
Abstract
Despite decades of research on narcissism, its neural basis remains poorly understood. Competing accounts hypothesize either heightened responsiveness to socially rewarding feedback (reflecting motives for status and validation) or neural disengagement (reflecting a self-protection motive). To test these hypotheses, we recorded EEG from 98 participants as they completed Monetary and Social Incentive Delay tasks. We measured an event-related potential, known as the Reward Positivity, to examine whether narcissism was associated with altered neural sensitivity to monetarily and socially rewarding feedback. Results revealed a domain-specific dissociation. In the Monetary Incentive Delay task, narcissism was unrelated to neural responses. However, in the Social Incentive Delay task, rivalry narcissism (and overall narcissism) was associated with reduced Reward Positivity amplitudes, whereas admiration narcissism was not. These findings support the self-protective disengagement account, suggesting that individuals high in rivalry evince blunted immediate neural sensitivity to socially rewarding feedback. This early-stage disengagement may serve to maintain a sense of detachment from the self, while limiting opportunities for social learning and connection.
Plain Language Summary
Narcissism is a personality trait that combines self-confidence, perfectionism, and the belief that one is special. Although narcissistic individuals are often seen as craving attention, psychologists disagree about how they respond to praise. One view suggests they are highly sensitive to compliments, which boost self-esteem. Another argues that narcissists are disengaged, tuning out feedback to protect a detached self-image. Our study tested these ideas by examining brain responses to feedback. We recorded brain activity from 98 students while they completed two tasks. In one, participants could win or lose small amounts of money depending on how quickly they responded. In the other, correct responses were rewarded with a positive social feedback (thumbs up) and incorrect responses were followed by negative social feedback (thumbs down). In both tasks, we measured a rapid brain signal called the Reward Positivity, which occurs within a few hundred milliseconds after feedback and reflects how strongly the brain reacts to rewards. Narcissism was unrelated to brain responses to money. In the social task, however, higher narcissism predicted a weaker brain response to positive feedback. This held for both admiration (self-promotion) and rivalry (competitiveness). In other words, rather than showing heightened sensitivity to praise, narcissistic individuals were less engaged with it. These findings support the idea that narcissists disengage from social rewards. Such disengagement may help them maintain an image of self-sufficiency, but it also carries costs. Tuning out praise could make it harder to learn from feedback, strengthen bonds, or benefit from supportive relationships. By showing that narcissism is linked to reduced early brain responsiveness to social rewards, our study helps explain how narcissistic self-views are sustained and why interpersonal difficulties often arise.
Narcissism, characterized by egocentric exceptionalism and social selfishness (Sedikides, 2021), has long fascinated the public and experts alike. Perfectionism is a defining feature of narcissism, shaping how narcissists think, feel, and relate to others (Smith et al., 2018). Perfectionistic strivings mean that even subtle cues from others can carry weight, as these others affirm or challenge the flawless image narcissists strive to maintain. Yet psychologists disagree on what such strivings reveal. Some argue that narcissists appear confident but are inwardly insecure, leading them to seek and react strongly to social rewards such as praise and approval (Bushman & Baumeister, 1998; Stucke & Sporer, 2002). Others contend that narcissists do not necessarily seek out or react strongly to social rewards. Instead, they may downplay or even dismiss even positive feedback, adopting a self-sufficient and detached stance (Fernie et al., 2016; Zhu & Chen, 2015). This stance may not reflect genuine self-certainty but rather serve a protective function. By disengaging from social input, narcissists may assert superiority and stabilize their self-image, using denial or behavioral withdrawal to reduce reliance on others and guard against shame or self-threat (Fernie et al., 2016; Morf & Rhodewalt, 2001). Furthermore, their sense of entitlement may generate chronically high expectations, rendering positive feedback less surprising and thus less rewarding.
These conflicting views raise a key question: do narcissists exhibit heightened sensitivity to socially rewarding feedback or do they disengage from it? To address this question, we draw on the Narcissistic Admiration and Rivalry Concept (NARC) model (Back et al., 2013). This model distinguishes between a self-enhancement pathway, expressed as admiration, and a self-protective pathway, expressed as rivalry. Whereas admiration could imply heightened sensitivity to rewarding social feedback, rivalry instead predicts defensive devaluation and disengagement. Leveraging this distinction, we examine early neural responses to positive social feedback, indexed by the Reward Positivity (RewP), to test competing hypotheses of elevated reward sensitivity versus early-stage disengagement.
Although narcissism has been the subject of nearly 12,000 articles, its neural basis remains remarkably understudied. As noted in recent studies (Jauk & Kanske, 2021; Zhou et al., 2025), a survey of the literature identified 53 neuroscientific studies on narcissism. Although the field continues to grow, this figure illustrates the scarcity of neural relative to behavioral research (approximately 0.45%). Furthermore, the majority of these 53 articles examine grandiose narcissism, which is characterized by an extraverted, exhibitionistic, self-assured, dominant, and manipulative interpersonal orientation (Roberts et al., 2018; Thomaes et al., 2018). Methodologically, many of these studies have examined peripheral physiological responses, such as autonomic or neuroendocrine activity, collected at baseline or during laboratory tasks (Cheng et al., 2013; Noser, 2017; Pfattheicher, 2016; Reinhard et al., 2012; Wardecker et al., 2018) rather than direct neural activity. Indeed, only a small fraction of studies has linked narcissism to functional neural activity during experimental tasks, particularly those involving feedback or reward processing. Therefore, we focus our review on this subset of studies. Within this limited body of work, research using fMRI has yielded two neuroanatomical insights. First, higher levels of narcissism are associated with increased anterior cingulate cortex (ACC) activity in response to one’s own face (Jauk et al., 2017) and to experiences of social exclusion (Cascio et al., 2015). This heightened ACC response may explain why narcissism is associated with greater aggression following social exclusion (Chester & DeWall, 2016). Second, narcissism is associated with reduced activity in the anterior insula (AI) during tasks involving empathy for others (Fan et al., 2011) and anticipated interpersonal touch (Scalabrini et al., 2017). This explains why narcissists are low on empathy (Hepper et al., 2014) and need for intimacy (Emmons, 1989).
Both the ACC (Alexander & Brown, 2019; Botvinick, 2007; Gasquoine, 2013; Hyman et al., 2017) and AI (Corlett et al., 2022; Hoy et al., 2023; Ullsperger et al., 2010) are central to how people process feedback, detect errors, and evaluate outcomes. These regions help to track discrepancies between expected and actual results, assign meaning to feedback, and adjust behavior accordingly. Given that narcissism is marked by perfectionistic strivings (Smith et al., 2016), self-protective motivation (Grijalva & Zhang, 2016), and heightened sensitivity to evaluations (Hart et al., 2021; Horton & Sedikides, 2009; Kernis & Sun, 1994), the functions of the ACC and AI are especially relevant. As those high in narcissism show distinct ACC and AI activity during self-relevant and socially evaluative experiences (Jauk & Kanske, 2021; Zhou et al., 2025), these regions offer a promising window into how they process performance-related feedback in real time.
One neural signal that reflects these regions is the Error-Related Negativity (ERN; Gehring et al., 1993). The ERN is a negative deflection in the EEG waveform that peaks approximately 50 ms after an error and is reliably elicited during speeded reaction time tasks involving both social and non-social stimuli (Suzuki et al., 2020). It is interpreted as a marker of cognitive control, triggered when one’s behavior violates task demands and adjustment is needed. The ERN has been closely linked to the ACC (Carter et al., 1998; Debener et al., 2005; Dehaene et al., 1994; Edwards et al., 2012; Holroyd et al., 1998; Holroyd et al., 2004; Iannaccone et al., 2015; Keil et al., 2010; Kiehl et al., 2000; Mathewson et al., 2005; Menon et al., 2001; Miltner et al., 2003; Ullsperger & von Cramon, 2001; Veen & Carter, 2002) and AI (Bastin et al., 2016; Menon et al., 2001; Suárez-Pellicioni et al., 2013; Wessel et al., 2012), and is often used to index error sensitivity and conflict monitoring. Although the ERN has been extensively studied in relation to anxiety (Saunders & Inzlicht, 2020) and perfectionism (Barke et al., 2017; Drizinsky et al., 2016; Meyer & Wissemann, 2020; Perrone-McGovern et al., 2017; Schrijvers et al., 2010; Stahl et al., 2015), it has rarely been examined in the context of narcissism.
One of the few studies to address narcissism in conjunction with the ERN broke down grandiose narcissism into admiration and rivalry (Mück et al., 2023). Admiration is defined by the pursuit and maintenance of a haughty self-view through assertive self-promotion, whereas rivalry is defined by the protection of a haughty self-view through antagonizing or derogating others (Back et al., 2013). Participants first completed a standard measure of admiration and rivalry narcissism (Back et al., 2013), and then engaged in a speeded reaction time task across two blocks. During the second block, they received normative feedback suggesting that they had performed worse than their peers. Under this self-threatening condition, rivalry was associated with a more negative ERN, whereas admiration was unrelated to ERN amplitude. Hence, rivalry narcissism, being especially sensitive to self-threat, may involve heightened cognitive control when one’s self-image is challenged.
Several limitations, however, temper this interpretation. The Mück et al. (2023) study was likely underpowered, and small samples are known to inflate effect sizes and reduce replicability (Button et al., 2013; Lin, 2018). Moreover, measuring narcissism before task performance raises the question of whether narcissistic self-views were primed, potentially influencing participants’ neural responses (Nordlund, 2009; Weingarten et al., 2016). Given that trait priming can alter the active self-concept (Skowronski et al., 2010; Wheeler et al., 2014), it remains to be determined whether the observed neural blunting represents a stable, enduring pattern that persists even when self-schemas are not explicitly activated prior to the task.
Addressing these methodological questions, Robins et al. (2025) used a post-task measurement approach. In Study 1 (N = 144), participants completed the Eriksen Flanker Task while their neural responses to errors were recorded via EEG. In Study 2 (N = 50), participants completed a modified version of the Flanker Task that included trial-by-trial feedback. In both studies, participants then filled out the same measure of admiration and rivalry narcissism after the EEG recording. Notably, despite this difference in measurement timing, the results converged with Mück et al.’s (2023) findings regarding the antagonistic dimension: higher rivalry narcissism was associated with a blunted (i.e., less negative) ERN. These associations remained significant when controlling for number of errors and were supported by an internal meta-analysis. Such a cross-study consistency, observed across both pre-task (Mück et al.) and post-task (Robins et al.) designs, indicates that the neural disengagement in rivalry is a robust phenomenon, representing a genuine deficit in error sensitivity rather than a transient artifact of measurement priming.
We next turn to a related neural signal that reflects how individuals respond to external feedback, the RewP. It is a positive-going event-related potential that typically peaks 200 to 300 ms after feedback in reward-related tasks (Proudfit, 2015). The RewP is thought to reflect a reward prediction error, driven in part by activity in the ACC (Gehring & Willoughby, 2002; Hauser et al., 2014; Miltner et al., 1997; Ruchsow et al., 2002; Warren et al., 2015; Zhou et al., 2010) and AI (Becker et al., 2014; Billeke et al., 2020; Hauser et al., 2014), occurring when outcomes exceed expectations (Cavanagh & Holroyd, 2025). Crucially, however, beyond prediction errors, the RewP also indexes the fundamental neural sensitivity to reward outcomes (Proudfit, 2015). Thus, a blunted RewP can reflect either a failure of prediction (e.g., “I fully expected this”) or a deficit in reward sensitivity (e.g., “I do not value this”). Only one study has examined the association between grandiose narcissism (assessed with the Narcissistic Personality Inventory; Raskin & Terry, 1988) and the RewP. In this study (Yang et al., 2018), participants completed a monetary gambling task involving high- and low-risk trials while EEG was recorded. Participants high in grandiose narcissism did not differ in RewP amplitude from those low in narcissism. Grandiose narcissism may modulate the RewP, but the study was likely underpowered to detect such a relation. This issue is compounded because the researchers manipulated an additional independent variable (i.e., outcome magnitude) along with feedback. It is also possible that any aberrations in narcissists’ feedback processing, whether elevated or blunted, may not emerge in monetary tasks but may instead be specific to socially evaluative contexts.
Here, we examined how distinct dimensions of narcissism relate to feedback-related neural responses, focusing specifically on the RewP in the context of both monetary and social rewards. Building on evidence that narcissists show blunted neural responses to their own errors, we tested whether they also exhibit reduced neural sensitivity to external feedback. Drawing on the NARC model (Back et al., 2013), we propose distinct pathways: admiration narcissists may evince heightened responses to social rewards (Sheldon et al., 2020), driven by their desire for adulation (Back et al., 2013), dominance (McGregor et al., 2023), and visibility (Nevicka et al., 2018). Alternatively, rivalry narcissists may downplay or dismiss socially rewarding feedback. Crucially, rather than reflecting mere self-sufficiency, we posit the reflection of a defensive disengagement to protect the self from dependence (Morf & Rhodewalt, 2001). Such a pattern would be consistent with previously reported ERN findings (Robins et al., 2025). Furthermore, the high temporal resolution of ERPs allows us to move beyond a simple binary distinction between heightened sensitivity and disengagement by examining the time course of reward processing. Theoretically, narcissists might exhibit heightened initial sensitivity followed by a later, automatic attenuation. However, the RewP (occurring ∼250–300 ms) specifically indexes the early, relatively automatic assignment of incentive salience (Proudfit, 2015), distinguishing it from subsequent and more controlled regulatory processes. Therefore, focusing on the RewP allows us to determine whether the proposed disengagement operates as a late-stage tendency or as a pre-emptive, automatized gating mechanism at the stage of initial valuation. These competing possibilities raise a deeper theoretical question: do narcissists remain highly sensitive to feedback or do they disengage from it altogether?
Method
Participants
We recruited 99 University of Southampton undergraduates and excluded one due to a neurological condition, resulting in a final sample of 98 (M = 19.10 years, SD = 1.40; 80.6% female). All participants provided written informed consent at the beginning of the study and received course credit upon completion. We conducted a power analysis in G*Power (version 3.1.9.2; Faul et al., 2009) based on the effect size from Robins et al. (2025, Study 2 1 ). This analysis indicated that a minimum of 84 participants would be needed to detect a medium effect size (r = .30) with 80% power at an alpha level of .05. Our final sample exceeded the minimum required by our power analysis, ensuring adequate statistical power for the planned analyses. The study was approved by the Psychology Ethics Committee of the University of [MASKED; Protocol Number 68828).
Tasks
Participants first completed the Monetary Incentive Delay (MID) task and the Social Incentive Delay (SID) task, adapted from a previous ERP study (Ait Oumeziane et al., 2017). We counterbalanced the order of the two tasks across participants, with half engaging in the MID task first and the other half engaging in the SID task first.
In the MID task, participants completed trials marked by one of two cues, each signaling a different reward contingency: a circle containing a pound symbol (£) represented the monetary incentive condition (potential gain or loss; n = 50), whereas an empty circle denoted the neutral, no-incentive condition (certain break-even; n = 20). Each cue was followed by a variable anticipation interval (2000–2500 ms) during which a fixation cross (“+”) was displayed. Subsequently, a white box target appeared, prompting participants to quickly press the spacebar. After the target disappeared, the fixation cross reappeared for 1300 ms during the feedback anticipation period. For incentive trials, correct responses (button press during the target display) resulted in a green upward arrow (“↑”), indicating a £0.32 gain, whereas incorrect responses (button press outside the target window) led to a red downward arrow (“↓”), indicating a £0.16 loss. In neutral trials, participants always received break-even feedback (£0). We displayed feedback for 2000 ms, followed by a 1000 ms inter-trial interval. We dynamically adjusted task difficulty to maintain ∼50% accuracy, increasing target duration by 10 ms after errors and decreasing it by 10 ms after correct responses. We used an initial practice block of 10 trials (8 incentive and 2 neutral) to calibrate task difficulty. Midway through the task, we gave participants a short break and feedback on their cumulative earnings.
The SID task mirrored the MID task in overall structure, particularly regarding stimulus timing and sequence, but differed in its cues and feedback. Each trial was signaled by one of two cues marking the reward contingency: a blue circle that contained an “f” (resembling the Facebook logo) represented the social incentive condition (potential positive or negative social evaluation; n = 50), whereas an empty circle indicated the neutral, no-incentive condition (no social evaluation; n = 20). Each cue was followed by a variable anticipation interval (2000–2500 ms) during which a fixation cross (“+”) was displayed. A social evaluation target then appeared, prompting participants to press the spacebar as quickly as possible. After the target offset, the fixation cross reappeared for 1300 ms during the feedback anticipation period. In incentive trials, correct responses (i.e., button press during target presentation) resulted in a “thumbs up” icon, indicating positive social evaluation; incorrect responses (or responses outside the target window) produced a “thumbs down” icon, reflecting negative evaluation. Neutral trials consistently led to break-even feedback, displayed as an equal sign (“=”). We presented feedback for 2000 ms, followed by a 1000 ms inter-trial interval. We dynamically adjusted task difficulty to maintain ∼50% accuracy, increasing target duration by 10 ms after errors and decreasing it by 10 ms after correct responses. We used an initial practice block of 10 trials (8 incentive and 2 neutral) to calibrate task difficulty. Participants received a short break and feedback on their cumulative earnings halfway through the task.
Measurements
After the EEG tasks, participants completed the Narcissistic Admiration and Rivalry Questionnaire (Back et al., 2013), an 18-item self-report measure, to assess grandiose narcissism across two forms: Admiration (9 items, α = .89; e.g., “Being a very special person gives me a lot of strength”) and Rivalry (9 items, α = .84; e.g., “I can barely stand it if another person is at the center of events”). We also computed an overall narcissism score (α = .87). Participants responded on a scale ranging from 1 (not agree at all) to 6 (agree completely).
EEG Data Collection and Processing
We recorded continuous EEG data during task performance from 32 scalp sites using Ag/AgCl electrodes that were embedded in a flexible cap (Brain Products, UK) and positioned according to the international 10–20 system. We acquired the data via a Brain Vision actiCHamp Plus amplifier (Version: label 004 05/2015; Brain Products, UK) at a sampling rate of 500 Hz. We placed the reference electrode at Cz and the ground electrode at FPz (Zhou et al., 2025). We recorded vertical electrooculogram activity from an electrode positioned below the right eye and maintained electrode impedances below 10 kΩ throughout the recording. We conducted offline preprocessing using EEGLAB 2025.0 functions (Delorme & Makeig, 2004), implemented in MATLAB 2025. Prior to ERP computation, we reconstructed the Cz electrode and re-referenced the data to the average of all scalp electrodes. We band-pass filtered continuous data between 0.1 and 40 Hz. We excluded data segments with voltage deviations exceeding ±100 µV on any channel, including the vertical electrooculogram channel, from further analysis. We applied Independent Component Analysis to identify and correct artifacts related to eye blinks, eye movements, and other sources of noise (Delorme & Makeig, 2004). We automatically classified and rejected artifactual components using the ICLabel plugin (Pion-Tonachini et al., 2019), and we automatically removed components with a probability greater than 90% of being non-neural (e.g., ocular or muscle artifacts). For ERP analysis, we segmented EEG data into epochs from −200 to 3000 ms relative to feedback onset, with baseline correction applied using −200 to 0 ms pre-feedback interval. We corrected noisy or malfunctioning channels (FP2) using spherical spline interpolation (Perrin et al., 1989). A substantial number of trials remained in both the MID (M = 37.73, SD = 19.94) and SID (M = 36.74, SD = 20.09) tasks. We report the distribution of trial numbers across conditions in Supplemental Material (Table S1).
Consistent with previous research (Brown et al., 2022; Kujawa, 2024; Kujawa et al., 2018), we calculated the RewP as the mean amplitude within a time window of 250 to 300 ms post-feedback onset in the MID task and 275 to 325 ms in the SID task, following the parameters established in prior studies (Ait Oumeziane et al., 2017). To isolate reward-related neural activity, we computed the gain-minus-loss difference waveform (ΔRewP), defined as the RewP difference between gain and loss trials (Bao et al., 2025). Additionally, we calculated a regression-based residualized difference score (residual RewP), because recent evidence suggests that this approach may yield improved reliability compared to conventional subtraction-based difference scores (Brush et al., 2018; Meyer et al., 2017). We calculated the unstandardized positive residual RewP (indexing pure reward sensitivity) by regressing mean amplitudes of positive feedback trials on negative feedback trials. These unstandardized residuals represent neural activity specific to reward processing (in µV), with variance shared across conditions (e.g., low-level visual processing or general feedback evaluation) removed. Furthermore, to rigorously test the valence specificity of our effects, we computed the reciprocal unstandardized Negative Residual RewP by regressing negative feedback amplitudes on positive feedback amplitudes. This dual-index approach allowed us to determine whether narcissism is associated with a specific alteration in reward processing (Positive Residuals) or merely reflect a generalized blunting across all feedback valences.
Statistical Analyses
We conducted data analysis in SPSS version 29 (IBM Corp., Armonk, NY). First, we calculated descriptive statistics for demographic information and variables of interest. Subsequently, we carried out a 2 (task condition: incentive, neutral) ×2 (feedback type: positive, negative) repeated measures ANOVA to examine group-level effects on RewP. Finally, we conducted Pearson correlational analysis to investigate the association between narcissism and both ΔRewP and residual RewP.
Results
Description of Demographic Characteristics and Amplitudes of Reward Positivity (RewP) and Related Indices in Different Conditions
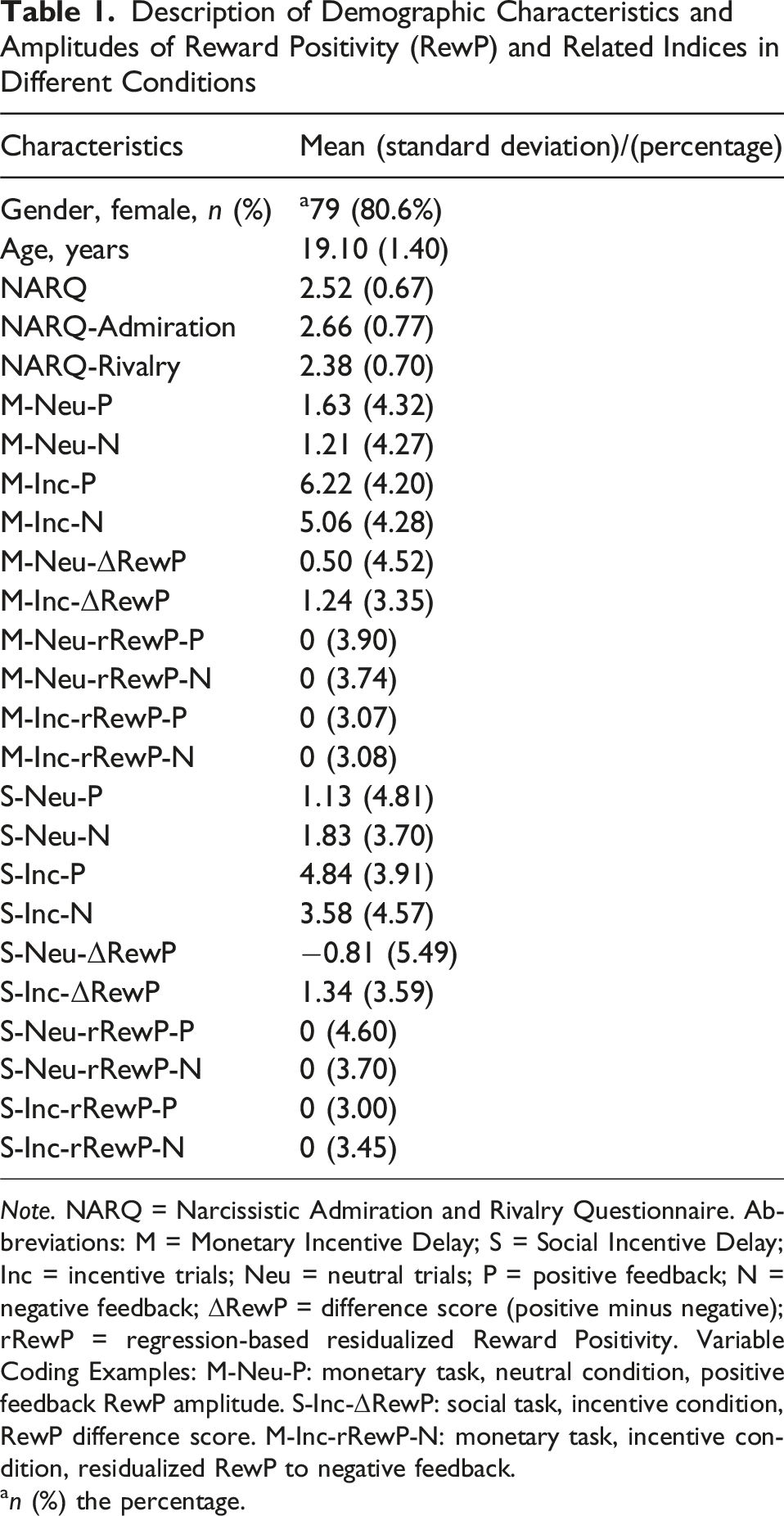
Note. NARQ = Narcissistic Admiration and Rivalry Questionnaire. Abbreviations: M = Monetary Incentive Delay; S = Social Incentive Delay; Inc = incentive trials; Neu = neutral trials; P = positive feedback; N = negative feedback; ΔRewP = difference score (positive minus negative); rRewP = regression-based residualized Reward Positivity. Variable Coding Examples: M-Neu-P: monetary task, neutral condition, positive feedback RewP amplitude. S-Inc-ΔRewP: social task, incentive condition, RewP difference score. M-Inc-rRewP-N: monetary task, incentive condition, residualized RewP to negative feedback.
an (%) the percentage.
ERP Analysis
We present the RewP and difference waveforms (ΔRewP) in Figure 1. For the MID task, a 2 (Trial type: incentive, neutral) × 2 (Feedback type: positive, negative) repeated measures Analysis of Variance (ANOVA) at the Cz electrode revealed a significant main effect of trial type such that RewP amplitudes were larger on incentive trials (M = 5.68 ± 0.44 μV) compared to the neutral trials (M = 1.45 ± 0.40 μV), F(1, 81) = 107.76, p < .001, η
p
2
= .57. We also observed a significant main effect of feedback type, such that RewP amplitudes were larger for positive (M = 3.96 ± 0.41 μV) than negative (M = 3.18 ± 0.40 μV) feedback, F(1, 81) = 5.22, p = .025, η
p
2
= .06. The interaction between trial type and feedback type was not significant, F(1, 81) = 1.41, p = .239, η
p
2
= .02. RewP waveforms (left), results of repeated measures ANOVA for RewP (middle), and ΔRewP scalp topographies (right) under incentive and neutral conditions in the MID (A) and SID (B) tasks
For the SID task, we carried out a repeated measures ANOVA at the Cz electrode. We observed a significant main effect of trial type such that RewP amplitudes were incentive trials (M = 4.22 ± 0.43 μV) compared to neutral trials (M = 1.55 ± 0.34 μV), F(1, 83) = 40.65, p < .001, η p 2 = .33. The main effect of feedback type was not significant, F(1,83) = 0.53, p = .469, η p 2 = .01. However, the interaction between task condition and feedback type was significant, F(1,83) = 8.33, p = .005, η p 2 = .09. We next examined the simple main effect of feedback type separately for incentive and neutral trials. On incentive trials, positive feedback (M = 4.86 ± 0.43 μV) elicited a larger RewP than negative feedback (M = 3.58 ± 0.50 μV), F(1, 83) = 12.34, p < .001, η p 2 = .13. On neutral trials, there was no difference in RewP amplitude between positive (M = 1.16 ± 0.51 μV) and negative (M = 1.93 ± 0.40 μV) feedback, F(1, 83) = 1.66, p = .202, η p 2 = .02. We present mean and standard deviations of trial numbers across conditions in Table S1.
RewP and Narcissism
We report bivariate correlations in Figure 2. In the MID task, the RewP from positive or negative feedback on incentive or neutral trials was unrelated to admiration, rivalry, or overall narcissism. In the SID task, the same was true for neutral trials and for negative feedback on incentive trials. However, positive feedback showed a small but non-significant negative association with overall narcissism (r = −.18, p = .084) and rivalry narcissism (r = −.24, p = .025), though the latter association was not significant under the Benjamini–Hochberg adjusted threshold of p = .017). Finally, the association with admiration narcissism was not significant, r = −.07, p = .489. Correlations of narcissism and indices of RewP in Monetary (lower diagonal) and Social (upper diagonal) Incentive Delay Task
Given the small associations in the SID task, we next sought to more precisely isolate reward-related neural activity. To do so, we used two approaches. First, we implemented a difference wave approach (Bao et al., 2025; Meyer et al., 2017), subtracting the RewP amplitude to negative feedback from the RewP amplitude to positive feedback (ΔRewP) for incentive trials separately for the MID and SID tasks. Evidence indicates that regression-based residual scores are a more reliable method of isolating reward-related neural activity than subtraction-based approaches (Brush et al., 2018). Consequently, we also computed a residual RewP for incentive trials separately for the MID and the SID. To create the residual RewP, we saved the unstandardized residual as a variable from a linear regression predicting RewP to positive feedback from RewP to negative feedback (Meyer et al., 2017). We did this separately for the MID and SID tasks and only used incentive trials.
The correlations between ΔRewP and overall narcissism, as well as its two forms, differed across the two tasks (Figure 2). In the MID task, ΔRewP was not significantly correlated with overall narcissism or its forms (ps > .05). In contrast, during the SID task, ΔRewP showed a significant negative correlation with overall narcissism (r = −.24, p = .023), with a similar (but not significant) association with admiration narcissism (r = −.20, p = .065) and rivalry narcissism (r = −.20, p = .066), even though these results did not survive the Benjamini–Hochberg false discovery rate correction for multiple comparisons. The residual RewP exhibited a similar pattern: In the MID task, the residual RewP was not significantly correlated with overall narcissism or its forms (ps > .05). In the SID task, residual RewP was significantly negatively correlated with overall narcissism (r = −.27, p = .011) and rivalry narcissism (r = −.27, p = .011). Both survived false discovery rate correction for multiple comparisons (Benjamini–Hochberg adjusted threshold = .013), but admiration narcissism did not (r = −.18, p = .101). We report the full set of correlation coefficients between narcissism and the indices of RewP in Supplemental Material (Table S2).
Sensitivity Analyses
We followed up on the bivariate correlations between narcissism and both ΔRewP and residual RewP with sensitivity analyses in G*Power (version 3.1.9.2; Faul et al., 2009). Based on our sample size (n = 98), 80% power, and α = .05, the study was sufficiently powered to detect effects as small as r = .278. Correlations between ΔRewP and overall narcissism (r = −.24, p = .023, Observed Power = 67%), admiration narcissism (r = −.24, p = .027, Observed Power = 67%), and rivalry narcissism (r = −.21, p = .053, Observed Power = 55%) fell below this threshold. Correlations between residual RewP and overall narcissism (r = −.27, p = .011, Observed Power = 77%), admiration narcissism (r = −.24, p = .022, Observed Power = 67%), and rivalry narcissism (r = −.27, p = .011, Observed Power = 77%) were stronger. The correlation between overall narcissism was just below the threshold and the association with rivalry narcissism was appropriately powered.
Discussion
We asked whether higher levels of narcissism are associated with sensitivity to feedback or disengagement from feedback. Using the MID and SID tasks, we examined how overall, admiration, and rivalry narcissism related to neural responses to monetary and social feedback. Narcissism (overall, admiration, and rivalry) was unrelated to RewP responses in the MID task but was associated with a blunted RewP to positive social feedback on incentive trials in the SID task. Crucially, our analysis revealed a specific dissociation. Admiration narcissism was unrelated to RewP responses, whereas rivalry narcissism was significantly associated with a blunted RewP to positive social feedback on incentive trials in the SID task. Rather than showing more intense responses to socially rewarding outcomes, individuals high in rivalry were less engaged with them. This pattern aligns with previous ERN findings (Mück et al., 2023; Robins et al., 2025) and indicates that, specifically within the antagonistic dimension of narcissism, individuals may downplay or dismiss interpersonal feedback to avoid reliance on others for self-evaluation.
The findings offer insight into the two competing theoretical accounts of narcissistic self-regulation: heightened sensitivity versus disengagement (Bushman & Baumeister, 1998; Fernie et al., 2016; Stucke & Sporer, 2002; Zhu & Chen, 2015). Our results bolster the dual-pathway perspective of the NARC model (Back et al., 2013). The specific negative association between rivalry narcissism and RewP in the SID task supports the defensive disengagement account for this dimension. Specifically, individuals high in rivalry downregulate their neural responses to socially rewarding feedback, potentially as a self-protective tendency to mitigate perceived self-threat (Fernie et al., 2016; Hepper et al., 2022; Zhu & Chen, 2015). For rivalry narcissism individuals, tuning out this feedback serves to protect the self from potential threat or shame (Morf & Rhodewalt, 2001). One might interpret such an attenuated response to social feedback as a sign of emotional stability or strength (i.e., a secure independence). Indeed, such stability is often a hallmark of admiration narcissism. However, our results show that the blunted RewP is specific to rivalry narcissism, a dimension characterized by neuroticism, hostility, and interpersonal fragility (Back et al., 2013). Thus, rather than reflecting a secure, self-sufficient stance, this neural blunting in rivalry likely represents a maladaptive defensive tendency to ward off potential social threats.
This dimension-specific finding helps resolve the apparent discrepancy with prior research. Classic accounts have consistently linked narcissism to stronger approach motivation (Foster & Trimm, 2008; Sedikides & Campbell, 2017) and reward-focused behaviors such as gambling (Lakey et al., 2008). Our null association between admiration narcissism and RewP does not support the prediction of heightened neural sensitivity to social reward, raising the possibility that such sensitivity may be specific to certain types of social reward. Consistent with this possibility, recent process models propose that narcissism is rooted in affective contingencies that are stronger for status pursuit than affiliation (Grapsas et al., 2022). Within this framework, individuals high in admiration may be responsive to status-relevant rewards such as respect or prestige, but not necessarily to affiliative signals such as the “thumbs-up” used in the present study. Accordingly, the absence of an association for admiration may reflect a lack of sensitivity to this specific type of social reward rather than evidence for generally heightened or intact reward processing.
Consistent with this framework, the blunted RewP in rivalry narcissism aligns with evidence that individuals high in rivalry exhibit decreased affective reactivity to affiliative reward (Grapsas et al., 2022). Given that the “thumbs-up” in our task is primarily an affiliative signal, the blunted RewP likely reflects reduced engagement with affiliative feedback. This interpretation is further supported by the distinction between glory and affiliation (Wallace & Baumeister, 2002), as rivalry-oriented individuals may not perceive generic positive feedback as a meaningful opportunity for status enhancement. Instead, given their amplified physiological sensitivity to negative evaluation (Harjunen et al., 2023), they may devalue affiliative rewards as a means of minimizing potential social threat. Crucially, this pattern is not readily explained by a reward prediction error account based on chronically high expectations (i.e., entitlement). If high expectations were driving the association, admiration narcissism (the more entitled dimension) would have shown stronger attenuation of the RewP. Instead, admiration narcissism was unrelated to neural responses, whereas rivalry narcissism, which is associated with cynicism and negative social expectations (Back et al., 2013), exhibited stronger attenuation of the RewP. This dissociation indicates that the blunting in rivalry is unlikely to reflect fulfilled expectations of praise and instead points to reduced sensitivity to affiliative reward. Beyond this reduced sensitivity, contemporary process models further suggest that narcissism reflects a prioritization of status pursuit over affiliation (Grapsas et al., 2022). From this perspective, affiliative signals may carry less motivational relevance for individuals high in rivalry, not only because they are defensively devalued but also because they are less aligned with status-related goals. These accounts are not mutually exclusive and may jointly contribute to the observed pattern.
The finding that rivalry narcissism is associated with a blunted RewP offers insight into the temporal dynamics of this defensiveness. Given that the RewP captures early and relatively automatic reward evaluation, this blunting suggests that the disengagement in rivalry is likely an automatized defensive bias as opposed to a late-stage and controlled suppression. For individuals high in rivalry, positive social cues are devalued at the stage of initial encoding, implying a pre-emptive motivational gate that filters out potential affiliative signals before they are consciously appraised. This conclusion aligns with the NARC model, which distinguishes the defensive motivation of rivalry from the agentic and reward-seeking motivation of admiration. By isolating the blunting effect to rivalry, our findings clarify why the heightened sensitivity account (Bushman & Baumeister, 1998; Stucke & Sporer, 2002) was not supported in this context; such sensitivity is theoretically characteristic of the admiration dimension. Ultimately, the specificity of our results suggests that blunting of neural responses is a predominant tendency unique to the antagonistic, defensive aspect of narcissism. The pattern of results, however, varied across operationalizations. For raw RewP, the association with rivalry was statistically significant at the nominal level but did not survive multiple-comparison correction. For ΔRewP, the effect did not reach conventional levels of statistical significance. In contrast, the association remained significant for the residualized RewP after multiple-comparison correction. One interpretation of this pattern is that the evidence for the rivalry effect is weak and sensitive to analytic specification. An alternative interpretation is that the residualized approach may provide a more precise index of reward-related variance, consistent with prior methodological work (Brush et al., 2018; Meyer et al., 2017). We therefore interpret these findings with appropriate caution.
In addition, the findings advance understanding of the motivational significance of the RewP. Although the RewP is an established neural marker of reward processing (Proudfit, 2015) and has been extensively studied in monetary and social incentive contexts (Ait Oumeziane et al., 2017; Pegg et al., 2021), it is also sensitive to approach-motivated states, such as anger (Angus et al., 2015; Threadgill & Gable, 2020; Tsypes et al., 2019). Greater effort similarly enhances the RewP, with higher cognitive or task demands eliciting larger responses (Bogdanov et al., 2022; Ma et al., 2014; Pan et al., 2023), particularly when rewards are perceived as effort-contingent (Harmon-Jones et al., 2024; Harmon-Jones, Clarke, et al., 2020; Harmon-Jones, Willoughby, et al., 2020). Building on this effort literature, self-control exertion also increases the RewP (Huang et al., 2025). Collectively, these findings imply that the RewP is shaped by diverse self-related motivational processes. Our research extends this motivational account by illustrating that self-control exertion, which reflects the executive regulation of the self, increases the RewP, whereas antagonistic self-protection, indexed by rivalry narcissism, is associated with a reduction in it. This perspective explains why rivalry narcissism predicts a lower RewP. To the extent that the RewP is modulated by self-regulatory effort, the results highlight the lack of effort that individuals high in rivalry invest in getting along with others (Back, 2018; Back et al., 2013), particularly when the benefits are socially contingent and require acknowledging external validation.
An attenuated early neural responsiveness to socially rewarding feedback could have real-world implications, particularly for individuals characterized by rivalry narcissism. Such attenuation may diminish the reinforcing value of praise (Brummelman et al., 2026), curtail the positive impact of social approval (Chen et al., 2024), and reduce the likelihood of sustaining prosocial behavior (Wu & Hong, 2022). Furthermore, by systematically tuning out positive social information, rivalry narcissists may fail to integrate signs of social acceptance into their self-concept. This defensive devaluation mechanism, while momentarily protecting their autonomy, likely reinforces interpersonal difficulties and vulnerability to social exclusion (Cascio et al., 2015), a core feature of the rivalry dimension. Future research could test these ideas by examining whether interventions that enhance sensitivity to positive social cues can improve social functioning and well-being among individuals high in narcissism.
Limitations
The interpretation of these findings should be considered in light of several limitations. First, the primary limitation of this study concerns statistical power and sample size, long-standing challenges in both neuroscience and personality-social psychology. In neuroscience, statistical power has been estimated to range between 8% and 31%, with substantial heterogeneity across subfields (Button et al., 2013; Nord et al., 2017). EEG and neuroimaging studies commonly use sample sizes under 30 (Marek et al., 2022), which severely restricts the stability and replicability of effects. Social and personality psychology have faced similar concerns. Only 25% of findings in social psychology were found to be reproducible in a large-scale effort (Open Science Collaboration, 2015), with accompanying updated effect size expectations and sample size recommendations (Lovakov & Agadullina, 2021; Schönbrodt & Perugini, 2013). For small-to-moderate effects, the norm in psychological research, samples of at least 250 participants are now recommended to ensure reliable estimation and replicability. Situated at the intersection of personality psychology and neuroscience, our sample size (N = 98) exceeds typical standards for EEG studies and met our prior power target. However, it still falls below the increasingly stringent thresholds for stabilizing individual differences in neural measures (DeYoung et al., 2022; Marek et al., 2022). Despite these constraints, our primary finding linking rivalry narcissism to blunted social reward sensitivity survived the false discovery date correction for multiple comparisons. Furthermore, several key associations, particularly those involving the methodologically superior residual RewP, approached acceptable statistical power. Taken together, these metrics suggest that the observed effects are robust and not merely artifacts of exploratory analysis, although future large-scale, pre-registered replications remain essential to further cement these mechanistic insights.
Second, our social feedback paradigm (i.e., the SID task) is necessarily more constrained than face-to-face social interactions. The task relies on symbolic, computer-mediated cues (e.g., thumbs-up icons), which may not capture every aspect of social evaluation as it unfolds in naturalistic settings. However, such symbolic signals are a central feature of contemporary digital social life, where “likes” function as established indicators of social value and approval (Lowe-Calverley & Grieve, 2018). Moreover, this paradigm is extensively used in social neuroscience to isolate trial-wise feedback processing with the temporal precision required for ERP research (Ait Oumeziane et al., 2017; Davis et al., 2023). Critically, admiration narcissism was not associated with RewP amplitudes in the SID task, indicating that these cues were sufficient to elicit robust reward-related neural responses in individuals motivated by social affirmation. The selective blunting observed for rivalry narcissism therefore is unlikely to reflect a general task limitation and instead points to a specific pattern of defensive disengagement from positive social feedback. Nonetheless, future work should examine whether similar dissociations emerge in more interactive or naturalistic social-evaluative contexts.
Third, the number of retained trials for the RewP analysis was on the lower end for optimal signal stability. This was a structural consequence of the adaptive paradigm, which dynamically adjusted difficulty to maintain ∼50% accuracy, thereby capping the maximum possible positive feedback trials at approximately 25 per condition. To preserve a representative sample size given these constraints, we did not apply a strict exclusion threshold based on trial counts. Nevertheless, the resulting data demonstrated sufficient psychometric properties to support our conclusions. Specifically, despite the modest number of trials per condition, the RewP evinced good internal consistency, with split-half reliability of r SB = .77 in incentive-positive trials and r SB = .64 in incentive-negative trials for both the MID and SID tasks. Furthermore, the observation of a theoretically meaningful dissociation where blunting was specific to rivalry narcissism and the SID task, along with surviving the false discovery rate correction, argues against the presence of random measurement noise or generalized signal degradation. Future research using this adaptive paradigm, however, should consider substantially increasing the total number of blocks to overcome this ceiling effect and further strengthen signal robustness.
Fourth, the generalizability of our findings is limited by the demographic characteristics of the sample, which consisted largely of female undergraduates (80.6%) from a single institution. Given established evidence of sex differences in both narcissism expression (Grijalva et al., 2015) and neural reward processing (e.g., RewP amplitudes; Spreckelmeyer et al., 2009), the current unequal gender ratio prevents us from examining potential sex-specific effects or interactions. Although preliminary analyses in our sample indicated no significant correlation between sex and narcissism, ruling out a systematic trait bias, it remains possible that the observed neural associations are specific to women or the developmental stage of young adulthood. Future investigations, employing large, gender-balanced, and diverse community samples, are necessary to determine whether these neurophysiological patterns extend to the broader population.
Fifth, the cross-sectional nature of the study precludes causal inferences. Although we interpret the blunted RewP as a potential mechanism maintaining the detached stance of rivalry narcissism, it is equally plausible that early-life neural hyposensitivity to social rewards predisposes individuals to develop a rivalry orientation (i.e., a “bottom-up” pathway). Alternatively, both the personality trait and the neural profile may stem from shared third variables, such as developmental history or temperament. Longitudinal designs are required to disentangle these directional pathways.
Finally, to advance biologically informed theories of narcissism, the field must move toward collaborative, large-scale efforts. These should involve harmonized protocols and pooled data across laboratories. EEG research has already begun to embrace this shift: the EEGManyPipelines Project included 168 analysis teams across 37 countries (Trübutschek et al., 2024), highlighting the feasibility and necessity of collaborative approaches to ensure robustness against analytical variability (Botvinik-Nezer & Wager, 2023). Similar multi-site initiatives are emerging to test the reproducibility of ERP effects across diverse settings and populations (Clayson et al., 2025; Paul et al., 2025; Strzelczyk et al., 2023). For personality neuroscience to thrive, such initiatives must become the norm, enabling the field to examine subtle brain–personality relations with sufficient power and generalizability.
Conclusion
This study provided preliminary neurophysiological evidence that rivalry narcissism is characterized by a selectively blunted immediate neural responses to socially, but not monetarily, rewarding feedback, as indexed by the RewP. The findings support the self-protective disengagement account over the heightened sensitivity account, suggesting that individuals high in rivalry may proactively attenuate the early processing of social signals to shield their self-image. Although the observed effects are typical for personality neuroscience, their functional implications warrant cautious interpretation. We proposed that the reduced early responsiveness may theoretically limit the reinforcing value of praise and impede the integration of affirming feedback into the self-concept, potentially contributing to the interpersonal difficulties often observed in narcissism. Future work will do well to prioritize replicating these patterns in larger samples and exploring whether enhancing early sensitivity to positive social cues can serve as a viable intervention for improving social functioning and well-being.
Supplemental Material
Supplemental Material - Tuned Out, Not Drawn In: Blunted Neural Sensitivity to Social Reward in Narcissism
Supplemental Material for Tuned Out, Not Drawn In: Blunted Neural Sensitivity to Social Reward in Narcissism by Zhiwei Zhou, Douglas J. Angus, Constantine Sedikides, and Nicholas J. Kelley in European Journal of Personality.
Footnotes
Author Note
This research was not pre-registered.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Open Science Statement
Supplemental Material
Supplemental material for this article is available online.
Note
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.