
Abstract
The search for the subjective nature of our self is intensely debated in philosophy and neuroscience. However, despite all progress, the subjectivity of self and how it fits into the seemingly objective world remains elusive. Drawing on recent empirical data, we show how the self is shaped by the brain’s scale-free activity, that is, long-range temporal correlation (LRTC) and the world’s ecological context. We assume that the scale-free LRTC of the world–brain relation provides the ontological basis for the point of view as the foundation of subjectivity within the world. We conclude that the temporal, that is, scale-free based point of view through the world–brain relation provides the ontologically necessary a posteriori condition for the subjectivity of self on a deeper neuro-ecological level. This extends phenomenological concepts like subjectivity and world beyond both Heidegger’s fundamental ontology and Sartre’s phenomenological ontology: it complements their subjectivity-based ontologies with a truly world-based ontology.
What is the self? The concept of self has long been discussed in philosophy and psychoanalysis, and, more recently, in neuroscience. Since at least the beginning of modernity in the Western world, the concept of self has been conceived as the placeholder of basic human subjectivity including the mind, consciousness, and self-consciousness in particular. This basic subjectivity was targeted in philosophy by, among others, Kant and his German idealistic successors (Fichte, Schelling, Hegel). More recently, subjectivity has surfaced extensively in philosophy in both phenomenology and philosophy of mind.
Briefly, phenomenology associates basic subjectivity with what is described as prereflective self-consciousness (Gallagher & Zahavi, 2019; Zahavi, 2005). In contrast, philosophy of mind often characterizes subjectivity by “what is it like,” which Thomas Nagel attributes to a particular point of view (see below for details) in his famous article, “What is it Like to be a Bat?” (1974).
Parallel to philosophy, there has been abundant empirical research on the self in both contemporary psychology and neuroscience. For instance, the underlying neural correlates of various cognitive processes associated with the self, like self-esteem, self-recognition, self-monitoring, and self-referential processing are investigated and debated (Gallagher & Daly, 2018; Northoff, 2016; Sui & Humphreys, 2015). The present article aims to reconcile and connect the subjectivity and objectivity of self—we are therefore operating right at the cross-roads between philosophy and neuroscience.
Subjectivity of the self: From world and brain to point of view
There is a gap between philosophy and neuroscience, though. Both disciplines are not talking about exactly the same kind of self. Philosophy focuses predominantly on the self as a placeholder of subjectivity: Why and how is it possible for something as subjective as the self to exist in an otherwise purely objective world? In contrast, neuroscience, following the conceptual split between subjective self (“I”) and objective self (“me”; James, 1890; Northoff, 2016), focuses more on the objective psychological and neuronal aspects of the self while leaving out its most basic subjectivity (as that is supposedly not amenable to scientific third-person investigation; Klein, 2014; Sui & Humphreys, 2015). That leaves us with a dichotomy between the subjectivity of self in philosophy and the objectivity of self in neuroscience.
How can we bridge the gap between subjective and objective features of self? The aim of our article is to make a first proposal for bridging this gap by introducing three key concepts: (a) the world–brain relation, (b) the neuro-ecological self, and (c) the point of view. Converging these three key concepts reveals the source of the basic subjectivity of self: the self is ontologically situated and embedded within the world (i.e., “being in the world”; Gallagher & Daly, 2018, p. 3, footnote 3; Heidegger, 1927/2008) through what is often described as a “point of view” (Nagel, 1974). At the same time, the point of view can be well connected with the more objective psychological and neuronal characterization of self as related to the brain and how it relates to the world in a scale-free way (i.e., world–brain relation).
Our main argument is that the concept of world–brain relation (see below and Northoff, 2016, 2018) can well account for the subjectivity of self in an ontological way, namely through the point of view that situates the self within the world. This ontological situatedness of the self within the world is, empirically, mediated by the brain’s temporal features (i.e., scale-free activity) which connect it to the world. Accordingly, the first part of our paper provides empirical support for a neuro-ecological concept of self. That, in the second part, is extended to the ontological domain where the subjectivity of the neuro-ecological concept of self is traced to its point of view which, in turn, is supposed to be based on the temporal scale-free nestedness of the brain within the world.
Nonreductive neuro-philosophical approach: Point of view as an ontological bridge between world, brain, and subjectivity
Phenomenology introduced the notion of prereflective self-consciousness as the most basic feature of subjectivity as manifest in consciousness (Zahavi, 2005). This was further extended to the world by subsequent phenomenologists like Heidegger and Sartre who assumed an existential foundation of the self in the world as reflected in “being in the world” (Heidegger, 1927/2008) and a phenomenological ontology (Sartre, 1948/1993). At the same time, neuroscience made considerable progress in recent years in showing that our sense of self is closely related to the brain (Northoff et al., 2006; see below for details). Moreover, there is not only one self but multiple concepts of self, which may be related to distinct neural features (Frewen et al., 2020; Gallagher & Daly, 2018; Qin et al., 2020).
Can we converge the phenomenological and neuroscientific perspectives? This would require a bridge between the world-based subjectivity of self in the phenomenological context and the neuroscientific evidence of the brain-based self. More specifically, it would mean that we need to integrate the brain, as dealt with in neuroscience, with the concept of world in such way that their relationship (i.e., world–brain relation; Northoff, 2016, 2018) permits a truly world-based subjectivity of self that is aimed for in phenomenology. First and foremost, this raises the question of which methodological strategy allows us to link them.
We currently pursue a nonreductive neurophilosophical approach (Northoff, 2014a, 2014b, 2016, 2018). Rather than reducing or simply identifying empirical and phenomenological concepts, we search for empirically plausible bridging concepts that provide necessary (a posteriori) connections between world-based subjectivity and brain-based self. We postulate that one such concept is the point of view. As implied by Nagel’s (1974) description of the bat, the point of view situates and locates us within the world which, as we see it, engenders (but cannot be equated with) the first-person perspective as a key feature of consciousness. This makes the point of view a viable bridge between the world on the one hand and the first-person perspective of consciousness on the other—it may thus provide the source of world-based subjectivity, that is, the link between world and consciousness.
How is the point of view related to the brain? To address this question, we first need to go back to the brain and how it is related to the world. Ontologically speaking, the brain is part of the world as a whole with both standing in a mereological relationship. Given that the brain is part of the world, both the brain and world must share certain features while, at the same time, remaining distinct. Drawing on the recent development of temporo-spatial ontology (Northoff, 2018), we assume that the world and brain share their temporo-spatial features, that is, they stand in a temporo-spatial relationship to each other with the latter being nested within the former (just like the smaller Russian doll nests within the next larger one). Without being able to elaborate the details, this marks temporo-spatial ontology as a form of relational ontology (as distinct from property-based ontology; Northoff, 2018). World and brain are thus ontologically linked by a temporo-spatial relation. Ontologically conceived, the brain is therefore viewed in an intrinsically relational and henceforth neuro-ecological way, rather than purely neuronally. We employ the term “ecological” in a double sense. Taken in the context of the subjectivity of self, the term ecological refers to the environment as we perceive it; the environment in a subjective sense—this is in line with the concepts of environment introduced by Gibson (1967, 1979) and Lewin (1936). Here, the environment is intrinsically subjective as it is related to, and perceived by, the subject itself. However, we aim to extend beyond that. Specifically, we raise the question of how such an environment, taken in a subjective sense, is constituted. This leads us to the world understood in an ontological sense, independent of any subject. Hence, the term “world” refers to the world as it is prior to the subject (i.e., mind-independent) as distinguished from the subjective or mind-dependent environment.
At the same time, the world in this mind-independent sense provides the necessary conditions for the possible constitution of the subject (i.e., the predisposition of its possible existence and reality in the world as featured by its subjective environment within the objective world). We consider the brain, understood ontologically, to be key in mediating that predisposition of the world (in an objective sense) to constitute the subjective nature of self including its subjective environment. This amounts to what we describe ontologically as world-based subjectivity for which we consider the world–brain relation a key feature. Our paper can be understood as an ontological investigation into these relationships as distinguished from a more psychological approach as in ecological psychology, where the concept of world is understood in a more subjective sense, that is, as environment.
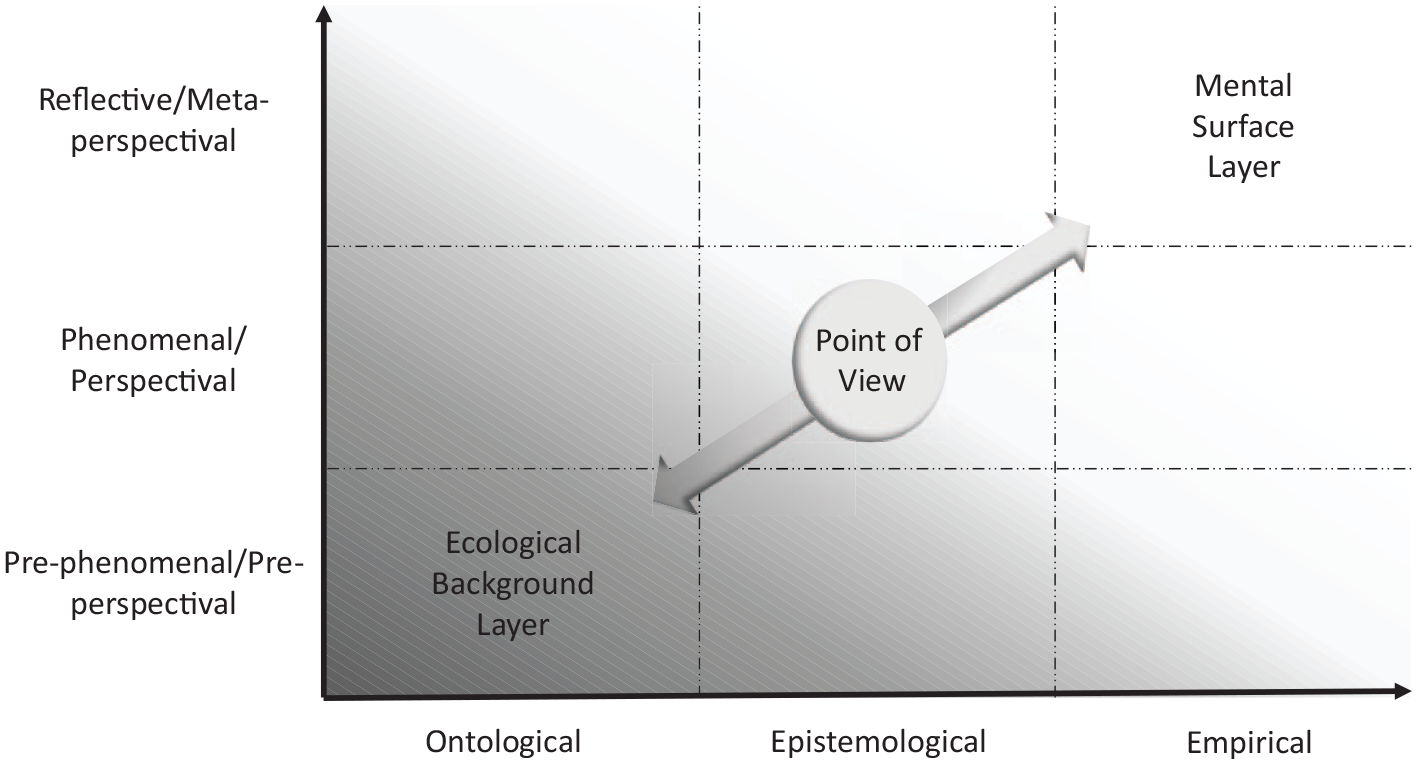
The goal of our article is now to connect both perspectives, that is, the determination of world-based subjectivity with a point of view with the ontological view of the brain in terms of the world–brain relation characterized by temporo-spatial features. For that purpose, we characterize the point of view in temporo-spatial terms, which provides the ontological bridge between world, brain, and subjectivity (see Figure 1 for a general overview).
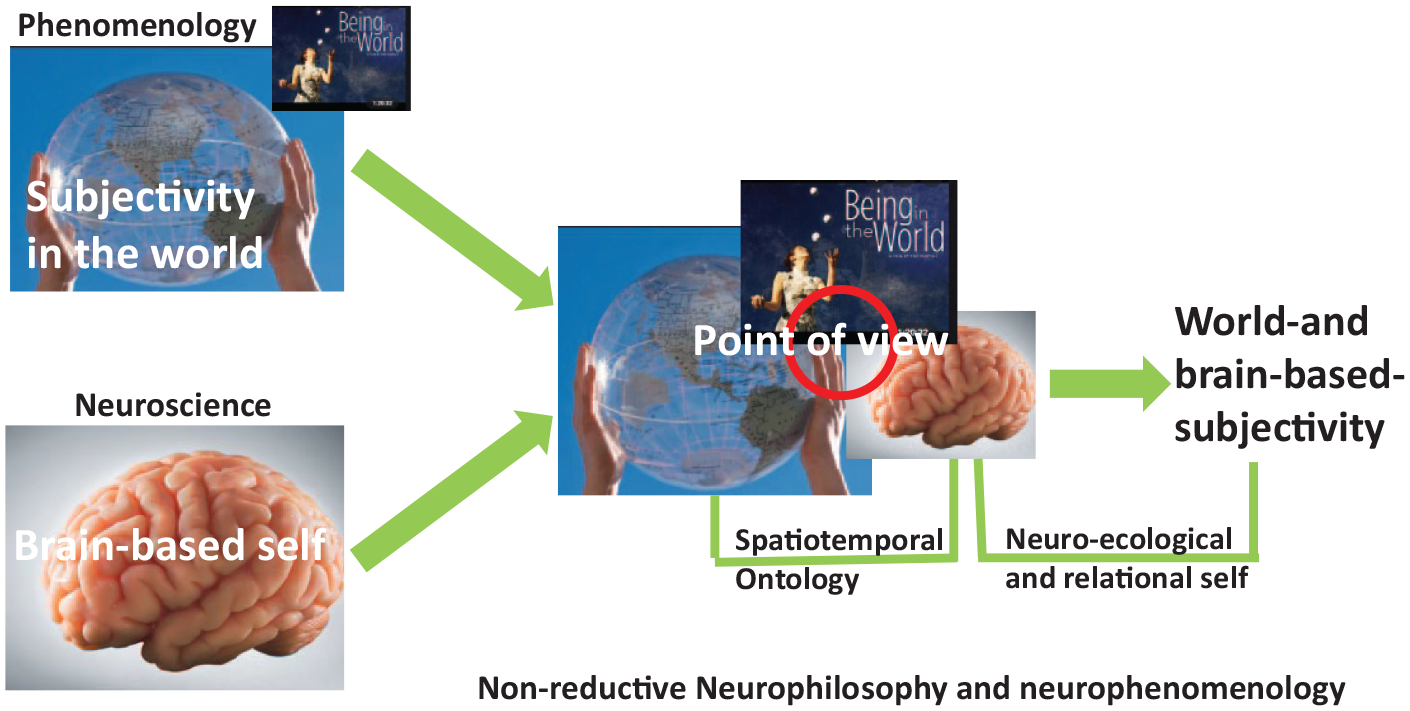
Overview of the main claim – convergence of phenomenological (upper) and neuroscience (lower) approaches to subjectivity (left) within the point of view (middle) as a basis for world-based subjectivity (right).
Overview of the article
We first focus on the empirical side by discussing neuroscientific data that characterize the brain-based nature of self in temporo-spatial (i.e., scale-free) terms, which is highly dependent upon the environmental context as revealed by traumatic life events and cultural shaping. This points to the neuro-ecological (rather than purely neuronal) nature of the brain and the self, which carries important conceptual implications. These data illustrate the need to conceive of the brain in terms of a world–brain relation, which can serve as the ontological basis of the self.
Note that we currently only target a most fundamental layer of self: the neuro-ecological self. In contrast, we leave out various other conceptualizations such as the minimal self or prereflective self-consciousness (Gallagher & Daly, 2018) and other concepts of self like bodily and cognitive self (Frewen et al., 2020). Instead, our approach targets a deeper layer, that is, the neuro-ecological self featured by its point of view, that, as we assume, provides the most basic layer shared by these various concepts of self. Finally, it shall be noted that it is beyond the scope of this article to discuss all these different concepts of self and how they relate to the neuro-ecological self. Instead, our paper focuses on sketching the neuro-ecological self and how that, being based on the world–brain relation, is related to the point of view.
The second more conceptual aim is to connect the neuro-ecological self and world–brain relation with the point of view as the basis of world-based subjectivity. The focus here is on elaborating the concept of point of view: it is situated within the world (i.e., neuro-ecological) while, at the same time, providing a necessary (a posteriori) connection to the brain (i.e., world–brain relation) as the basis or source of world-based subjectivity.
Next, we will distinguish different notions of the point of view (and, importantly, will characterize it in ecological and temporo-spatial terms) which permits a connection to the temporo-spatially determined world–brain relation. That, in turn, makes it possible to hypothesize that the temporo-spatial features of the point of view provide the necessary a posteriori connection between the world–brain relation and world-based subjectivity. We therefore conclude that an ontological and temporo-spatial concept, the point of view, is a viable candidate to bridge the gap between phenomenological conceptualizations of world-based subjectivity of self on the one hand, and neuroscientific views of the brain-based nature of self on the other.
World, brain, and self: Empirical evidence of a scale-free neuro-ecological self
How is our self shaped by its environment? We will present two lines of empirical evidence linking psychology and neuroscience that demonstrate how the environment and brain interact to shape the self. The first empirical line takes a more general view by showing how the brain’s spontaneous activity is characterized by scale-free (see below for explanation) temporal features and how those are related to the self. The second line of empirical evidence concerns the impact of early childhood traumatic experience on the self through the brain’s resting state (i.e., its spontaneous activity). Note that we focus primarily on the link between self and world (i.e., neuro-ecological self) while neglecting other concepts such as minimal and bodily self (Frewen et al., 2020; Gallagher & Daly, 2018). Furthermore, we will omit details regarding the generative mechanisms underpinning scale-free activity as this currently remains an open issue (He, 2014).
The scale-free brain: Scale-Free temporal structure of the brain’s spontaneous activity
The brain’s spontaneous neural activity can be characterized by different frequencies ranging from infraslow (0.01–0.1 Hz), over slow (0.1–1 Hz), fast (1–40 Hz), to ultrafast (40–180 Hz); Buzsaki, 2006). Power is strongest in the infraslow range and decreases across the slow, fast, and ultrafast ranges following a power law distribution (He, 2014; He et al., 2010; Huang et al., 2016). Together, the different frequencies and their distinct degrees of power constitute a complex temporal structure in the brain’s spontaneous activity which, in large parts, can be featured by the balance between infraslow, slow, and faster frequencies.
The relationship between these frequencies is maintained across different temporal scales and can therefore be characterized by what is described as “scale-free dynamics” (He, 2014; He et al., 2010; Linkenkaer-Hansen et al., 2001). Roughly, scale-free activity describes the fractal (i.e., self-similar) organization and thus temporal nestedness in the relationship between power and the different frequency ranges: the longer and more powerful slower frequencies nest and contain the shorter and less powerful faster frequencies—this amounts to long-range temporal correlation (LRTC), which operates across different time scales or frequencies (He, 2014; He et al., 2010; Linkenkaer-Hansen et al., 2001; Northoff & Huang, 2017; see Figures 2a and 2b). This makes it clear that we here understand scale-freeness in explicitly temporal terms, that is, in terms of frequencies and their power spectrum; in contrast, we refrain from any spatial exploitation of scale-free activity as it has recently been criticized on methodological grounds (Barabasi, 2009; Broido & Clauset, 2019).
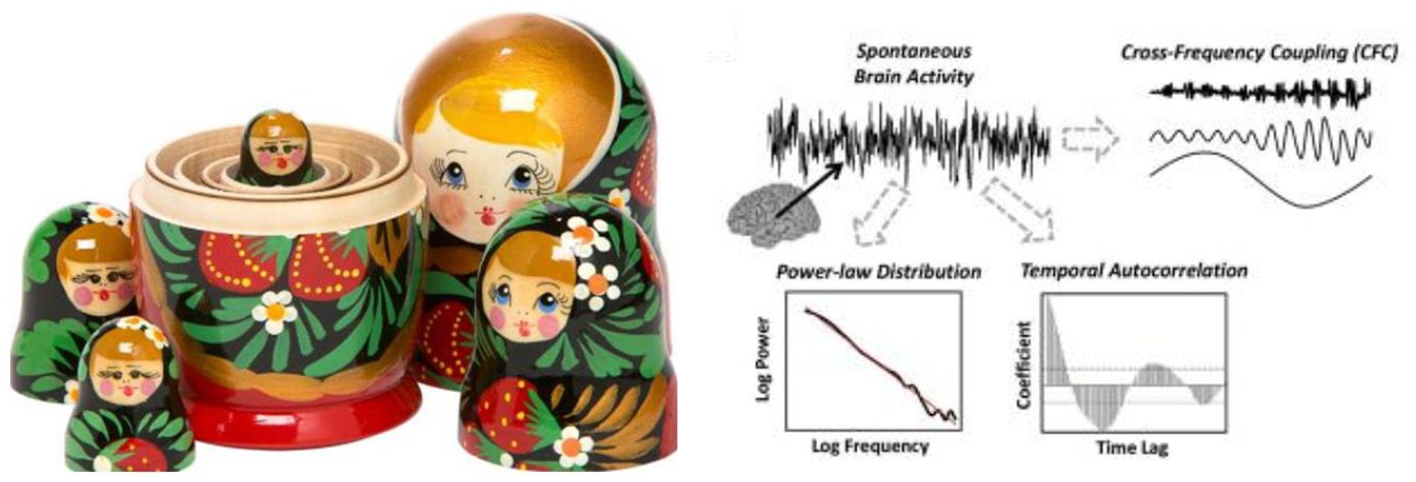
Nestedness of Russian dolls (left) and scale-free activity in the brain (right).
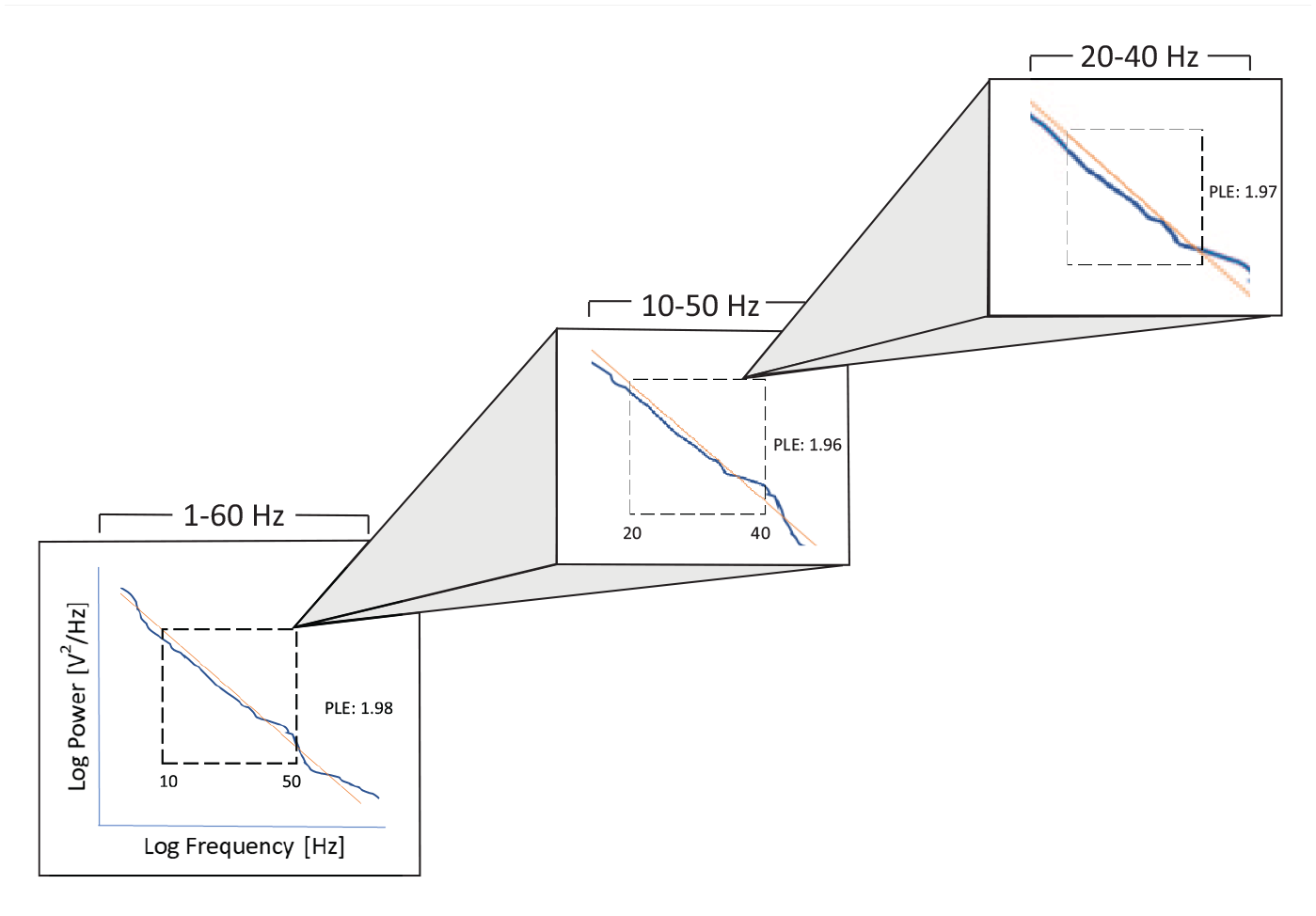
Scale-Freeness of the power spectrum with temporal nestedness.
The LRTC can be described as scale-free or scale-invariant that can be expressed by, P ∝1/fβ, where P is power, f is frequency, and β is called the power-law exponent (PLE; He, 2014). A high PLE value indicates relatively stronger power in slow frequencies and relatively less power in the faster ones, whereas the opposite is the case in a low PLE value. As an alternative to the PLE that accounts for the power relationship across different frequencies, one can also probe the LRTC in the temporal domain by measuring oscillatory amplitude fluctuations using detrended fluctuation analysis (DFA; He et al., 2010; Linkenkaer-Hansen et al., 2001; Palva et al., 2013; however, see Bryce & Sprague, 2012, for criticism of scale-free activity).
Independent of their differences, both PLE and DFA measure neural activity across multiple time scales and are therefore indices of scale-free or scale-invariant activity. That makes it possible to assess the degree to which past neuronal patterns exert their influence on future dynamics, thus accounting for LRTC (Linkenkaer-Hansen et al., 2001; Northoff & Huang, 2017). High values in PLE/DFA indicate high degrees of LRTC, with long stretches of neural activity exhibiting temporal correlations between past, present, and future states—this reflects the relatively stronger impact of slower frequencies with their longer cycles. The opposite is the case in low PLE/DFA values, where only the most recent time intervals exert any impact on present and future ones—this is manifested in the relatively stronger impact of faster frequencies with their shorter cycles.
The scale-free self: Temporal nestedness and long-range temporal correlation (LRTC) on neuronal and mental levels
Recent studies have shown that the brain’s scale-free activity, as measured with either PLE or DFA, is related to mental phenomena such as the self (Huang et al., 2016; Scalabrini et al., 2019; Scalabrini et al., 2017; Wolff et al., 2019; see Figure 2c). These studies show that the degree of resting state PLE directly predicts: (a) the degree of self-consciousness (as operationalized by the self-consciousness scale; Huang et al., 2016; Wolff et al., 2019), (b) task-related activity during self-specific stimuli (like animate versus inanimate touch; Scalabrini et al., 2019), and (c) the degree of temporal integration on a psychological level of self-specificity as measured by a matching task where participants have to associate the own self, nonself, or celebrity with a particular geometric shape (Kolvoort et al., 2020).
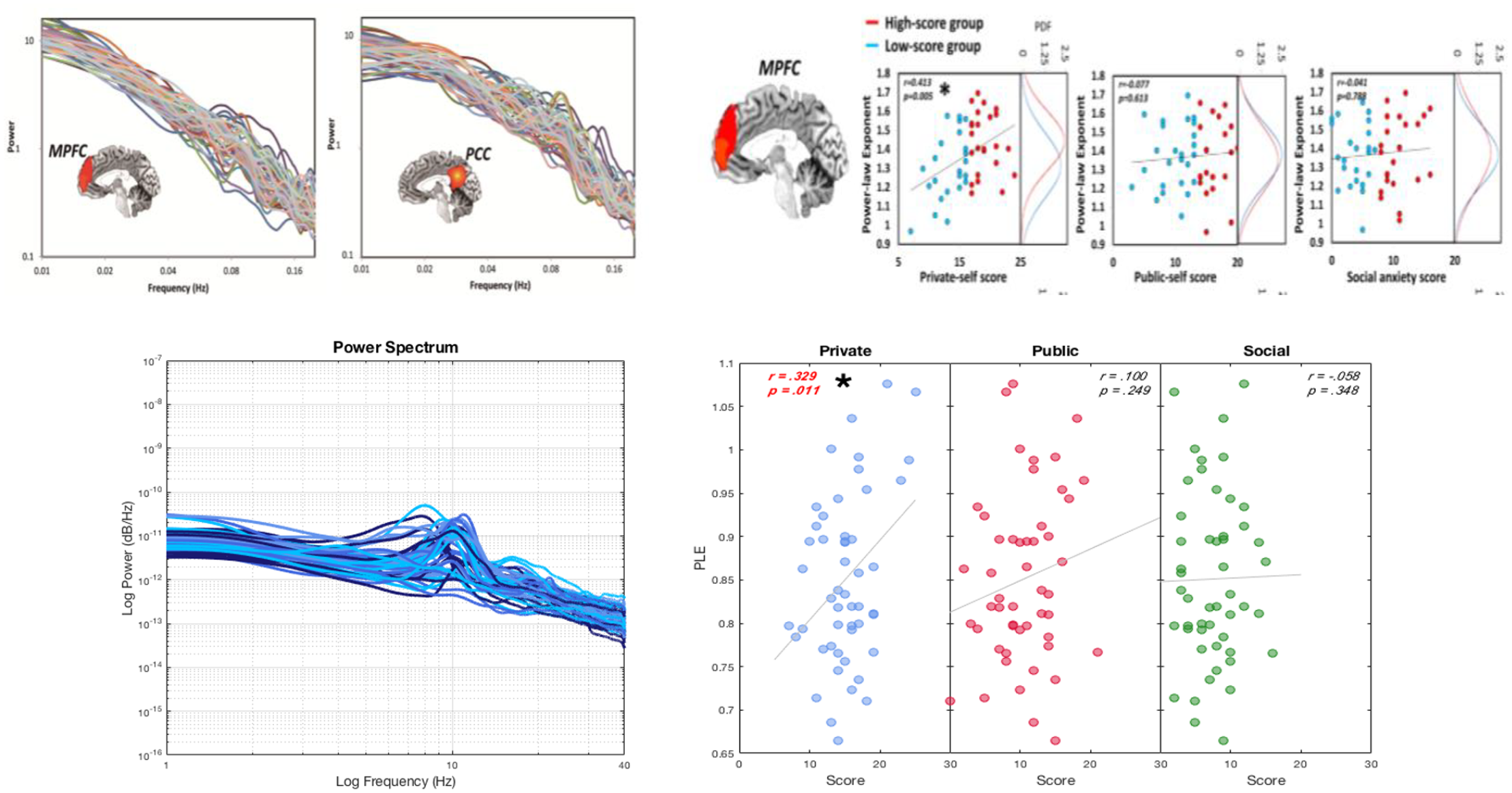
Scale-free activity is related to self in infraslow frequencies of fMRI (upper/Huang et al., 2016) and faster frequencies of EEG (lower/Wolff et al., 2019).
Given that in all these studies it is the resting state’s scale-free activity that correlates with different psychological tasks all probing for self-specificity, we assume that the resting state scale-freeness seems to be central in mediating distinct components of self-specificity like self-consciousness, task-related activity, and temporal integration. Moreover, additional studies have demonstrated that scale-free activity is central in mediating other mental features such as consciousness (Northoff & Huang, 2017; Zhang et al., 2018) and mental abnormalities in psychiatric disorders like autism (Damiani et al., 2019) and schizophrenia (Northoff et al., 2020).
Taken together, these findings suggest that the brain’s scale-free properties are central for mental features and thus, more generally, the mind including self and consciousness. Mental features such as self and consciousness seem to operate across different time scales by integrating and nesting them within each other. Applying this terminology to the self in a novel way, the self can be considered scale-free and can therefore be characterized by temporal nestedness and long-range temporal correlations (LRTC) on a mental level, which seem to find their analogues on the neuronal level in the brain’s spontaneous activity.
The neuro-ecological self: Traumatic life experience shapes the brain’s spontaneous activity
We all experience adverse life events in both childhood and adulthood which shape our self. How can traumatic experiences, especially those in early childhood, shape our self in adulthood? Various brain imaging studies have shown that early traumatic childhood experiences impact the spontaneous activity’s temporo-spatial dynamics later in life. For instance, one fMRI study by Lu et al. (2017) compared participants with and without a history of childhood trauma. They demonstrated changes in both intra- and interregional synchronization (e.g., regional homogeneity and functional connectivity) in regions of the default-mode network (DMN) and salience network (SN) such as the insula in participants reporting early traumatic experiences. Somewhat analogous resting state functional connectivity in the insula and related regions of the SN was also observed in a study by Gupta et al. (2017).
The impact of traumatic life experiences on the brain’s spontaneous activity is further corroborated by results in studies examining posttraumatic stress disorder (PTSD; Disner et al., 2018; Koch et al., 2016). One recent large-scale meta-analysis showed that here too, regions of the DMN (e.g., the inferior parietal lobule [IPL]) and the SN (e.g., the amygdala and caudate) exhibit decreased resting state activity (i.e., intra-regional synchronization and neural variability) in subjects suffering from PTSD (Disner et al., 2018). Focusing more on interregional changes (i.e., functional connectivity), a meta-analysis by Koch et al. (2016) observed disbalance between decreased resting state functional connectivity (rsFC) in the DMN and increased rsFC in the SN.
Additional studies demonstrate how early traumatic experience does not only impact the resting state but that the latter, in turn, shapes the adult subject’s task-related activity. For instance, one fMRI study by Duncan et al. (2015) first measured entropy (i.e., the degree of disorder) in the resting state in adult participants reporting early traumatic childhood experiences. They observed that the degree of resting state entropy in a region of the DMN (anterior cingulate cortex) directly correlated with the degree of early life traumatic experience: the more participants experienced early trauma, the higher their entropy in resting state activity during adulthood (Duncan et al., 2015).
In a second step, the same study investigated task-related activity by applying an aversive stimulus (i.e., an electric shock). They observed that participants reporting a high degree of early traumatic experience exhibited reduced somatomotor cortex and insula activity in anticipation of the shock. Critically, this was directly modulated by increased entropy in the DMN (i.e., anterior cingulate cortex; see Figure 3). Accordingly, as anticipation implicates spontaneous or resting state activity during the prestimulus period, these data show how abnormal imprinting of the spontaneous activity by traumatic life events affects how participants respond to aversive stimuli in their environment.
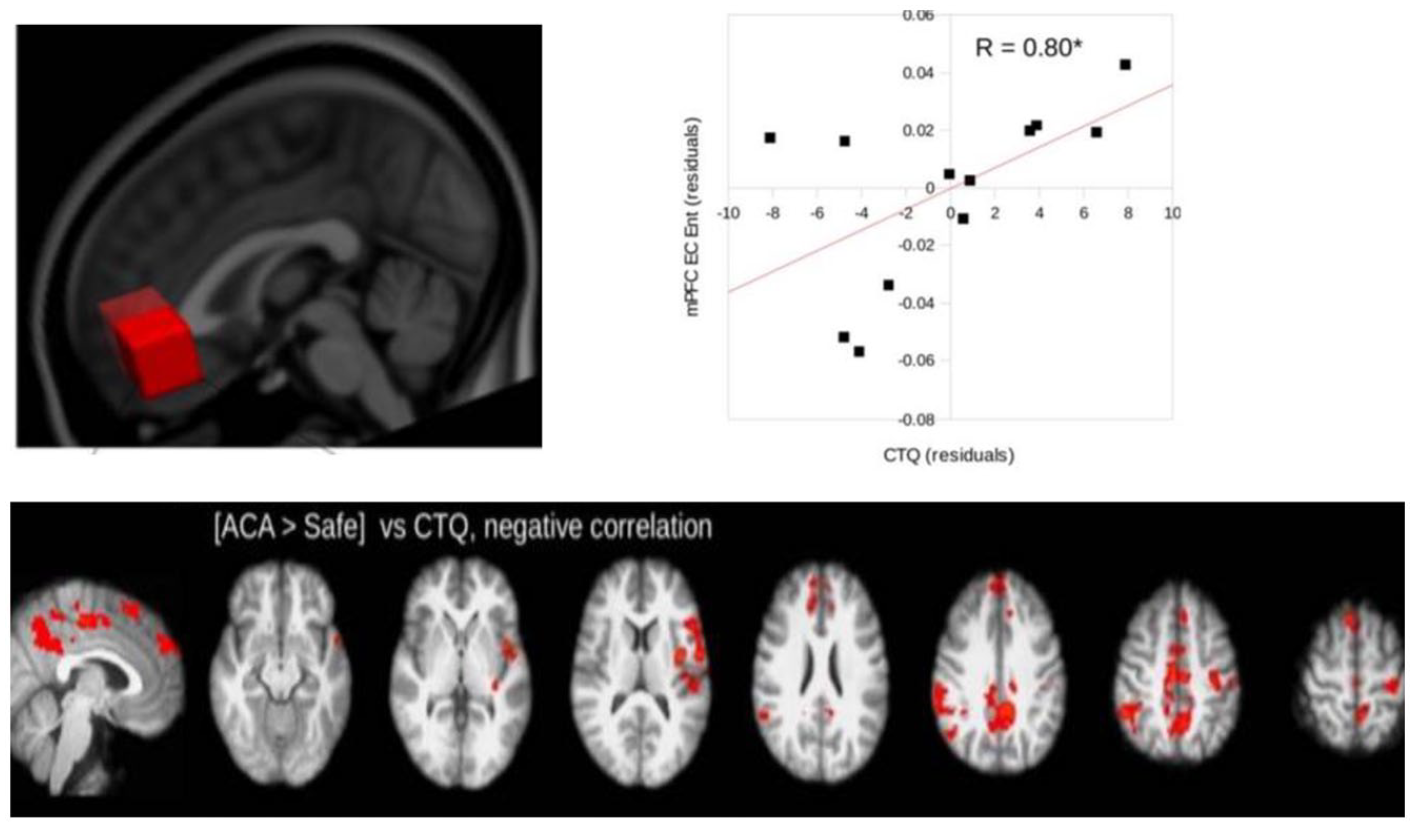
Early childhood trauma modulates the brain’s ongoing and task-related activity (Duncan et al., 2015).
Another study by Nakao et al. (2013), using near-infrared spectroscopy (fNIRS), demonstrated that power in the very low infraslow frequency ranges (< 0.04 Hz) was negatively related to early traumatic life experience in the anterior DMN (i.e., medial prefrontal cortex): the higher the degree of early traumatic experiences, the less power in infraslow frequency ranges in the resting state of the medial prefrontal cortex during adulthood.
Nakao et al. (2013) also included two tasks: one task involving the self (i.e., color preference judgment), and one task not involving the self (i.e., color similarity judgment). Interestingly, the degree of early traumatic life experience only correlated with infraslow frequency power in the medial prefrontal cortex during the self-related task: lower infraslow frequency power and fewer self-related judgments correlated with a higher degree of early traumatic experience.
Together, these studies clearly demonstrate that early traumatic experience strongly shapes both resting state and task-related activity in adulthood. In particular, this concerns brain regions such as the DMN (i.e., medial prefrontal cortex, anterior cingulate, posterior cingulate) and SN (i.e., insula, amygdala) that have been implicated in processing self-specificity (Qin & Northoff, 2011; Qin et al., 2020). Self-specific, self-related, or self-referential processing means that a stimulus is processed relative to and in its meaning for the self, that is, its relative proximity or distance to the own self—the degree of neural activity may then be dependent upon the degree or strength of the relation between self and stimulus (Northoff, 2011, 2016). The own face, for instance, is usually processed in closer relation to the own self than to another person’s face.
We can consequently assume on empirical grounds that the self in adolescence and adulthood is strongly shaped by its subjectively perceived environmental context (see above for the conceptual distinction between environment and world) through the brain’s spontaneous activity and its temporo-spatial dynamics in a scale-free way (i.e., spanning both short and long timescales). In more conceptual terms, there is empirical support for: (a) the world–brain relation, with significant life events shaping the brain’s spontaneous activity; (b) the brain’s spontaneous activity, in turn, shaping the self in a neuro-ecological way through its own modulation by significant life events; and (c) such neuro-ecological shaping operating in a scale-free way, that is, across different time scales.
We should mention, however, that these findings concern the impact of the childhood environmental context on the adult self. There is a long timespan between childhood and adulthood. What we want to illustrate is that, despite this interval and the numerous changes in the self from childhood to adolescence, the effects of the former can still be seen in the latter and its brain’s spontaneous activity. Albeit tentatively and on conceptual grounds, we therefore assume some temporal continuity between the self in childhood and the self in adulthood that, on a deeper layer, connects them with each other even though they, on a more surface layer in terms of their mental contents, may markedly differ.
On the more empirical side, this raises the question of whether there are analogous contextual influences on the self during adulthood. This is indeed the case. For instance, cultural contexts are known to shape our self and its brain. Asian cultural contexts are signified by a more interdependent self that is defined through its relation to and similarities with others (Han & Northoff, 2008; Markus & Kitayama, 1991). Western contexts show a more independent self characterized by its distinction to others (see also Scalabrini et al., 2021). Notably, these cultural differences in self go along with neural differences in the brain including in their scale-free activity (Han & Northoff, 2008). Together, these empirical findings strongly support our conceptual assumption of the intrinsic context dependence of brain and self and, more generally, the neuro-ecological nature of both. These conceptual implications shall be pointed out in the next section.
World, brain, and self: Conceptual implications of the empirical findings
Shared features of brain, self, and world: Scale-Free temporal structure
What do the brain, weather, seismic waves, and stock markets have in common? Prima facie, you will insist that they are unrelated; the brain is a lump of grey matter consisting of neurons, which of course, cannot be observed in the others. Despite their differences on the surface, these seemingly disparate phenomena nevertheless show a degree of commonality on a deeper level. Namely, fluctuations in their activity exhibit the same scale-free structure characterized by temporal nestedness and LRTC.
One of the most interesting aspects of scale-free activity is its universality. Scale-free activity is not unique to the brain; rather, it is ubiquitous in nature, evident across systems as varied as climate, seismic activity, magnetic fields, and stock markets (Cocchi et al., 2017; He et al., 2010). Basically, wherever irregular fluctuations in activity are observed, LRTC and scale-free activity may provide structure to what initially appears to be random noise. There is “structure to irregularity,” and that seems to be a unifying principle and key feature of nature.
For instance, He et al. (2010) investigated the scale-free dynamics of the brain’s neural activity and its nested frequencies using electrocorticography (ECoG). In addition, they investigated the time series of spontaneous seismic activity collected over 4 months and fluctuations in the Dow Jones index obtained over a period of 80 years. Time series from both seismic waves and stock market fluctuations followed a power-law distribution in their temporal power spectrum. Interestingly, their power-law exponents (1.99 for seismic waves and 1.95 for the stock market) resembled that of the brain’s intrinsic activity measured in ECoG during wakefulness (mean of 2.2 for < 0.1 Hz). Of further significance, like the brain’s intrinsic activity, the time series of both seismic waves and stock market fluctuations contained nested frequencies (i.e., higher frequency fluctuations nesting in lower frequency fluctuations).
Neuro-Ecological self: Long-Range temporal correlations of the brain connect self and world
How are the world’s scale-free features related to those of the brain? Given that the world can shape the brain through traumatic life events, one would expect a close relationship, if not an interaction, between their respective scale-free features. Specifically, one would predict that the LRTC of the environmental structures contain and nest those of the brain’s spontaneous activity—both ecological and neuronal LRTC would be expected to match across (and despite) their different time scales. Such “complexity matching” has indeed been shown in recent studies on language and music.
Borges et al. (2018), for instance, demonstrated how the degree of scale-freeness in the brain in different frequency bands follows variations in the scale-free envelope of speech, and how their degree of correspondence impacts speech comprehension. Analogously, Borges et al. (2019) show how the scale-free structures of the brain and music adapt to each other, with the brain’s neural activity somewhat rescaling the musical structure. The timing of the neuronal fluctuations, as observed in the brain’s scale-free activity, followed the timing of the fluctuations in the music (i.e., its scale-free activity), although the latter operated on a wider range of temporal scales than did the brain. Moreover, the scale-free adaptation of participants’ neural activity to the music predicted their degree of pleasure. Together, these findings show that the degree of matching or concordance of the scale-free properties in the environmental context (i.e., music in our case) and brain strongly shape one’s mental features like the feeling of pleasure and the sense of self. This suggests a truly neuro-ecological basis of subjectivity within the world, mediated by what we describe as the world–brain relation.
Conceptual characterization of the neuro-ecological self: Self is nested within the world through the brain’s long-range temporal correlations
Taken together, scale-free activity is ubiquitous in nature rather than being a unique feature of the brain. This allows the brain to compare and match its own scale-free nestedness and LRTC with those of its respective environment, with the degree of their matching shaping mental features like perception. Unfortunately, no studies have been reported that investigate whether the degree of matching between world and brain LRTC is related to the self and how we perceive ourselves to be part of the world (i.e., the neuro-ecological self).
We nevertheless may want to make a more philosophical or theoretical claim: the fact that the brain’s scale-freeness is directly related to the self implies that the self is connected and integrated within the world’s scale-free structure through temporal nestedness and LRTC. Analogous to how the smallest Russian doll is integrated within the next larger one and so forth, the self is integrated and nested within the brain which, in turn, is ontologically nested and integrated within the largest and most comprehensive temporal scale: the world. On conceptual grounds, we can thus say that what connects the self through the brain to the world is its scale-freeness featured by temporal nestedness and LRTC.
To summarize, we can say that: (a) the world–brain relation is scale-free and characterized by temporal nestedness and LRTC, implying that the world’s larger scale nests the brain’s smaller scale; (b) the scale-free nature of the world–brain relation may be critical in shaping and constituting the self; (c) the self is intrinsically neuro-ecological and scale-free; and (d) the self is intrinsically integrated within the world through the temporal nestedness and LRTC of the world–brain relation.
These theoretical but empirically plausible considerations imply that we may also need to change our conceptual determination of the self. Instead of a purely neuronal concept of self, we may need to view the self through a neuro-ecological lens entailing a biological–ecological approach as described, for instance, by ecological psychology (Bruineberg et al., 2018; Heft, 2020a, 2020b); this, through the world–brain relation, can be extended to neuroscience (i.e., ecological or temporo-spatial neuroscience; Northoff et al., 2020). Admittedly, we omit the psychological and neuronal details related to the concept of the world itself—concepts like affordances, ecological niche, and others that describe different layers within the world (Heft, 2020a, 2020b) will need to be explored in their relevance for both the world–brain and world–self relations.
Point of view between world and brain: Conceptual characterization of its ecological background layer
Point of view: Mental surface layer and ecological background layer
What is a point of view (POV)? The notion of POV is pervasive in literature and theatre, with different persons expressing different points of view on the same topic. Painting and photography rely on a slightly different notion of POV that involves providing access to events or objects in the world. Despite the extensive colloquial usage of POV in many disciplines, the concept is often neglected in philosophy, as there is no established theory (see Campos & Gutierrez, 2015, for a notable exception). Yet another noteworthy exception is the embodied approach where the lived (rather than objective) body is supposed to provide the point of view or anchor of the self in the world (Gallagher, 2005; Gallagher & Daly, 2018). Without being able to go into details (see Northoff, 2016, 2018), we assume that the notion of the lived body presupposes the scale-free nature of the brain: the body’s various temporal scales are connected with those of the brain’s spontaneous activity as well as with those of the world that are processed through the brain. Hence, conceptually considered, the brain can be conceived as a multiscale integrator as it connects the different timescales of the brain, body, and world in a scale-free way. If, for instance, the brain’s capacity to integrate these timescales in a scale-free way is diminished (i.e., indexed with PLE or DFA), we lose consciousness as in that case, the neuro-ecological connection of brain and self to body and world is severed and thus, conceptually speaking, the point of view is lost (Zilio et al., 2021).
We will introduce Campos and Gutierrez’s (2015) account of POV and then provide our own ecological and ontological extension. Our main argument will be that the concept of POV can provide the source of subjectivity by constituting an intrinsic, necessary connection between the world and the self—the necessary intrinsic connection of world and self as mediated by POV can ultimately be traced to the necessary connection of world and brain, the world–brain relation (Northoff, 2018). In a nutshell, we propose that the world–brain relation as the basis of the point of view provides the most fundamental ground of subjectivity within the world.
According to Campos and Gutierrez (2015), POV can be determined by two main features: reference to mental life (including subject), and access to the world beyond the self and its mental life. Following in their footsteps, we will reformulate and rename these two features as the surface and background layers of POV, respectively. The POV’s reference to the subject and mental life is the surface layer of POV—we call this a mental surface layer. At the same time, the POV is situated within the world as its ultimate ontological background, which renders it ecological—we therefore additionally posit an ecological background layer. Let us detail these two concepts in the following.
The mental surface layer of POV refers to a subject with personal and mental features; importantly, this layer provides the source or basis for the mental features of self: In that variety of uses, the notion of point of view may have two distinct meanings. In one of them, points of view are part of a mental life. They are connected to the mental life of some subjects with a personal character. In that sense, the expression “point of view” is interchangeable with words like “view”, “opinion”, “belief”, “attitude”, “feeling”, “sentiment”, “thought”, etc. Points of view in that sense could not exist without a subject with quite a rich mental life. (Campos & Gutierrez, 2015, p. 2)
In contrast, the ecological background layer of POV is characterized by providing access to something that lies beyond the POV itself, namely the world with its ecological features. Rather than on mental states within the subject itself, the focus here is on how the subject connects and relates to the ecological features of the world. Intra-subjectivity is therefore replaced by intersubjectivity, and isolation is replaced by relation: There is another quite important meaning in the ordinary notion of point of view. In that second sense, points of view could exist without any actual subject exemplifying them. Here, points of view explicitly have a strong relational and modal, especially subjunctive, character. Points of view offer possibilities of having access to the world. They offer possibilities of seeing things (hearing them, touching them, etc.), possibilities of thinking about them (considering them, imagining them, etc.), and possibilities of valuing them (assessing them, pondering them, etc.). (Campos & Gutierrez, 2015, p. 3)
Ecological background layer of point of view I: Scale-Free relation of world and self
We argue that the ecological background layer of POV is key in providing the ontological ground of subjectivity and ultimately of the self. This distinguishes our approach from both past and present philosophical and neuroscientific approaches that, usually, claim the phenomenal and mental features of self (and hence the mental surface layer of POV) to provide the source of subjectivity (see our conclusion where we situate our approach in a methodological–historical context).
The key feature of the ecological background layer of POV is its relational character, as it relates and connects the self to and within the world. Relation means that the POV is connected and related to something beyond itself. Put into the context of ecological psychology, that “something beyond itself” is the environment characterized by its ecological features. This includes natural, social, and cultural kinds of information along with their descriptive and normative aspects—for the sake of simplicity, we will lump them all together under the notion of ecological information understood in a broad sense.
The ecological background layer of POV includes that ecological information that it shares with the world. The world itself contains at least two types of ecological information: that which is shared with the organism and that which is not shared with the specific organism, thus extending beyond the latter. Importantly, this entails partial rather than total overlap since the sharing between world and brain, and ultimately of world and POV, is incomplete. Consider the timescales. An organism has a limited repertoire of timescales compared to the world. For instance, we as humans cannot directly perceive the ultraslow seismic waves preceding earthquakes, nor can we perceive the ultrasonic frequency ranges accessible to bats.
The overlap in the amount of ecological information between the world (as a whole) and the ecological background layer of POV (as part of the world) constitutes an ontological relation between the world and POV as this relation (i.e., the overlap in ecological information) defines the existence and reality of POV and consecutively the subjectivity of self. What do we mean by ecological information shared between world and POV? We refer to biophysical features and their related spatial and temporal scales, which can be traced to the brain and how it stands relative to the world. For instance, the bat, based on its biophysical features, shows a POV that enables it to access ultrasonic information (Nagel, 1974). In contrast, the human POV does not allow us to access ultrasonic features, since our brain does not possess the proper biophysical characteristics, as it is ultimately based on different temporal and spatial scales (when compared to those of the bat).
This leads us back to scale-free activity and its contribution to constituting the ecological background layer of POV: the broader the range of temporal and spatial scales encompassed by the ecological background layer of POV, the more extensively the self can relate to the world and its ecological information. We consequently assume that the ecological background layer of POV is nested and contained within the world and its ecological information in a scale-free way, analogous to how a smaller Russian doll is nested within a larger, self-similar version of itself. Scale-free nesting of the POV’s ecological background layer within the world implies that one would expect to find long-range temporal correlations (LRTC) between the world and self: the world’s much longer timescales are related to the self’s shorter time scales in a self-affine and cross-scale way, as the former nests and contains the latter.
We propose that such scale-free LRTC between world and self constitutes the ecological background layer of POV as the source of subjectivity for the self. The concept of “source” refers to a necessary nonsufficient condition, or a predisposition (Northoff, 2018). Moreover, this necessary condition is to be understood not in the traditional philosophical terms as necessary a priori but, drawing on Kripke (1972) and Nagel (1998), as necessary a posteriori (see Northoff, 2018, Chapter 10, for a full development). Moreover, subjectivity is determined here in an ontological (rather than epistemological [Kant], phenomenological [Husserl], or existential [Heidegger, Sartre]) and temporo-spatial way: it reflects the existence and reality of the self in the world, that is, how its own range of temporo-spatial scales relates to the temporo-spatial scales of the world. Moreover, the ontology of subjectivity is scale-free as it operates across the different temporo-spatial scales of world and self. The ecological background layer of POV is the relation between their different temporo-spatial scales as marked by their degree of cross-scale LRTC between world and self based on the organism’s biophysical features (like in humans or bats; Northoff, 2014a, 2014b).
This makes it clear that the ontological nature of self is determined by temporo-spatial relations. Namely, the scale-free relationship between world and brain provides the necessary a posteriori condition for the possible constitution of the neuro-ecological background layer of POV as the basis of self. More generally, this presupposes the determination of existence and reality by relation, rather than by properties, which therefore entails a relational rather than property-based ontology (Northoff, 2016, 2018). This relation-based ontology is now specified in temporo-spatial terms (i.e., Spatiotemporal Ontology; Northoff, 2016, 2018). Applied to self, this means that the neuro-ecological self and its POV are intrinsically relational.
Ecological background layer of point of view II: Scale-Free temporo-spatial relation of world and brain
What about the brain and its role in constituting the ecological background layer of POV? We have seen that the self is scale-free and mediated through the brain’s scale-free activity. The latter, in turn, is strongly shaped by the scale-free activity of the world, entailing the scale-invariant nature of the world–brain relation. Putting it all together, we now postulate that the ecological background layer of POV is ontologically based on the world–brain relation through scale-free activity: the more the world and brain are temporally (and spatially) nested within each other and exhibiting LRTC, the greater the temporospatial range of the ecological background layer.
Ultimately, this broader, more expansive range of LRTC of self with the world (through the brain and world–brain relation) permits greater ecological extension of the self towards and within the world. In contrast, if the temporal range of the ecological background layer is limited, meaning lower degrees of temporal nestedness and LRTC, the self becomes more restricted and isolated in its relation to the world.
Let us consider the comparison of bats and humans. Bats, as pointed out by Nagel (1974), can process ultrasonic waves, which humans and their brains cannot. This means that the world’s LRTC aligns with different timescales in bats and humans, namely the ultrasonic and nonultrasonic, respectively. The bat’s point of view and its neuro-ecological background layer are consequently nested and contained within a different timescale of the world when compared to humans and their POV. The differences in the temporal extension of the bat’s and human’s points of view within and relative to the world leads to differences in their subjectivity, which is manifest in their different “what is it like” (Nagel, 1974). Importantly, the differences highlighted here do not concern the actual mental contents themselves, but rather the predisposition for processing the range of possible contents. Our argument is that bats and humans differ in the ontological predispositions of their POV due to their different timescales. This temporal predisposition for the range of possible contents is then complemented by the actual contents, that is, those contents they are exposed to in their environment, and that fall within the range of their POV-based temporal predisposition. Together, we can see that the mental contents are doubly determined: by the predisposition for a particular range, related to the POV, and the actual contents themselves.
This example points out that the ecological background layer of POV is intrinsically relational: it describes the POV in terms of its temporo-spatial scales and ranges relative to those of the world as a whole. Campos and Gutierrez (2015) hint upon that when describing the POV as a “relational entity.” However, by combining “relational” and “entity,” they remain somewhat ambiguous as their concept of entity means that something is clearly delineated from that which surrounds it. This is not compatible with the intrinsically relational and scale-free nature of the POV’s ecological background layer, though. Without its temporo-spatial relation and relative difference to the world’s temporo-spatial scales, the ecological background of POV would not be possible at all—its existence and reality is dependent upon the temporo-spatial relation. Relation is thus understood in an ontological sense, as it constitutes the existence and reality of the POV as the source of subjectivity within the world. The ecological background layer of POV is purely relational without any kind of entity as apparently still assumed by Campos and Gutierrez.
Given the scale-free integration of the ecological background layer within the world and its scale-free features, it is better to describe POV as a “relational hub” or “node.” Much like hubs or nodes are constituted by their relation or connectivity to the rest of the network, POVs are characterized by their relation or connection to the rest of the world’s temporo-spatial networks. Taken in this sense, the ecological background layer of POV provides “structure to irregularity” by organizing and structuring the world’s various temporal scales for the self within its limited temporo-spatial range such that the latter can access the former’s ecological information in a temporo-spatially limited and compressed way. An analogous prototypical example here is seismic activity. Seismic waves operate on an extremely slow frequency range, the ultra-slow, which we as humans cannot process as such. However, due to clever measurement devices, we are able to compress their timescales and, due to self-similarity related to scale-freeness, are able to decipher certain patterns in these seismic waves that may index the increased risk of potential earthquakes. Based on its own more limited but scale-free timescales, the brain, analogously, compresses the larger timescales from the world and, due to its scale-free nature, processes them in a more or less self-similar way—the ecological information of the world may thus, in partial and compressed form, be restructured and preserved in the brain’s processing and its temporal patterns.
Ecological background layer of point of view III: Intrinsically temporal and scale-free
Traditionally, the point of view has been associated with a fixed entity like mental or physical substances that can be described as static and atemporal. This distinguishes the traditional concept of a point of view from our own characterization of the ecological background layer of POV that, rather than being static and fixed, is highly dynamic and therefore intrinsically temporal. As previously discussed, scale-free activity provides “structure to irregularity.” This means that, even if the activity of different regions or the power of single frequencies changes in an irregular way, the overall structure can maintain its integrity. Therefore, scale-free activity is neither completely irregular nor entirely fixed; rather, it operates on a continuum between the conceptual extremes of total change and total stability (see also Northoff & Tumati, 2019).
This points to a core feature of the POV’s ecological background layer, namely that its intrinsically temporal nature is entailed by its scale-free features. That converges with Campos and Gutierrez (2015) who also characterize POV as intrinsically temporal: Let us say understand temporal points of view as follows: A temporal point of view is a point of view identifying some differences in non-conceptual contents (qualitative, phenomenal, experiential contents) as “changes” of content. The identification can be either conceptual or not conceptual. This is a very important point. Subjects without conceptual capacities could be capable of adopting temporal points of view. In any case, in a temporal point of view certain differences in non-conceptual content count as a “change”: something future becoming present, or something present becoming past. The idea behind that characterisation of temporal points of view is very simple. Temporal points of view take some differences in the non-conceptual contents of experience as being temporal differences entailing a “change”. This is the crucial point. (p. 93)
Where does the time of the POV’s ecological background layer originate? We suppose that the intrinsically temporal nature of the ecological background layer comes from within the world itself (i.e., world-based time; Northoff & Chen, 2019). By connecting and relating to the world in a scale-free way, the ecological background layer of POV participates and integrates with the continuously ongoing construction of time in the world. As its temporal scale is much larger, the world’s time nests and contains that of the POV—the world emplaces POV within its own larger range of timescales, which renders the ecological background layer of POV intrinsically temporal and scale-free.
Ecological background layer of point of view: Prephenomenal and preperspectival neuro-ecological condition of world-based subjectivity
Concept of world: “Emplacement world” and “perspectival world”
What do we mean by the concept of “world”? The intrinsically temporal and scale-free emplacement of the POV’s ecological background layer within the world’s larger temporal scales converges well with the concept of emplacement world proposed by Campos and Gutierrez (2015, pp. 13–14, 40). They distinguish two concepts of world: emplacement world and perspectival world. The former is the world within which POV is situated, while the latter is the world that we perceive and cognize from a certain POV. Without going into too much philosophical detail, we will use these two concepts of world to further characterize the ecological and mental layers of POV.
The emplacement world reflects the background ecological layer of POV; it is the world within which POV is situated, allowing for the subject’s, through its POV, direct contact and relation with the world’s external reality. POV is emplaced in the world’s temporal and spatial structures that constitute the world prior to and independent of POV itself. Briefly, we characterize the emplacement world as intrinsically temporal and scale-free between itself and its various parts such as POV-based subjects or selves that, through scale-free activity with LRTCs, are nested and contained within the world’s much larger temporo-spatial scales. In this sense, the concept of emplacement world is primarily ontological and mereological as it refers to the existence and reality of the world as a whole and how it relates to its parts.
In contrast, the concept of perspectival world is primarily epistemological since it refers to the surface cognitive layer of POV and the world that we as subjects perceive from the first-, second-, or third-person perspective (FPP, SPP, TPP). The self is typically associated with FPP, which is considered to be subjective and can be distinguished from the more objective TPP. While not elaborating on this association, we argue that all three perspectives merely characterize the mental surface layer of POV rather than the ecological background layer that remains preperspectival. Accordingly, all three perspectives, FPP, SPP, and TPP, must be considered subjective rather than objective: constituting the mental surface layer of POV, they are all based necessarily on the ecological background layer of POV as the most basic source of subjectivity within the world. We will illustrate this shift in the characterization of FPP, SPP, and TPP by discussing the notion of prereflective self-consciousness in phenomenology.
How to interpret Thomas Nagel’s “What is it Like to be a Bat?”: First-Person perspective versus point of view
We experience our own self and the world in first-person perspective (FPP). FPP is considered the hallmark feature of both subjectivity and consciousness in phenomenology, as both are conjoint in what is described as prereflective self-consciousness (Gallagher & Zahavi, 2019; Zahavi, 2005). In a nutshell, prereflective self-consciousness describes the immediate and first-personal givenness of experience that is already there prior to and independent of any reflection, attention, or recognition. Even when I am conscious of an event or object in the world, I am already conscious of myself in the prereflective mode. Prereflective self-consciousness signifies the intrinsically subjective nature of our experience and consciousness featured by its first-person perspective and its phenomenal character.
How does prereflective self-consciousness stand in relation to POV? Colloquially, the first-person perspective is thought to be based on a specific point of view, marking FPP as subjective, perspectival, phenomenal, and isolated (from the rest of the world). However, that pertains only to the mental surface layer of POV rather than the ecological background layer, which is intersubjective, ecological, relational, and what we refer to as preperspectival.
Thomas Nagel’s famous example of “What is it Like to be a Bat?” (1974) is usually considered an illustration of the subjective, perspectival, phenomenal, and isolated nature of FPP as distinct from the intersubjective, nonperspectival, nonphenomenal, and nonisolated TPP. Subjectivity and consciousness, including prereflective self-consciousness are here related exclusively to FPP while, at the same time, not being available to TPP. We contest such mental interpretations of Thomas Nagel that equate subjectivity with FPP and thus the mental surface layer of POV. Instead, we argue that Nagel’s example of the bat may be better conceived within an ecological context that traces subjectivity to the ecological background layer of POV.
When Nagel (1974) speaks of a point of view, he does not equate POV with FPP. For instance, this is reflected in the following quote from his famous paper, “What is it Like to be a Bat?”: “I am not advertising here to the alleged privacy of experience to its possessor. The point of view in question is not one accessible only to a single individual. Rather it is a type” (p. 441). Moreover, it is noteworthy that Nagel does not mention the concept of FPP once in this paper. He only refers to a point of view that distinguishes bats’ experience and subjectivity from those of humans. We therefore claim that Nagel presupposes a wider notion of POV that is not restricted to FPP as distinct from TPP. Instead, we postulate that he presupposes the notion of POV in a wider sense, prior to and independent of the distinction of FPP, SPP, and TPP—this is very compatible with the assumption of POV being a necessary condition of the possible differentiation into FPP, SPP, TPP, as we postulate. This carries important implications.
How can we know the ecological background layer of point of view? Preperspectival and prephenomenal access
Presupposing this wider conceptualization of POV means that the ecological background layer of POV can neither be characterized by any particular perspective like FPP, nor by prereflective self-consciousness. Characterizing the background layer in this sense would be to confuse it with the mental (or phenomenal) surface layer of POV as the sufficient condition for FPP and prereflective self-consciousness. We need to characterize POV in nonmental terms distinct from those used to describe its mental surface layer. Instead, we need to revert to ecological terms to describe such wider, more basic concepts of POV.
Without being able to fully flesh out the details, we propose that the ecological background layer of POV is preperspectival and prephenomenal (Northoff, 2014a, 2014b, 2018) as it provides the necessary, albeit nonsufficient, condition of possible perspectives (FPP, SPP, TPP) and phenomenality without realizing them as such. In other terms, the ecological background layer of POV provides the ontological capacity or predisposition of perspectives (i.e., FPP, SPP, TPP) and prereflective self-consciousness (see Northoff, 2018, Chapter 10, for the concept of ontological predisposition). This means that the ecological background layer of POV is preperspectival and prephenomenal rather than being either perspectival and phenomenal by itself or, alternatively, nonperspectival and nonphenomenal (in which case it would no longer provide the predisposition at all).
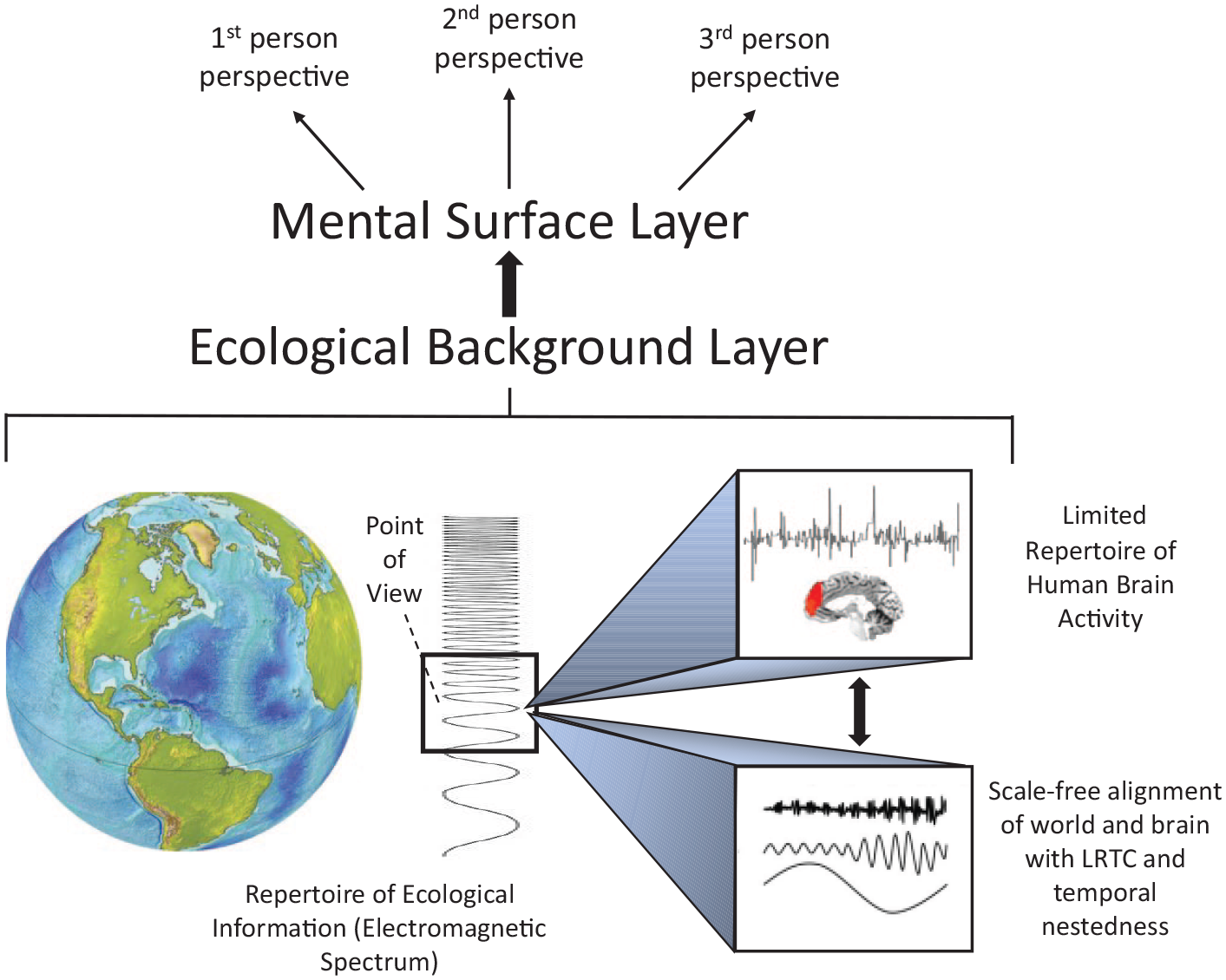
Together with the aforementioned characterization, we assume that the preperspectival and prephenomenal nature of POV’s ecological background layer (as an ontological predisposition of the perspectival and phenomenal nature of self) is based on its intrinsically temporal and scale-free nature that constitutes its relation to, and emplacement within, the world (i.e., emplacement world). This marks the self as preperspectival, prephenomenal, and neuro-ecological (see Northoff, 2014b, for the concept of prephenomenality), which first and foremost predisposes the phenomenal, mental, and cognitive features of self as they can be associated with the mental surface layer of POV (see Figures 4a and 4b). More generally, this means that the prephenomenal and preperspectival nature of POV provides the source of necessary (a posteriori) albeit nonsufficient condition of subjectivity in the world (i.e., world-based subjectivity), as elaborated in phenomenological accounts by Husserl, Heidegger, and Sartre.
Scale-Free nestedness of self within brain and world – world–brain relation and neuro-ecological self.
Basis model of self-specificity (BMSS): Point of view and its scale-free nature as the basis of the subjectivity of self.
Conclusion
Ontological, temporo-spatial, and neuro-ecological view of subjectivity
We are now ready to address the more philosophical issues of how to converge the world-based subjectivity of self in phenomenology with the brain-based nature of self as observed in neuroscience. Albeit tentatively, this conclusion serves to highlight some of those connections based on our neuro-ecological and temporo-spatial notion of the point of view. The key question arising from this is: How do the ontological concepts of world–brain relation, neuro-ecological self, and point of view stand in relation to the concept of subjectivity discussed in philosophy? Without being able to provide a comprehensive account at this time, we will make some overtures in this direction.
The key concept serving as a bridge is the point of view. Following Thomas Nagel, we understand the point of view in a temporo-spatial sense, that is, as a temporo-spatial structure that, being intrinsically scale-free, operates across different spatial and temporal ranges. Critically, we conceptualize time and space in an ontological sense (i.e., as featuring the world’s existence and reality), and how they constitute the world’s relation to its parts like the brain (i.e., world–brain relation)—this makes it unavoidable that the brain is intrinsically neuro-ecological rather than purely neuronal as manifested in the ecological background layer of the point of view.
The point of view, understood in neuro-ecological and scale-free terms, allows the subjectivity of self to be constituted ontologically and temporo-spatially. Specifically, from an ontological perspective, we propose that the world emplaces the ecological background layer of POV through its relation to the brain’s temporo-spatial features in a scale-free way (i.e., by means of temporal nestedness and LRTC); this, in turn, constitutes the situatedness of the self within the world and thereby its subjectivity. This presupposes a relational, rather than a property-based ontology: Relations between the world and brain, rather than singular entities like the world, brain, or body alone are the basic units of existence and reality (Northoff, 2018). This relational ontology is intrinsically ecological, mereological, and temporo-spatial as these are the features through which the world as a whole and its parts like brain and self are related to each other in a scale-free way—one may thus want to speak of a scale-free ontology.
We consider a relational and ecological scale-free ontology to be key in developing a proper ontology of subjectivity and self. We have focused only on the point of view and, specifically, its ecological background layer as to link the self intrinsically to its ecological context within the world. This sets up a novel framework for the development of a neuro-ecological, temporo-spatial, and relational ontological concept of self which, through its intrinsically temporal nature, may then also be closely linked to temporal continuity, long-term temporality, and ultimately, personal identity (Brüntrup et al., 2020; Wagner & Northoff, 2014, 2017).
Finally, conceived in a larger context, the ontological triangle of world–brain relation, point of view, and neuro-ecological self addresses the incommensurability of subjectivity and objectivity in its ontological facet (Nagel, 1974, 1986) that has plagued psychology, neuroscience, and philosophy in both past and present. The dichotomy of subjectivity and objectivity has led to some of the key questions of our time: How can something subjective like a self exist and be real in an objective world? Rather than reducing or parallelizing them, we opt for reconciling and integrating subjectivity and objectivity within a commonly shared ontological framework that, by being temporo-spatial, allows for the connection between world and self through the world’s scale-free temporo-spatial organization that ontologically predisposes the world–brain relation as a necessary, a posteriori condition for the possible generation of POV and its neuro-ecological self.
Avoiding methodological circularity: Subjectivity-Based versus world-based view of subjectivity
We are obviously not the first to search for a deeper background layer of self and subject. Kant introduced a deeper background layer of the subject when distinguishing transcendental and empirical self-consciousness. The transcendental subject was determined mainly in conceptual-logical terms entailing what can be described as “reflective self-consciousness.” Husserl went beyond Kant by replacing the latter’s transcendental level of the subject by focusing on consciousness and its phenomenal level. The subject could now be determined phenomenally rather than in conceptual-logical terms—this shifted the focus from reflective self-consciousness to prereflective self-consciousness (Zahavi, 2005).
We now extend Husserl from prereflective self-consciousness to an even more basic notion of the prephenomenal self (see Northoff, 2014b, for a discussion of the prephenomenal as distinct from the prereflective self). That is possible by introducing the ecological background layer in addition to the prereflective surface layer of POV. The ecological background layer of POV makes it possible to situate and emplace prereflective self-consciousness within the context of the world in a necessary (a posteriori) and intrinsic way. Since that is ontologically mediated by the scale-free world–brain relation, this marks the self as intrinsically neuro-ecological. Importantly, the neuro-ecological character is not experienced as such but provides the necessary condition or predisposition for any possible experience of the self in terms of prereflective self-consciousness (which, phenomenologically, is also often described as the minimal self; Zahavi, 2005). For this reason, we designate the neuro-ecological layer of self as prephenomenal rather than either nonphenomenal or prereflective. Let us explicate that.
Our main claim is that the neuro-ecological layer of self provides the capacity or predisposition for its possible experience in terms of prereflective self-consciousness as distinguished from the latter’s actual realization. This means that the neuro-ecological level cannot be described by prereflective self-consciousness itself, as that would confuse the necessary conditions or predisposition of its possible realization with the sufficient conditions of the self’s actual realization. As the neuro-ecological layer and its point of view cannot be experienced as such, we therefore characterize it as prephenomenal rather than as phenomenal as entailed by prereflective and reflective self-consciousness. The reflective and prereflective self are therefore now complemented by what we describe as the prephenomenal self. This extends Husserl’s phenomenological notion of prereflective self-consciousness to an ontological level, namely to a prephenomenal self based on the neuro-ecological background layer of POV and its scale-free nature which, ontologically, is ultimately based on the temporo-spatial features of the world–brain relation.
The presupposed ontological level is even deeper and more fundamental than what Heidegger described as “Fundamental Ontology.” Unlike Heidegger, the present approach no longer infers the self from the phenomenological level and its existential extensions like Dasein and Being-in-the-world. Rather than taking the self as the point of departure for characterizing its role and place in the world, the present approach takes a reverse stance: it describes the world in temporo-spatial and ontological terms, which serves as a basis for exploring the ontological similarities between the world and self which, as we postulate, can be found in the temporo-spatial and scale-free features of the point of view (and its basis on the world–brain relation). Hence, where Heidegger describes the world in terms of subjectivity (i.e., in existential terms), we, taking a reverse stance, describe the subjectivity of self in terms of the world as featured by scale-free and temporo-spatial relationships. Converging both, we tentatively consider the scale-free and temporo-spatial relation of world and self through the world–brain relation as a necessary (a posteriori) condition of the kind of world-based subjectivity (i.e., Being-in-the-world) that Heidegger and Sartre describe.
The unravelling of a deeper prephenomenal, temporo-spatial, and neuro-ecological layer of the subjectivity of self makes it possible to methodologically extend beyond both Heidegger’s fundamental ontology and Sartre’s phenomenological ontology. Even though they strive for an ontology of subjectivity in the world, both Heidegger and Sartre still determine such subjectivity in dependence on, and in terms of, the concept of subjectivity itself. Specifically, their characterization of the world is still based on the phenomenal and existential features of the self. Methodologically, this amounts to what can be described as a subjectivity-based ontology of the self in the world. However, that cannot avoid being methodologically circular, at least to some extent, as here the ontological account of subjectivity in the world is methodologically dependent and based upon itself (i.e., the phenomenal and existential features of subjectivity).
To avoid any such methodological circularity, we require an ontological concept of subjectivity within the world that, methodologically, remains independent of the subjectivity itself (including its phenomenal and existential features)—a world-based ontology of self in the world that must unravel its prephenomenal (rather than phenomenal and existential) features. Our triangular conceptualization of the world–brain relation, point of view, and neuro-ecological self serves exactly that purpose, namely to establish a truly ontological and prephenomenal, world-based account of the subjectivity of self within the world. While further developments are necessary, we hope that our combined ontological–empirical approach can avoid the kind of methodological circularities that plague the concept of subjectivity in both past and current philosophy and neuroscience.
Footnotes
Declaration of conflicting interests
We have no conflicts of interest to disclose.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: this project is supported by funding from the European Union’s Horizon 2020 Framework Program for Research and Innovation under the Specific Grant Agreement No. 785907 (Human Brain Project SGA2). GN is grateful for funding provided by UMRF, uOBMRI, CIHR and PSI. We are also grateful to CIHR, NSERC, and SSHRC for supporting our tri-council grant from the Canada–UK Artificial Intelligence (AI) Initiative “The self as agent–environment nexus: crossing disciplinary boundaries to help human selves and anticipate artificial selves” (ES/T01279X/1 ; together with Karl J. Friston from the UK).