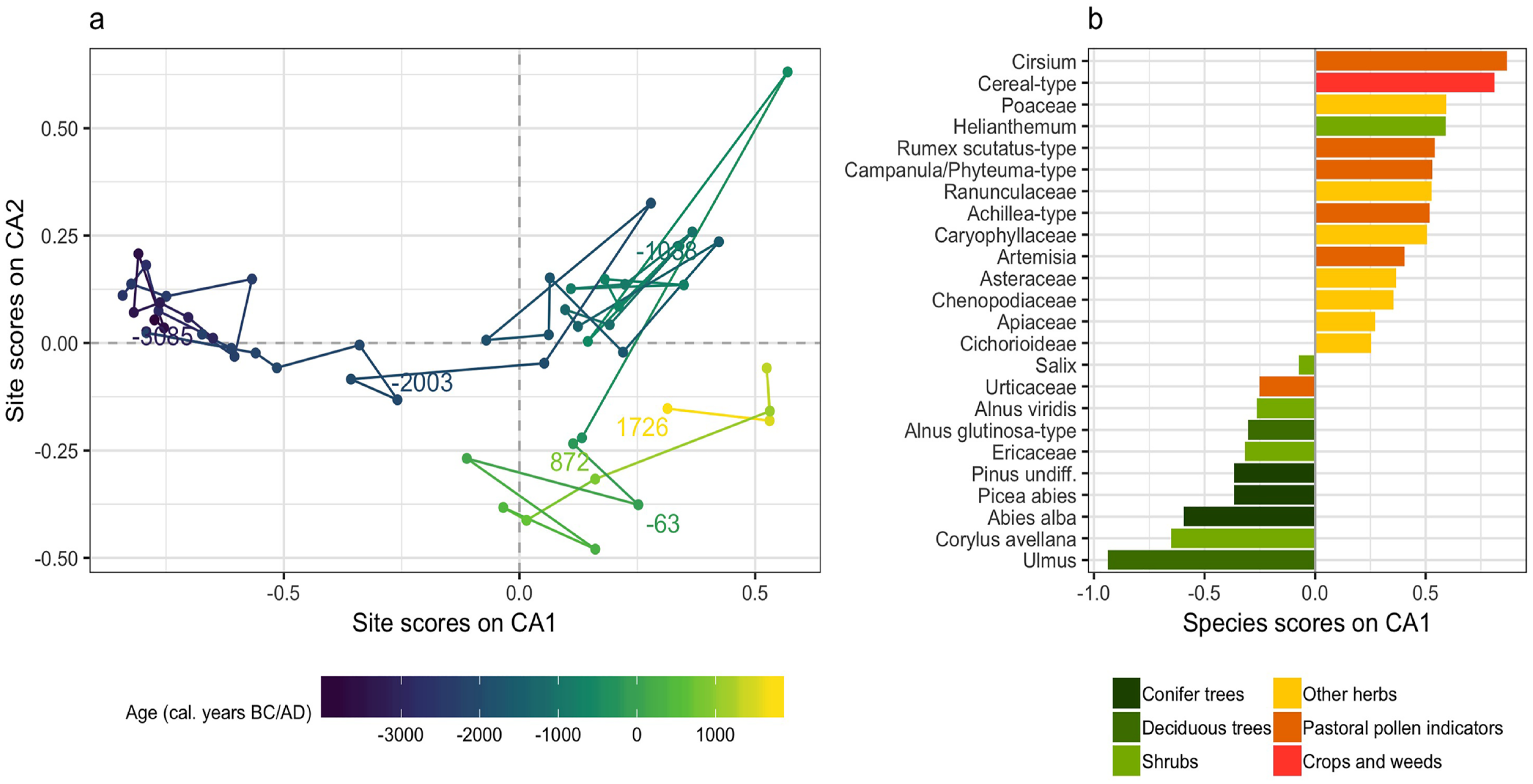
Site biplot (a) and species scores of taxa on the first axis (b) of a correspondence analysis made on pollen data from the Plan da Mattun fen (Urschai Valley, Canton of Grisons, Switzerland, core PMM-2010-C, 2280 m a.s.l.). The taxa presented accounted at least once in the sequence for 1% or more of the pollen sum. Graphic by Benjamin Dietre, August 2019.
To understand the vegetation shifts reflected in these major changes, one may look at the species scores of taxa on the first axis of the ordination (Figure 5b). Only taxa accounting at least once for 1% of the pollen sum are represented. Ages which are found with negative site scores are likely to exhibit important amounts of taxa that are found with negative species scores. Thus, assemblages older than 1950 BC contained larger amounts of pollen of Ulmus, Corylus avellana, conifers (Abies alba, Picea abies and Pinus undiff.), Ericaceae, Alnus glutinosa-type and A. viridis. However, younger assemblages contained larger quantities of pollen from taxa that are found with positive species scores, particularly Cirsium and cereal-type pollen, and, generally speaking, of herb pollen from crops, weeds, pastoral indicators and open land taxa.
Geochemical analyses
The oldest fen peat of the 175-cm-long core PMM-2A/B used for XRF scanning dates back to 3550 BC (Figure 3b). Some selected proxies (Figure 6) describe the palaeoenvironmental and palaeoclimatic development of the site and support the palaeoecological analyses described above.
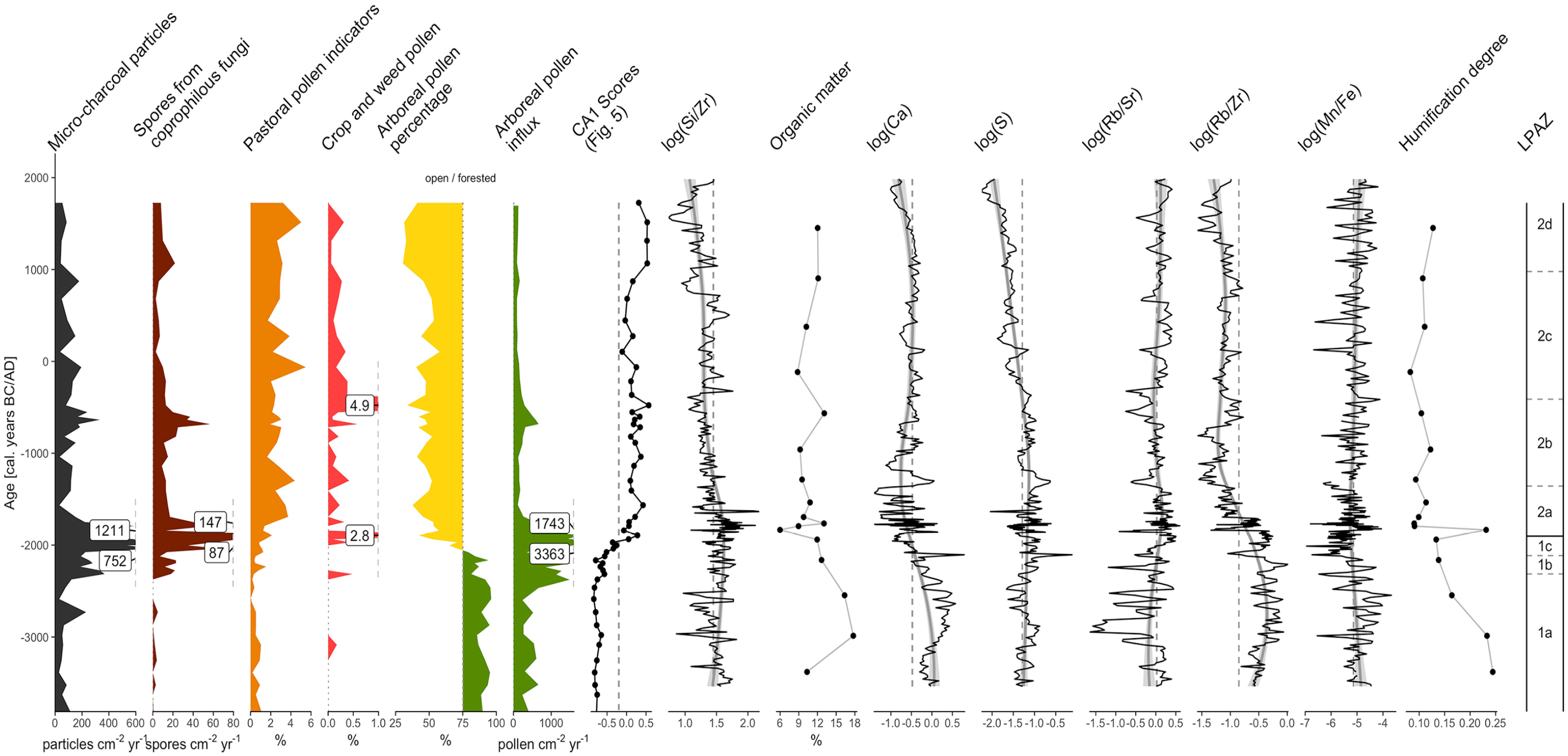
Diagram of a selection of geochemical proxies measured from the Plan da Mattun fen (Urschai Valley, Canton of Grisons, Switzerland, core PMM-2A/B, 2280 m a.s.l.), including chemical elements, organic matter (OM) and humification degree (HD), together with selected palynological proxies, site scores on CA1 (Figure 5) and the LPAZs. If not presented as specific element ratios (e.g. Si/Zr), individual element counts were first normalised to the total counts. Dashed lines represent the overall mean of the proxies displayed, except for site scores on CA1 where a threshold of −0.2 was chosen. Solid grey lines and envelop represent LOESS smoothers and their confidence intervals, respectively. Arboreal pollen denote the opening of the landscape following the threshold value (75%) for forested vs open landscape as proposed by Magny et al. (2006). Graphic by Benjamin Dietre, August 2019.
At the beginning of the record, the Si/Zr ratio was close to its overall mean but progressively increased thereafter with maximum values from 2350 to 1750 BC. A continuous decrease followed, however, with a phase of slightly higher values (closer to the mean) occurring from 300 BC to AD 700. Except for the very first value (10.3%, 3400 BC), the organic matter content (OM) was initially high (16–18%, 3000–2550 BC) but decreased quickly down to 6% at 1850 BC. It then slightly increased to 9–12% for the rest of the record, with a notable higher value (13%) at 550 BC. At the beginning of the record, the fen peat was rich in calcium (Ca), but its values rapidly decreased thereafter. The lowest values were observed at 2000–1000 BC. A slight recovery occurred at AD 500, with values close to the overall mean of the record, before Ca decreased again. Sulphur (S) showed a similar pattern as Ca, except that no phase of low values was observed from 2000 to 1000 BC. However, a continuous decreasing trend was recorded since 1200 BC. Despite the high variability at the beginning of the record and the lowest values at 2950–2800 BC (around −1.5), the Rb/Sr ratio gradually increased from 3250 to 1650 BC. After that, it exhibited lower variability and values close to its overall mean. The Rb/Zr ratio showed relatively high values before 2000 BC; however, it rapidly decreased until 900 BC. From there on, it remained low, except for a period of slightly higher values during the first millennium AD. The Mn/Fe ratio showed a subtle trend with a high variability. It remained stable throughout the entire record, close to its overall mean, but with an inflection towards lower values at 2350–750 BC, and minimal values around 2000 BC. The HD was high at the beginning of the record (0.24), but decreased rapidly down to 0.09 at 1800–1750 BC, except for a peak value at 1850 BC (0.23). After that, it slightly increased to reach 0.13 at AD 1450. In addition, Pearson’s correlation coefficients were calculated for pairs of geochemical proxies. Calcium and sulphur show a correlation coefficient of 0.45; Ca and Bromine (Br) a coefficient of 0.72.
Discussion
The primeval forest of the Urschai Valley
The high amounts of more than 85% arboreal pollen recovered at Plan da Mattun (composed of trees and shrubs potentially growing at or above 2000 m a.s.l. such as Pinus – including Pinus cembra and P. mugo – Picea abies, Abies alba, Larix decidua, Alnus viridis, Salix, Juniperus; Figure 4) suggest that the landscape of the Upper Urschai Valley was situated within the timberline ecotone and that the area was covered with forests from 3900 to 2350 BC, that is, during and up to the end of the Neolithic. The timberline must have been close to, or even above, the Plan da Mattun fen (2280 m a.s.l.) when compared with subfossil Pinus cembra tree stems found at 2360 m a.s.l. in the Las Gondas Bog (5 km NE of the Urschai Valley; Nicolussi, 2018). In addition, this compares well with the reconstructed forest elevation in the nearby Kauner Valley (2300 m a.s.l., 4500–2200 BC; Nicolussi et al., 2005). Magny et al. (2006) suggested a minimum threshold value of 75% of arboreal pollen to imply a local presence of subalpine forests in the Eastern Alps. At Plan da Mattun, arboreal pollen exceeded this threshold value of 75% from 3900 to 2050 BC (Figure 6), and the CA suggests as well that the vegetation was mainly composed of trees and shrubs prior to 2100/1950 BC. This, and the graduate decline of tree pollen influx between 2300 and 1700 BC (Figure 6), advocates that a primeval forested landscape was locally present before 2300 BC, and that no alpine meadows were existing for livestock. This supports the archaeological interpretation of, for example, the Neolithic Abri Urschai site, where all archaeological, archaeozoological and archaeobotanical information (see above) argue for the presence of seasonal camps of hunter-gatherers using wild animal and plant resources until the third millennium BC (Kothieringer et al., 2015; Reidl, 2015; Reitmaier, 2017).
The ratio between rubidium and strontium (Rb/Sr, Figure 6) reflects a proxy for chemical weathering and erosion in the catchment (Jin et al., 2006). This is based on the assumption that Sr can substitute calcium (Ca) in carbonate rocks and co-precipitates in lakes (Cohen, 2003), while Rb can substitute potassium (K) in siliciclastic sediments, especially in clay minerals. Hence, high Rb/Sr ratios in lake or mire/fen sediments suggest an increased erosion of the catchment. This can be either due to wetter climatic conditions with more precipitation in the catchment and thus increased influx of Rb or cooler conditions and thus less Ca weathering and less evaporation in the wetland. Lower Rb/Sr ratios would consequently indicate drier and/or warmer climatic conditions. However, erosion (and thus Rb/Sr) varies also with changes in vegetation cover or human land use. The Rb/Sr ratio is highly variable before c. 2000 BC with generally lower than average values, particularly from 2950 to 2800 BC, suggesting less erosional activity due to either warmer or drier climate or less human activity, which compares well to Europe-wide climatic scenarios (Haas et al., 1998; Magny, 2013). After 2000 BC, the variability in Rb/Sr is weaker, with values slightly above the overall mean, suggesting more erosion due to either cooler or wetter conditions or due to less vegetation or more intense human land use. The increase in erosion is corroborated by the coarsening trend shown by decreasing Rb/Zr ratios. This again agrees with the record from the Kauner Valley, where the treeline decreased down to 2200 m a.s.l. at 2200 BC (Nicolussi et al., 2005). Assuming that fen peat becomes more humified under oxic conditions, the HD may correlate with the Mn/Fe ratio. High HD values (before 1800 BC) indicate relatively drier hydrological or climatic conditions, while a lower HD suggests more water-logged conditions due to a wetter climate (Zhong et al., 2010). However, we only obtained a low number of photometric HD measurements, which is not sufficient for drawing major conclusions.
Alpine farming in the Urschai Valley
While our geochemical record suggests that climatic forcing might have been responsible for a retreat of forested lands in the Upper Urschai Valley, it was probably not the main reason. The high amounts of micro-charcoal particles found at Plan da Mattun at 2300–1750 BC (Figure 6) point to important local to regional fire events, most likely of human origin in order to establish pastoral areas. The resulting prehistoric landscape openness was contemporaneous to the fire-induced openness at Saglias (1420 m a.s.l.; 10 km to the south of Plan da Mattun) in the lowlands of the Lower Engadine Valley (Dietre et al., 2017). However, limitations inherent to the analysis of micro-charcoal particles (possible fragmentation during preparation, and potential long-distance transport) prevent us from being more specific regarding the proximity of such fires to the Plan da Mattun fen (Conedera et al., 2009). Nevertheless, spores of coprophilous fungi were continuously recorded at Plan da Mattun since 2300 BC, and in particularly high quantities and diversity around 1900 BC (Figures 4 and 6). Spores of coprophilous fungi are local markers of livestock grazing in mountainous areas (Cugny et al., 2010; Dietre et al., 2012). Together with the abundance of pollen indicators for pastoral activities, they suggest that pastoral practises started in the Upper Urschai Valley at 2300 BC and were responsible for the steady opening of the landscape and subsequent vegetation change between 2300 and 1700 BC. This is again in agreement with the Saglias record, where the forested landscape of the lowlands was cleared to establish agricultural fields (2900–1500 BC; Dietre et al., 2017). It also agrees with alpine-wide large-scale uses of fire and with the synchronous expansion of permanent lowland settlements and seasonal pastoral exploitation of the uplands (mainly by small ruminants) both in the Lower Engadine and throughout the Inner Alps (Gilck and Poschlod, 2019; Pini et al., 2017; Reitmaier, 2017; Tinner et al., 2005). According to the results from the CA (Figure 5), the threshold point of the transition from primeval forest to open pastoral lands was between 2100 and 1950 BC, but our palynological data (including the influx; Figure 6) suggest it was a long-term process.
The Rb/Zr ratio is often used as sedimentary grain size indicator (Heymann et al., 2013), with a high Rb/Zr ratio representing fine sediments and a low ratio reflecting coarse sediments. The decreasing trend of Rb/Zr from 2700 to 900 BC (Figure 6) indicates a slow but steady coarsening of the clastic sediment fraction in the fen peat. This coarsening might initially have originated from climatic forcing, but its acceleration around 2000 BC probably related to erosion processes favoured by livestock grazing around the Plan da Mattun fen. It corresponds well with the increased accumulation rate revealed by the radiocarbon dates and the age-depth model from 2300 to 1800 BC (Figure 3)
An additional clue to support the development of pastoral activities in the Upper Urschai Valley is given by the findings of cereal-type pollen grains. Cereals were cultivated during the Late Neolithic and Bronze Age in the Engadine (Gobet et al., 2003; Jacomet et al., 1999; Zoller et al., 1996), adjacent Müstair Valley (Brombacher et al., 2007) and the nearby Montafon (Röpke et al., 2011) and Vinschgau Valleys (Schmidl and Oeggl, 2005). According to the archaeobotanical records of the Eastern Alps barley (Hordeum vulgare), wheat (Triticum dicoccum) and millet (Panicum miliaceum) were cultivated up to an altitude of 1700 m a.s.l. and processed for straw, stew and bread production (Jacomet et al., 1999). It is therefore somehow surprising to find cereal-type pollen in high quantities at Plan da Mattun (2280 m a.s.l.) with amounts of 2.8% at 1900 BC or 4.9% at 500 BC (Figure 4). However, these findings may be explained by epi- or endozoochory in the context of vertical transhumance systems, where livestock may transport spores or pollen from lowland areas, either attached to their coat, or ingested and subsequently deposited as faeces. From mountain and alpine areas in Norway, Moe (2014) reported 35,000–63,000 pollen grains (all taxa) per gram of dry wool on the back and the belly of sheep, as well as more than 1 million pollen (all taxa) per gram of dry faeces. After a winter stay at low altitudes in the Lower Engadine Valley, small livestock would be brought up for summer grazing to high-altitude pastures and would carry important amounts of pollen, including cereal-type ones. This idea agrees with records from the French Alps, where pollen from ruderals and plants cultivated in the lowlands were found at higher altitude along trackways (Argant et al., 2006), while amounts of 5% of cereal-type pollen were only found in cultivated fields in the same area (Court-Picon et al., 2006). Indeed, the two peak values of cereal-type pollen found at Plan da Mattun correspond well in time with those found in the lowlands of the Engadine Valley, and particularly for those at Chanoua 8 km to the south (6%; 1630 m a.sl.; Zoller et al., 1996), a locality directly connected to the Urschai Valley via the Tasna Valley (Figure 1). The permanent settlements and terraced fields for agriculture would probably have been located around the prehistoric sites of Ardez-Suotchastè, Ftan-Umbrain and Ftan-Pednal (Reitmaier, 2017).
Livestock farming may have been an important economic/subsistence strategy in the Urschai Valley since the Late Neolithic/Early Bronze Age until the Iron Age as suggested by the important amounts of spores of coprophilous fungi and pollen from plants typical for pasture together with the archaeological record of seasonal campsites and dry stone structures for shepherds (Figure 1). Fireplaces and simple campsites were used during the Neolithic, whereas from the Middle to Late Bronze Age onwards, the first more permanent dry stone structures and pens appeared in this area. These new buildings may not only have served as seasonal shelters for the shepherds but also for processing milk into durable products. This socio-economic shift is similar to the scenario suggested for the Alpine Schnals Valley (Putzer et al., 2016) and for other European high-mountain areas such as the French Alps (Walsh and Mocci, 2011) and the Pyrenees (Galop et al., 2013; Orengo et al., 2014). Later, pastoralism in the Urschai Valley may have been less intensive (or less recordable) during Roman Times and the Early Middle Ages (Figure 4, LPAZ 2c, 400 BC–AD 950). During this phase, fewer spores of coprophilous fungi were found, and a subtle reforestation was observed. Simultaneously, the Rb/Zr ratio at Plan da Mattun (Figure 6) followed an increasing trend, suggesting a lesser coarsening of the clastic sediment fraction, and therefore pointing to less erosion processes. Archaeological evidence from Roman Times and the Early Medieval period (15 BC–AD 1050) is also scarce in the Silvretta Alps compared with the Lower Engadine Valley (Figure 1), suggesting a reduced interest in high Alpine meadows for pastoral use, or a different exploitation strategy, eventually related to large-scale shifts both in populations and in traffic and commerce routes.
Finally, the first written documents mentioning several villages and alpine pastures (with pre-Roman toponyms, for example, Jam, Fimba/Fenga, Id; see Kathrein, 2018) in the Lower Engadine Valley from AD 900 onwards do reflect the renewed agricultural interest. During the Medieval period (AD 1050–1350), the increased number of documented archaeological sites in highland areas (Figure 1) does also evidence this new interest towards alpine pastures, additionally reflected in the palynological and geochemical data. Important amounts of spores from coprophilous fungi as well as pollen indicating pastoralism were found again after AD 950. The palynological assemblages revealed a more open landscape, and the decrease in the Rb/Zr ratio also suggested a trend towards more erosion. New forms of mowing systems and haymaking may have allowed larger quantities of winter fodder and thus an increase in the number of livestock.
Alpine regional pastoral dynamics and landscape change due to dairy production since 1300 BC
The practises of prehistoric, high-altitude livestock farming associated with vertical transhumance may have required the processing of milk and its eventual transformation into storable forms such as fresh or hard cheese (see also discussion in McClure et al., 2018). This process essentially relied on upland dairy production in simple dry stone structures (huts, enclosures) and formed the basis for trade of durable food products such as cheese or butter. The analysis of lipid residues from pottery found in high altitudinal archaeological sites in the Silvretta Alps above 2000 m a.s.l. gave strong evidence for dairy production using cattle and for a fundamental shift in animal husbandry strategies since the Late Bronze Age/Early Iron Age (Carrer et al., 2016; Reitmaier, 2017; Reitmaier et al., 2018). However, here we suggest that the very first pastoral activities in the uplands above 2000 m a.s.l. started earlier at 2300 or 1900 BC at the latest (Late Neolithic/Early Bronze Age) when our palynological and geochemical data point to the beginning and perpetuation of pastoral activities and vertical transhumance in the Urschai Valley. This is in line with the changes in land occupation strategies observed at the Neolithic/Bronze Age transition, leading to a longer and more regular presence of people and livestock (mainly sheep/goat) in high altitudes and the use of seasonal campsites (Kothieringer et al., 2015; Reitmaier, 2017). This also compares with simulations based on subsistence models that suggest that vertical transhumance had become necessary to adapt and respond to the needs of a growing population and specialised economies (Reitmaier and Kruse, 2019). This scenario relates well with the supraregional Bronze Age settlement history throughout the Inner Alps (Reitmaier, 2017). In addition to climatic, economic and demographic reasons, the expansion of settlements was probably due to an increased demand for land for mixed farming, new commercial opportunities (mining of ore and salt as new key economic strategies) and control of the alpine passes, which saw the highest degree of inner and transalpine mobility and exchange during this time. This large-scale synchronous trend and the alpine subsistence based on an agro-sylvo-pastoral economy (mixed Alpine farming) led to a fundamental transformation of the Silvretta Alps – and European Alps in general – into a clearly structured and settled cultural landscape, strongly marked by human activity (Reitmaier, 2017). Our palaeoecological data, local pre-Roman toponyms (e.g. Jam, Fimba/Fenga, Id; Kathrein, 2018) as well as archaeological evidence from periodically used campsites beneath rock shelters and the first excavated remains of prehistoric buildings (Reitmaier, 2017) all point towards treeless alpine meadows that were grazed with sheep, goats and cattle at least from the Bronze Age onwards (Reitmaier et al., 2018). This alpine livestock herding has contributed fundamentally to the modification and diversification of plant communities due to grazing and produced indirect manuring of alpine meadows and soils. Moreover, at a later stage, it also led to the creation of more permanent buildings and first stone-built pastoral structures in the uplands of the Silvretta Alps: huts for the herders, fences, enclosures and byres for the livestock, and perhaps shelters for storing the dairy products. These micro-regional economic changes did probably relate to large-scale cultural and social-economic developments in the Eastern and Central Alps during the Middle and Late Bronze Age, in particular to the formation of the Laugen-Melaun culture. The typical small burnt offering place from this period located at Plan da Mattun is an important evidence not only for a different economic use of the alpine space but also for a newly structured perception of memory and territory within the engagement in higher altitudes (Walsh and Mocci, 2011). Further down in the Urschai Valley, as well as in the adjacent valleys (e.g. Fimba Valley), several buildings for prehistoric shepherds and their livestock were erected during this time, mainly connected with the processing of milk (Reitmaier, 2017). By this, for the first time, shifting economic strategies of dynamic high-altitude pastoral exploitation in the Silvretta Alps become evident, spanning a period of two millennia.
Conclusion
Thanks to the association of palynological, geochemical and statistical analyses from a 5900-year-old fen peat stratigraphy, the flora and vegetation history, altitudinal shifts in treeline and timberline, environmental conditions and the impact of prehistoric livestock grazing in the highland landscape of the Silvretta Alps were reconstructed for the subalpine Urschai Valley in Eastern Switzerland since the Neolithic. An important and steady opening of the arboreal vegetation towards Alpine pasture happened during the Late Neolithic/Early Bronze Age transition (2300–1700 BC). While regional climatic conditions may have contributed to this major transition, fire setting by the first permanent prehistoric settlers and Alpine farmers for the purpose of pastoral activities with sheep, goat and cows was probably the main cause. In addition, our results provide evidence for a distinct economic strategy of seasonal vertical transhumance already during prehistoric times and at least since the Late Neolithic. Dairy production evidenced in adjacent areas of the Silvretta Alps and starting with the Late Bronze Age/Early Iron Age (1300–500 BC) seems related to the synchronous emergence of first dry stone buildings in the Silvretta Alps and probably throughout the European Alps during the second millennium BC. We conclude that the complete pastoral system changed at the end of the Late Bronze Age towards an intensified cattle herding and milk and dairy production (as secondary products) in the subalpine zone above 2000 m a.s.l., and that therefore stone buildings and livestock enclosures were repeatedly used for these purposes. This highlights a range of dynamic economic strategies in the pastoral use of alpine meadows by prehistoric people and also highlights the sustainable use of the Alpine cultural landscape since millennia and up to the present day.
Footnotes
Acknowledgements
This study is part of the international and interdisciplinary project ‘Climate Change in the Alps – Junior Research Groups/10.000 years of climate and settlement dynamics in the Silvretta Massif between Paznaun (Austria) and Lower Engadine (Switzerland)’ and was supported by the Herzog-Sellenberg Foundation and Dr. Erich Ritter Foundation, Germany (2010–2014). Many thanks go to Thilo Kappelmeyer and Walter Bielowski (University of Innsbruck, Austria), as well as to Bertil Mächtle (University of Heidelberg, Germany), for their great help in coring the Plan da Mattun fen peat stratigraphies. The authors would also like to thank Werner Kofler (University of Innsbruck, Austria) for his help in laboratory work, and Daniela Festi and Notburga Oeggl-Wahlmüller (University of Innsbruck, Austria), as well as Timothy Taylor (University of Vienna, Austria), for fruitful discussions on the subject of prehistoric pastoral impact. They much appreciated the discussions with Dagfinn Moe (University of Bergen, Norway) on the endo- and epizoochory processes, as well as with Michel Magny (University of Franche-Comté, France) on the 4.2-ka climatic reversal in Europe. They are grateful to Ana Ejarque (University Clermont Auvergne, France) and an anonymous reviewer for their constructive comments that greatly contributed in improving the first version of the manuscript.
Author contributions
JNH, TR and KL initiated and designed the study; BD, TR, DR, CW, TW and IU compiled the datasets and analysed the data; IH was in charge of the radiocarbon dates; all co-authors wrote and commented on the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This study was funded by the Herzog-Sellenberg Foundation and Dr. Erich Ritter Foundation, Germany (2010–2014; grant no. T021/20506/2010). The printing of colour figures was funded by the Archaeological Service of the Canton of Grisons, Switzerland. Open access to this publication was funded by the University of Innsbruck, Austria.