
Abstract
Positive world beliefs (e.g., believing in a safe world) are associated with higher well-being and more social connections. While these associations have often been interpreted as causal, little is known about their origins. Here, we utilized a large (N ∼ 9000) Swedish twin sample to investigate the role of familial confounding (genetic influences and family environmental effects influencing both primals and social well-being) and test whether associations are in line with a causal effect. Indeed, positive world beliefs were associated with higher well-being, reduced loneliness, and less social isolation. When adjusting for familial confounding, associations were substantially attenuated, but remained significant—consistent with a causal association. Bivariate twin models strengthened this finding, indicating that genetic effects explain approximately half of the covariation. When also accounting for personality differences in addition to familial confounding, a small—yet significant—association remained. These findings highlight that familial influences and personality differences explain a substantial part of the observed associations between primals and social well-being, while also lending support for a small potentially causal effect—though the directionality of this effect remains unknown. We discuss the implications of these findings for primals research and positive psychology.
Plain language summary
People who believe in a more positive world report higher well-being and more social connections. These relationships are often interpreted as causal, suggesting that more positive world beliefs lead to feeling better and having more close friends. Yet, our upbringing and our genetic predispositions could also play a role in these relationships—factors, which may influence both, how positive we see the world and how we report our social well-being. Here, we analyzed around 9000 Swedish twins to understand the nature of such relationships. Utilizing twins, we can test whether associations are due to pre-existing differences such as genetic effects or family environment, or whether they are rather in line with causality. We found people who have more positive world beliefs report higher well-being, less loneliness, and less social isolation. Once we controlled for genetic and family environmental effects, the strengths of these relationships diminished, with around half of the relationships being due to genetics. However, we found that twins with more positive world beliefs were still more socially connected and reported higher well-being compared to their identical co-twins, in line with a small causal effect. When we also considered existing personality differences, associations became even smaller, but remained significant. Together these findings suggest that genetic predispositions and personality traits, can explain a substantial part of the relationship between primals and social well-being, meaning that the same factors may largely influences both, how positive we experience the world and our social well-being. Simultaneously, the significant remaining associations after accounting for genetic and personality influences, suggest a potential causal effect between these two, though much smaller than previously reported. Notably, the present data do not allow any conclusions about the direction of causality, that is, whether our world beliefs impact our social well-being, or the other way around.
Introduction
It is well established that well-being, loneliness, and social isolation are linked to health outcomes as well as longevity overall (e.g., Abdellaoui et al., 2018; Cacioppo et al., 2010; Holt-Lunstad et al., 2015; Howell et al., 2007). While loneliness describes the subjective feeling of not having one’s needs for social connection met, social isolation objectively assesses the number of social contacts one has or is in frequent contact with (Perlman & Peplau, 1981, pp. 31–32). Well-being commonly represents life satisfaction and positive emotions and is multifaceted (Bartels, 2015; Bautista et al., 2023). Loneliness, social isolation, and well-being have all independently been linked to health outcomes, including depression (Abdellaoui et al., 2018; Cacioppo et al., 2010; Cairney et al., 2008; Domènech-Abella et al., 2019; Kim et al., 2022; Lim et al., 2016; Richard et al., 2017), anxiety (Cairney et al., 2008; Domènech-Abella et al., 2019; Kim et al., 2022; Lim et al., 2016), cardiovascular disease (Abdellaoui et al., 2018; Boehm & Kubzansky, 2012; Freak-Poli et al., 2021; Hakulinen et al., 2018; Kim et al., 2022; Valtorta et al., 2016), and strokes (Hakulinen et al., 2018; Valtorta et al., 2016). Furthermore, lonelier or socially isolated individuals have a decreased life expectancy (Holt-Lunstad et al., 2015). Lower mortality rates are associated with higher subjective well-being (Chida & Steptoe, 2008; Howell et al., 2007; Lawrence et al., 2015; Steptoe et al., 2015), even after controlling for familial confounding (Sadler et al., 2011). Feelings of loneliness and social isolation are highly prevalent (Berlingieri et al., 2023; O’Sullivan et al., 2021; Richard et al., 2017; Schroyen et al., 2023; Su et al., 2023; Surklaim et al., 2022), making them a huge burden for health and economy (Goldman et al., 2024; Meisters et al., 2021; Mihalopoulos et al., 2020). To reduce this burden, in 2024, the world health organization (WHO) launched a new international commission, making social connections a global health priority (see https://www.who.int/groups/commission-on-social-connection). This highlights the importance of increasing well-being and decreasing loneliness and social isolation to reduce related costs and to ensure better health and life outcomes.
Interestingly, a more positive outlook has been associated with higher well-being and more satisfaction with social connections (Clifton et al., 2019; Clifton & Yaden, 2021; Dzuka & Dalbert, 2006; Lemay et al., 2024; Poulin & Cohen Silver, 2008; Stahlmann et al., 2020; Syropoulos, 2020). Conversely, a pessimistic, less positive outlook on the world has been linked to mental health problems (e.g., Clifton et al., 2019; Clifton & Yaden, 2021; Isaacowitz & Seligman, 2001; Karhu et al., 2024; MacLeod et al., 1997; Miranda & Mennin, 2007). Individuals largely differ in their beliefs about the general character of the world. For instance, some believe the world to be a funny, meaningful or enticing place, while others do not. Recently, Clifton et al. (2019) introduced a novel concept called primal world beliefs or in short primals. They identified 22 hierarchically organized primal world beliefs with three secondary overarching primals (Safe, Enticing, and Alive) and one primary primal (Good), corresponding to the belief that the world is good overall. Individual differences in primals can be assessed with the Primals Inventory, which has shown good validity or reliability in comprehensive studies (Clifton et al., 2019; Clifton & Yaden, 2021). For example, primals have been shown to be substantially associated with life satisfaction, number of close friends and loneliness (e.g., for Good and well-being, r = .55; Clifton et al., 2019; Clifton & Yaden, 2021; Lemay et al., 2024; Stahlmann et al., 2020). As primals were found to be reliable (Clifton et al., 2019), relatively stable (Clifton et al., 2019; Ludwig et al., 2023), and unrelated to socio-economic variables (Kerry et al., 2023; Ludwig et al., 2023), they are argued to serve as “stable lenses through which we perceive the world” (Clifton, 2020), which causally impact well-being, loneliness, and social isolation (see also Poulin & Cohen Silver, 2008). Based on the premise of a causal relationship, the first efforts to manipulate primals in order to improve related health outcomes have emerged (Hämpke et al., 2024), with more on the way. However, given that past research relied mostly on cross-sectional designs which do not allow for causal inferences, more work focusing on the nature of the associations between primals and related traits is needed. Given that nearly all psychological phenotypes are heritable (Polderman et al., 2015), one potential confounder is genetics. Social isolation, well-being, loneliness and primals have all been shown to be between 21% and 50% heritable (Bartels, 2015; Boomsma et al., 2005; Matthews et al., 2016; Perizonius et al., 2024). The observed associations between these phenotypes, therefore, could be due to genetic confounding rather than causal effects. Similarly, common family environmental effects (such as our rearing environment) could influence both, the way we believe the world to be and factors related to well-being and social interactions. The study of twins is one of the most widely utilized approaches (Posthuma et al., 2003) to further our understanding of the true underlying nature of associations between different human traits in the population. Twin modeling is particularly useful in situations where randomized controlled trials are not feasible. Although twin designs cannot fully solve the problem of causal inference, nor establish a direction of effect, they can help strengthen such inferences or lend strong evidence against causality (Gonggrijp et al., 2023; McGue et al., 2010).
The aim of the present study was to test whether the associations between positive world beliefs and well-being, loneliness, or social isolation can be explained by familial (including genetic and common environmental) factors or if they are in line with a causal effect (i.e., associations remain even after accounting for familial confounding). To test this, we utilized a large genetically informative Swedish twin sample. After estimating the phenotypic associations, we examined if associations hold once familial confounding is controlled for, utilizing discordant monozygotic twin pairs (e.g., Gonggrijp et al., 2023; McGue et al., 2010). If the observed associations are consistent with a potential causal relationship, we expect the twins with more positive world beliefs to report higher well-being, less loneliness and less social isolation compared to their identical co-twins. To explore familial confounding further, we disentangled the genetic and environmental influences on the associations between primals and well-being, loneliness, and social isolation, respectively, using bivariate twin modeling (e.g., Neale & Maes, 2004). These analyses estimate the relative importance of genetic, common (e.g., rearing/family), and unique environmental effects on the aforementioned associations. If associations are fully explained by genetic and common environmental effects, a causal effect is implausible. Primals are moderately related to both neuroticism and extraversion (r = −.33–.35, Perizonius et al., 2024). Furthermore, well-being as well as loneliness are most strongly related to neuroticism, while social isolation is associated with extraversion (Buecker et al., 2020; Kokko et al., 2015). For these reasons, sensitivity analyses were run to ensure that the observed associations were not simply explained by underlying personality differences.
Methods
Sample
In 2022/23, 9538 adult Swedish twins (Mage = 50.29, SDage = 7.75) of the Study of Twin Adults: Genes and Environment (STAGE) cohort in the Swedish Twin Registry (STR; Lichtenstein et al., 2002, 2006; Magnusson et al., 2013; Zagai et al., 2019) participated. After excluding individuals with unspecified zygosity (N = 45; 0.47% exclusion rate), the final sample contained 2423 complete twin pairs: 684 monozygotic (MZ) females, 416 MZ males, 405 dizygotic (DZ) females, 288 DZ males, and 630 DZ opposite-sex twin pairs. Twin singletons (N = 4651) were included in the analyses to contribute to the estimation of means and variances. Overall, more females participated (56.6%).
The participants gave informed consent before participation. This study conforms to the Declaration of Helsinki and was approved by the Swedish Ethical Review Authority (Dnr 2020-02575, 2021-02014, 2022-00109-02).
Materials and measurements
Primal world beliefs
Participants’ primal world beliefs, that is, how they believe the general character of the world to be, were assessed using Swedish translations of the Primals Inventories. A variety of primal world beliefs have been characterized, including beliefs that the world is meaningful, funny, enticing, safe, or alive. The latter three secondary primals—Enticing, Safe, and Alive—are further gathered under the overarching primary primal of believing the world to be Good overall (see Figure 1). Here, these four primals were assessed using the Swedish version of the short 18-Item primals inventory (PI-18; Perizonius et al., 2024). Eleven tertiary primals of interest were additionally assessed (e.g., Meaningful, or Just; for a full list see Figure 1 and Supplemental Table 1). Items ranged from, for example, “Most things in the world are good” to reverse-scored items such as “On the whole, the world is a dangerous place.” In total, the Swedish Primals Inventory spans 62 items. Participants indicated how strongly they (dis)agreed with the respective world beliefs on a 6-point Likert scale, ranging from “strongly disagree” (1) to “strongly agree” (6). After accounting for reverse scored items, mean scores were calculated per participant with higher scores indicating stronger agreement. Reliabilities were moderate to high with Cronbach’s alphas ranging from 0.67 to 0.96 (for details regarding the translation procedure and individual items see Clifton et al., 2019; Clifton & Yaden, 2021; Perizonius et al., 2024). Primals measurement. Note. PI-18 stands for the short 18-item primals inventory. For more details regarding these measures see Clifton et al. (2019), Clifton and Yaden (2021), and Perizonius et al. (2024).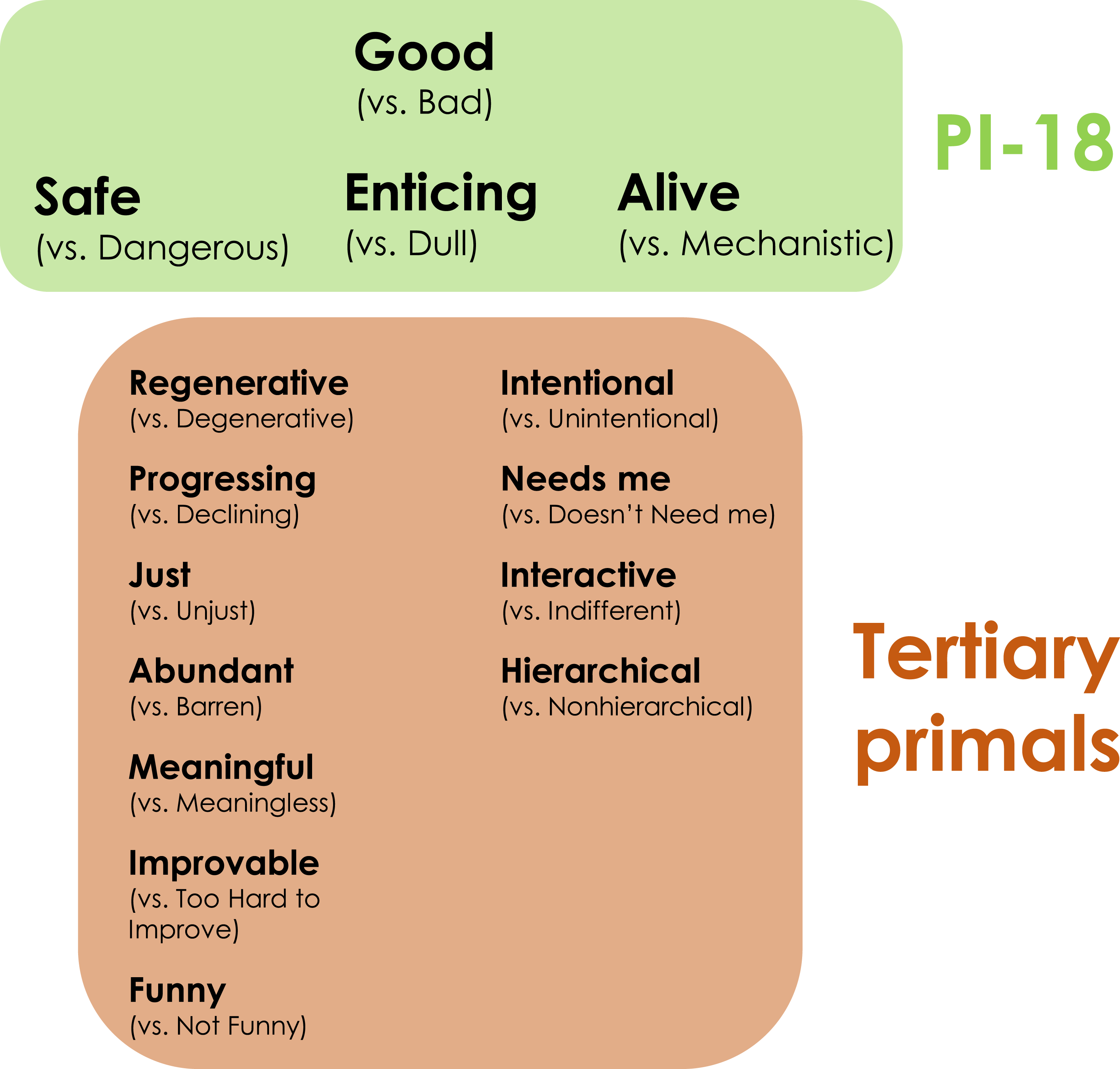
Well-being
Subjective well-being was assessed utilizing the Swedish translation of the unidimensional WHO 10-item well-being questionnaire (Löve et al., 2014). Participants were asked to indicate how they felt during the past week. Items included statements such as “I have felt calm and peaceful,” “I have lived the kind of life I wanted,” and “My daily life has been full of things that were interesting to me.” Answers were given on a 4-point Likert scale, ranging from “never” (0) to “all the time” (3). Sum scores were calculated for each participant, taking into account the first reverse scored item. A higher score corresponds to higher well-being. Cronbach’s alphas were high, with ratings ranging from 0.92 to 0.95 (Löve et al., 2014).
Loneliness
We assessed loneliness using the Swedish version of the UCLA loneliness scale (Hughes et al., 2004). This scale includes three questions asking how often one lacks companionship, feels left out or feels isolated from others. Answers were given on a 3-point Likert scale, ranging from “hardly ever” (1) to “often” (3). Sum scores were calculated per participant, with higher scores representing feeling lonelier (total sum scores ranged from three to nine). The reliability was moderate with a Cronbach’s alpha of 0.72.
Social isolation
Social isolation was assessed using an adapted version of the Lubben Social Network Scale (LSNS-6, Lubben & Gironda, 2003). The six questions address the number of friends or relatives, respectively, participants are (1) in contact with on a weekly basis, (2) feel they can confide in, or (3) can rely on for support (for more details see Supplemental Table 2). Answers were coded on a 6-point Likert scale with the response alternatives: “none” (5), “one” (4), “two” (3), “three to four” (2), “five through eight” (1), and “nine or more” (0). Sum scores were calculated, with higher total scores indicating more social isolation (ranging from zero to 30). Cronbach’s alpha was satisfactory at 0.80 and in line with past reports of reliabilities of 0.82–0.87 for the LSNS-6 (Jang et al., 2020).
Personality
Individual differences in neuroticism and extraversion were measured using a Swedish translation of the 30-item Big Five inventory (BFI-2-S; Soto & John, 2017a, 2017b; see also https://situationslab.squarespace.com/big-five-inventory-2, Supplemental Material 1 and Table 3). Scores were provided on a 5-point Likert scale ranging from 0 (“disagree strongly”) to 4 (“agree strongly”). The reliability was moderate with a Cronbach’s alpha of 0.67 for extraversion and 0.79 for neuroticism.
Data analyses
We standardized all measurement scores to ensure comparable effect sizes.
Phenotypic associations
We estimated associations between each of the 15 primals with well-being, loneliness, and social isolation, in the full sample, using multiple linear regressions. Clustering based on twin pair numbers was implemented to account for biased standard errors due to the relatedness of the sample. Sex and age have been found to be associated with all examined phenotypes (Barreto et al., 2021; Berlingieri et al., 2023; Cacioppo et al., 2006; Clifton et al., 2019; Cudjoe et al., 2020; Dahlberg et al., 2015; Dahlberg & McKee, 2014; Fugl-Meyer et al., 2002; Heikkinen & Kauppachin, 2011; Löve et al., 2014; Patulny & Bower, 2022; Steptoe et al., 2015; Su et al., 2023; Umberson et al., 2022; Yang & Victor, 2011), and were included in the analyses as covariates. Since p-value cut-off levels differ between phenotypic and genetic studies, multiple regression models were conducted here, and the sample was large, a stringent Bonferroni corrected significance level (correcting for multiple testing, significance level < 0.001) was chosen for all analyses (Figure 1). Nonetheless, the standard significance level of 0.05 and a stringent significance level of 0.01 (often used in genetic analyses of large samples) are also depicted for comparison in Figure 1 (see also Supplemental Tables).
Co-twin control design
To understand more about the underlying nature of the phenotypic associations observed in a cross-sectional design, co-twin control analyses can be conducted. The co-twin control design leverages on the fact that MZ twins have essentially identical genomes and a common environment of upbringing, if reared together (Gonggrijp et al., 2023; McGue et al., 2010). By investigating associations within pairs, we remove any part of the associations explained by influences shared between the twins of a pair. In other words, we estimate an effect free from familial confounding and as such closer to a potential true causal effect. Within-pair effect sizes and significance levels are compared to the associations observed in the full sample. In a scenario where causal effects between, for example, positive world beliefs and well-being, explain the full association, we would expect that the significance and effect size remain largely the same within MZ twin pairs compared to the full sample. This would suggest that no familial confounding is at play (for more details see Gonggrijp et al., 2023), that is, twins who are more positive than their identical co-twins also on average report higher well-being. If the relationship is fully confounded by familial influences, the association within MZ twins should be close to zero and non-significant. This would speak against a causal effect between the tested variables, since the observed associations were due to factors shared between twins of a pair. If the effect size of the association decreases compared to the effect size in the full sample, yet remains significant, partial familial confounding is suggested, while a small causal effect could still contribute to the association. Notably, while a co-twin control design can establish familial confounding and strengthen causal inferences, it can neither be used to infer the direction of causation nor to exclude unmeasured mediation effects (i.e., indirect causal effects).
Here, we assessed if associations remained significant after controlling for familial confounding for phenotypic significant (Bonferroni corrected) associations with absolute effect sizes larger than .10. This resulted in 34 within-pair linear regression models regressing well-being, loneliness or social isolation, respectively, on primals, clustering on exposure discordant MZ twins only (i.e., one twin believes in a more positive world than their respective co-twin). As MZ twins have the same age and sex, these variables are controlled for.
Bivariate twin modeling
To disentangle how much of the covariation between each primal and well-being, loneliness or social isolation is due to genetic or environmental effects, variance decomposition bivariate twin models were fit (Neale et al., 2006; Neale & Maes, 2004; Verhulst et al., 2019). Again, only phenotypic associations that had an absolute effect size larger than .10 and reached Bonferroni significance were assessed. We ran bivariate saturated models (SAT models) to test whether twin model assumptions were met (e.g., no mean and variance differences between twins of a pair; see Rijk & Sham, 2002). We expanded our bivariate SAT models to test age as a significant covariate and sex differences in the means, variances and/or covariances. Depending on SAT model results, either a simple model (no sex differences), a model with sex differences on the means or a model with sex differences on the covariances were fit (Neale et al., 2006). Covariances in these models are decomposed into A, C or D and E components. The genetic effects are reflected in the A (additive genetic) and D (dominance genetic/epistasis) components. The environmental effect is partitioned into two components, the common environmental component (C, e.g., rearing family environment) and the unique environmental component (E, which captures effects on twin pairs that make twins different from each other and measurement error; for more details see Neale & Maes, 2004). On the basis of cross-twin-cross-trait correlations, we estimated A, C or D and E influences on the covariance between primals and each outcome of interest. We, additionally, determined if the full ADE/ACE or more parsimonious AE, CE, or E models fitted the data best. Genetic and unique environmental correlations were estimated based on the best fitting model. Genetic correlations (rgenetic) capture how much of the underlying genetics are shared between the two phenotypes. How much the unique environmental influence on the two variables overlaps is captured in the environmental correlation (renvironment). To decide which model fitted best in the assumption testing and all of the above-mentioned analyses, structural equation model comparison utilizing chi-square tests with an alpha level of 0.01 were conducted.
Sensitivity analyses
We clustered people into lonely and non-lonely, following the questionnaire guidelines (Steptoe et al., 2013), due to loneliness not being normally distributed. This binary variable was used in a separately estimated liability threshold model to ensure that findings were not biased by non-normal distributions.
To ensure that the associations observed were not simply explained by underlying personality differences (Buecker et al., 2020; Kokko et al., 2015; Perizonius et al., 2024), we re-ran all analyses controlling for (i.e., regressing out) neuroticism in the analyses examining the associations between primal world beliefs and both well-being and loneliness, and for extraversion in the analysis of primal world beliefs and social isolation.
Transparency and openness
The present study and analyses were preregistered (https://aspredicted.org/r5hv-vvvj.pdf - note that the study proposed in the pre-registration has been split into two separate studies, with the present study focusing on the self-reported outcomes only, for example, see point 3, b (1)). As sensitive data were analyzed, data are only available through application (https://ki.se/en/research/swedish-twin-registry-for-researchers). All analyses were conducted in R (version 4.3.1; R Core Team, 2023). The following packages were used: psych (version 2.3.3; Revelle, 2023) and drgee (version 1.1.10; Zetterqvist & Sjölander, 2015) for the phenotypic and co-twin control analyses, OpenMx for twin modeling (version 2.20.6; Boker et al., 2023), tidyverse (version 2.0.0; Wickham et al., 2019) and dplyr (version 1.1.2; Wickham et al., 2023) for data handling, ggpubr (version 0.6.0; Kassambara, 2023) and corrplot (version 0.92; Wei & Simko, 2021) for visualization. All analyses scripts are accessible at https://osf.io/ayn72/overview.
Results
Phenotypic associations and co-twin control design
Phenotypically, believing the world to be good overall, that is, high scores on Good, was substantially positively associated with well-being, with an effect size of .41, and significantly negatively associated with both loneliness and social isolation, with effect sizes of −.22 and −.25, respectively. This pattern holds for 13 primals with well-being, 10 for loneliness and 11 for social isolation out of the 15 primals assessed here (for more details see Figure 2, orange lines, and Supplemental Tables 5, 7, and 9). Age and sex were significant in some associations, with females reporting significantly lower well-being and less social isolation, and older people reporting higher well-being and feeling less lonely (see Supplemental Tables 5, 7, and 9). Note that only (Bonferroni corrected) significant associations with absolute effect sizes larger than .10 were considered for forthcoming analyses, to meaningfully disentangle genetic and environmental influences (Figure 2). Phenotypic associations in the full sample and exposure discordant monozygotic (MZ) twins. Note. This figure displays the phenotypic associations in the full sample in blue. In orange, the associations adjusted for familial confounding using monozygotic (MZ) twins are displayed. Here, only the effect sizes that exceeded .10 in the full sample are shown. Notably, while most associations remained significant, the effect sizes dropped in MZ twins and confidence intervals widened. The tertiary primals Hierarchical and Interactive are not shown, as effect sizes of their associations in no case exceeded the .10 cut-off value. PI-18 stands for the short 18-item version of the primals inventory assessing only the primals Safe, Enticing, Alive, and Good. ns = non-significant, *p < .05, **p < .01, ***p < .001 (i.e., the last p-value corresponds to the Bonferroni multiple testing corrected p-value).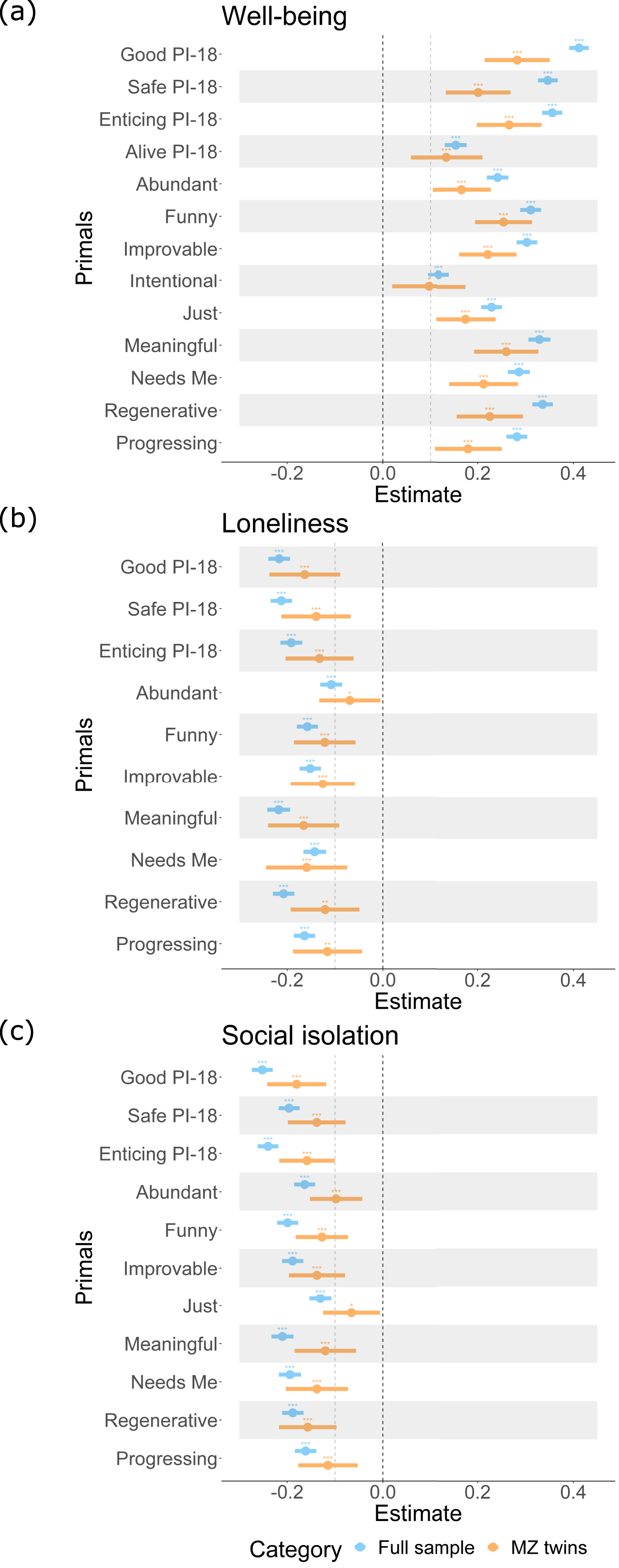
Utilizing the co-twin control design (see blue lines in Figure 2), we found that associations between most primals and well-being remained significant in MZ twin pairs, suggesting that the twins who had more positive world beliefs, also reported higher well-being than their respective identical co-twins. Similarly, for loneliness or social isolation, most associations remained significant in MZ twin pairs. All effect sizes, however, were reduced and confidence intervals widened due to smaller sample sizes (Figure 2). For more details regarding the co-twin control analyses see Supplemental Tables 6, 8, and 10.
Bivariate twin modeling
Bivariate SAT models revealed that age was a significant covariate in most models. Sex effects were observed on the means or variances, but not the covariances. Where indicated, age and/or sex were included as covariates in bivariate twin models. For associations that showed sex differences in the variances, sex differences were modeled on the variances as additional sensitivity analyses. Results did not differ from the ones treating sex as a covariate on the means (see Supplemental excel sheets).
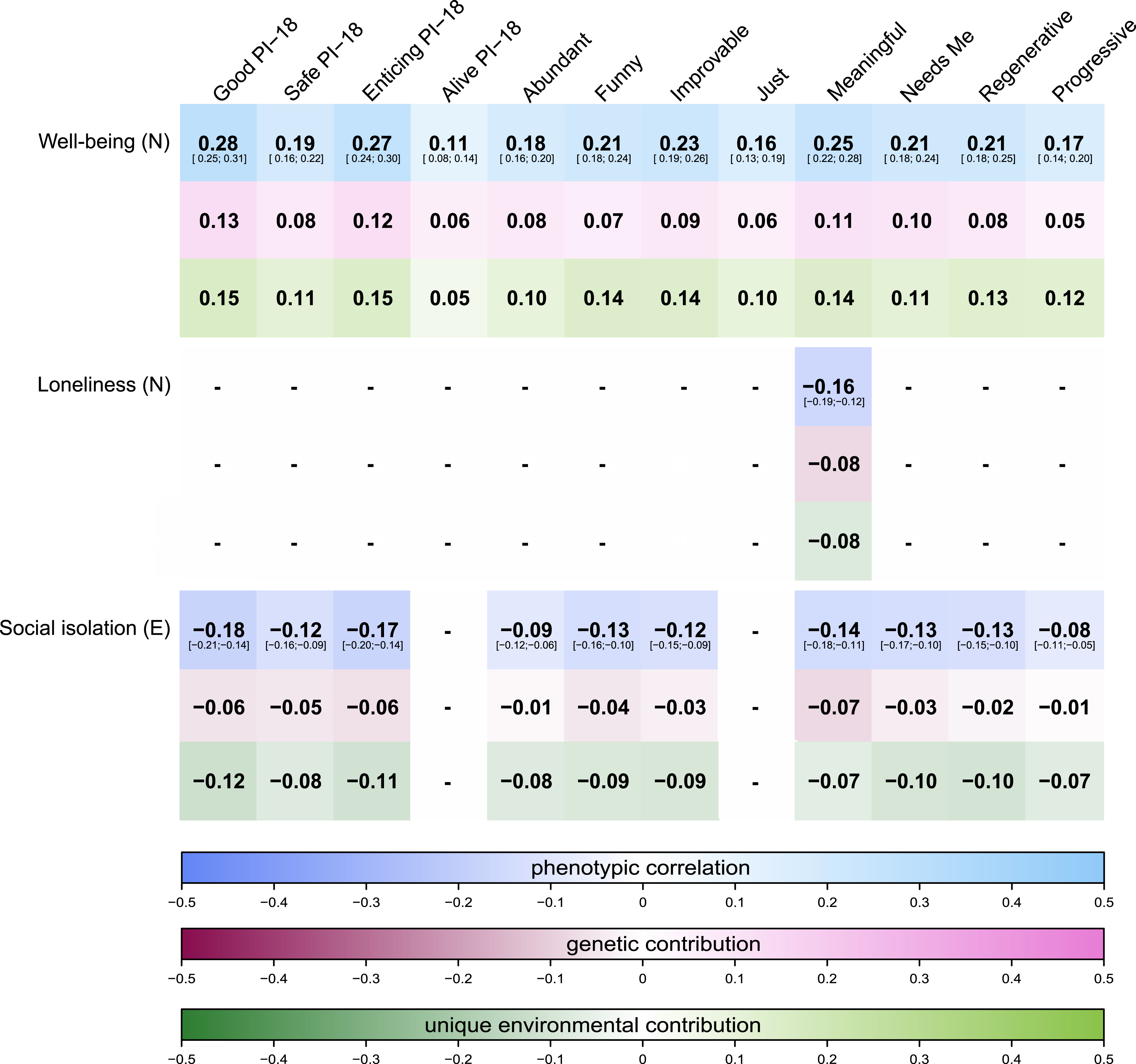
Bivariate twin modeling revealed that AE models fit the data best. For instance, the bivariate heritability for believing in a Good world and well-being was 53.8%, whereby bivariate heritability captures how much genetic influences contribute to the association between two traits. For loneliness and social isolation with Good, the bivariate heritabilities were 46.7% and 56.9%, respectively, with the rest of the variation being explained by unique environment and measurement error. The genetic correlation between, for example, Good and well-being was rgenetic = .60, while the correlation was negative for loneliness and social isolation and of slightly lower magnitude (rgenetic = −.31 and −.36). Similarly, the unique environmental correlation between Good and well-being was significant and positive (renvironment = .29), while slightly lower and negative for loneliness and social isolation (renvironment = −.17 and −.19). For the other primals, similarly approximately half of the phenotypic variation was explained by additive genetic effects. The proportions of the phenotypic association explained by genetic and unique environmental contributions can be observed in Figure 3. More bivariate analyses result details can be found in the Supplemental Materials and .excel sheets. Proportions of genetic and unique environmental contributions to the phenotypic associations. Note. Correlations are not displayed (−) when the associations did not survive the Bonferroni correction or did not exceed an absolute effect size of .10 in the full sample. Lighter colors reflect positive associations while darker colors reflect negative ones. Note, as here only complete twin pairs are used, the phenotypic effect sizes changed slightly in comparison to the full sample. For a comparison, 95% confidence intervals of the estimated phenotypic correlations are displayed underneath the point estimates, showing that although the phenotypic effect sizes change slightly, they are not significantly different from the estimates in the full sample. PI-18 stands for the short 18-item version of the primals inventory assessing only the primals Safe, Enticing, Alive, and Good.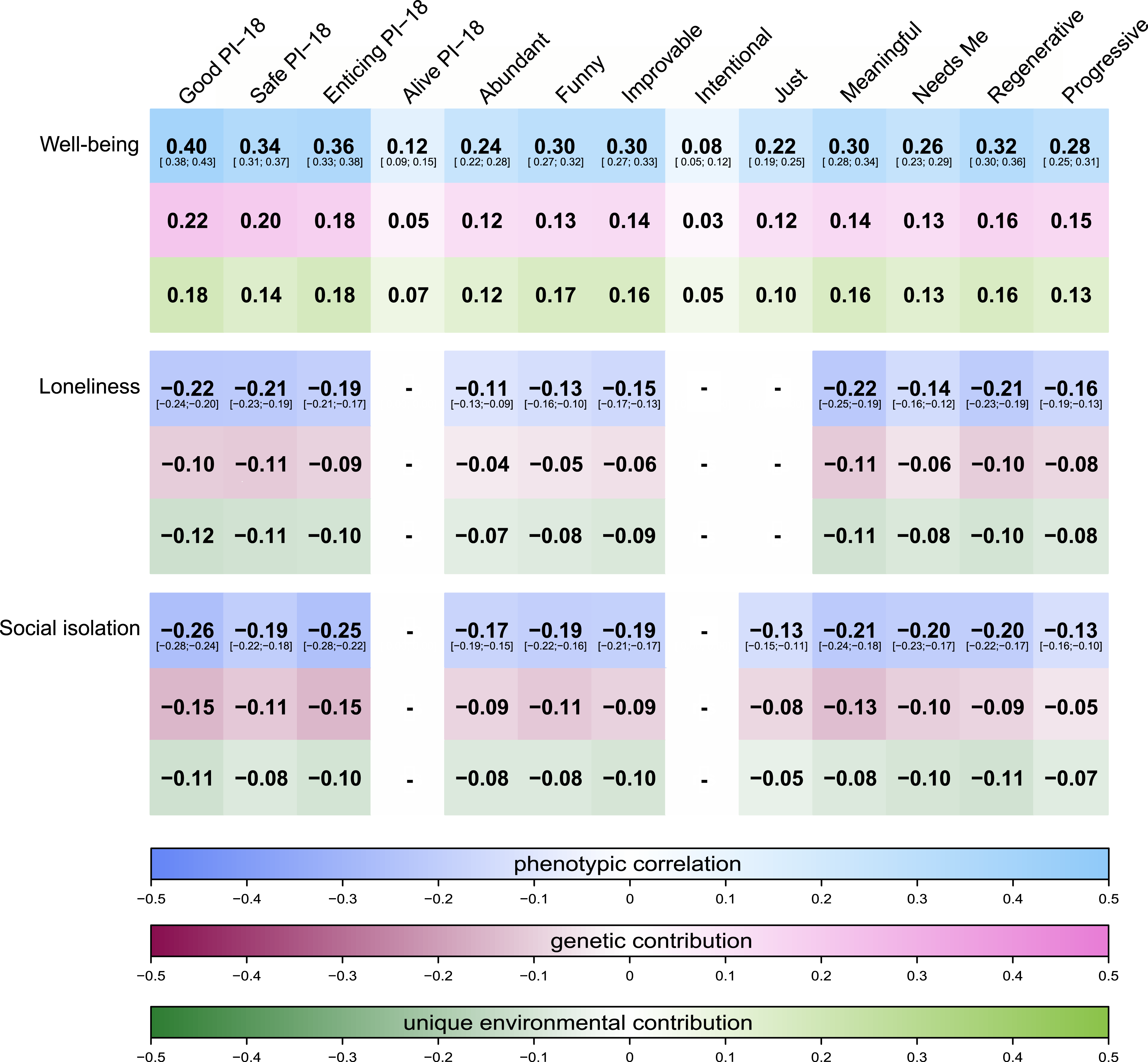
Sensitivity analyses
Treating loneliness as a binary variable showed very similar results (Supplemental .excel sheet). Neuroticism was moderately to highly correlated with well-being and loneliness, in the expected direction (r = −.61 95% CI [−.63; −.60], and r = .41 [.39; .43], respectively), while extraversion and social isolation were also moderately correlated (r = −.33 [−.35; −.31]). When adjusting the phenotypic associations for personality differences effect sizes were attenuated throughout (Figures 4 and 5). For loneliness, all phenotypic associations, except the association with Meaningful, diminished to effect sizes below .10. All other associations with well-being and social isolation were significant and most associations remained significant also within MZ twins (11 out of 13 and 8 out of 11, for well-being and social isolation, respectively; see Figures 4 and 5). Bivariate twin modeling further confirmed that at least half of the remaining associations were due to environmental influences (Supplement .excel sheets). Associations adjusted for personality differences (neuroticism or extraversion). Note. For well-being and loneliness, we adjusted for neuroticism, while for social isolation, extraversion was adjusted for. To compare these results to the ones where personality differences were not adjusted for, all previously significant associations that had an absolute effect size larger than .10 are displayed here. However, only these associations adjusting for personality that exceeded also .10 absolute effect sizes were considered for the bivariate twin modeling. PI-18 stands for the short 18-item version of the primals inventory assessing only the primals Safe, Enticing, Alive, and Good. ns = non-significant, *p < .05, **p < .01, ***p < .001 (i.e., the last p-value is the Bonferroni multiple testing corrected p-value). Proportions of genetic and unique environmental contributions to the phenotypic associations after adjusting personality differences. Note. Correlations are not displayed (−) when the associations did not survive the Bonferroni correction or did not exceed an absolute effect size of .10 in the full sample adjusting for personality differences. Lighter colors reflect positive associations while darker colors reflect negative ones. PI-18 stands for the short 18-item version of the primals inventory assessing only the primals Safe, Enticing, Alive, and Good.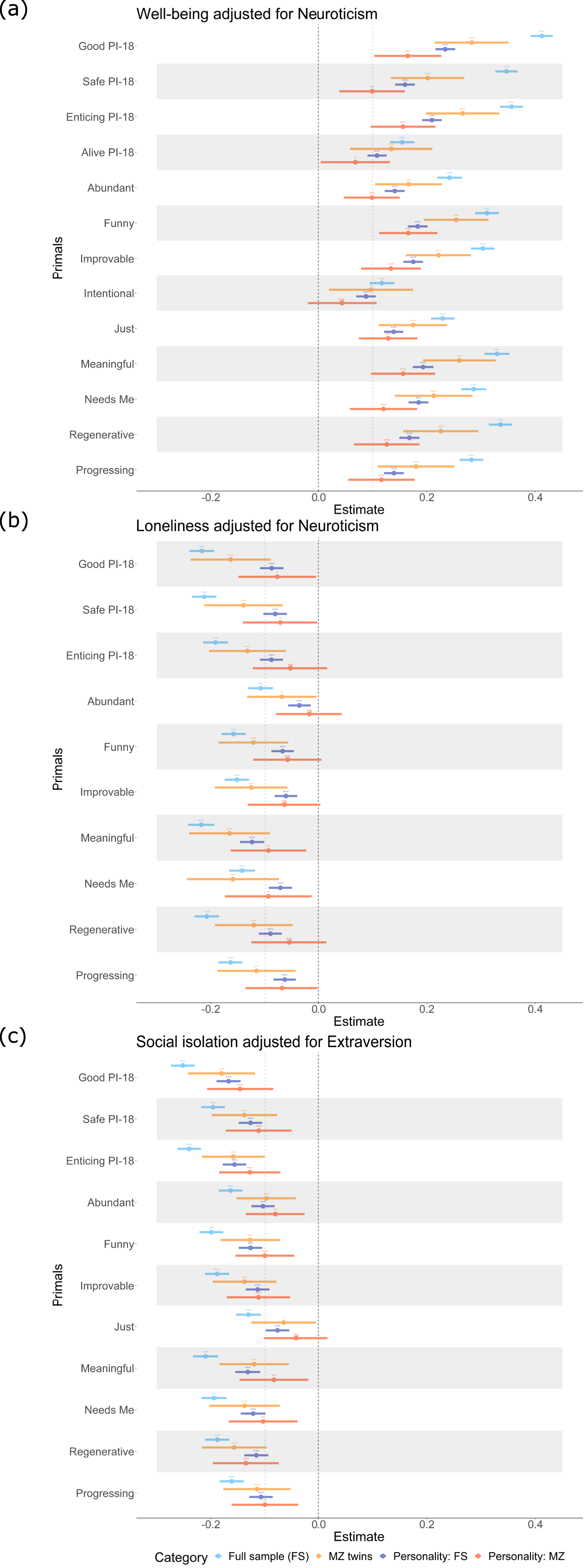
Discussion
Believing in a more positive world has consistently been associated with higher well-being, less loneliness, and less social isolation (Clifton et al., 2019; Clifton & Yaden, 2021; Lemay et al., 2024; Poulin & Cohen Silver, 2008; Stahlmann et al., 2020; Syropoulos, 2020). We replicated this pattern of findings in a Swedish population-representative twin sample showing that, indeed, those who saw the world more positively also consistently reported better social well-being. Utilizing twin data, we could show that effects sizes were substantially attenuated when adjusting for familial confounding, with around half of the associations being explained by shared genetic influences. Yet, most associations remained significant, in line with partly causal effects, even when personality differences in neuroticism or extraversion were also accounted for. In other words, our findings revealed robust associations between believing in a positive world and reporting higher well-being, less loneliness, and less social isolation, that could potentially reflect a causal effect. However, once genetic effects and personality differences were accounted for, these associations were small—much smaller than found on the population level, in the present study and in previous reports. It is important to note that our study design does neither allow for inferences about the directionality of these associations nor differentiates between direct and indirect causation (unmeasured mediation effects). The present findings highlight the importance of considering genetic confounding and individual differences in personality, when evaluating the relationship between positive world beliefs (primals) and social well-being.
Magnitudes of the relationships observed here differ from those previously reported, with Clifton et al. (2019) reporting slightly larger effects sizes for well-being (e.g., r = .55 for Good) and considerably larger effect sizes for loneliness (e.g., r = −.44 for Good), in a North American sample. The Alive world belief in particular related less to well-being and loneliness in our sample. This could be due to cultural differences. Notably, the effect sizes found by Stahlmann et al. (2020) in another European (German) sample were more similar to the ones found in this Swedish sample. Alternatively, it could be due to different measurements being used. Clifton et al. (2019) and Stahlmann et al. (2020) both assessed subjective life satisfaction to measure well-being. For loneliness and social connectivity, Clifton et al. (2019) used a single-item question. These measurements differ from the fine-grained multiple-item questionnaires utilized here. Future studies should explore whether these different phenotypic effect sizes are reflecting differences in the measurements or true cultural differences.
We showed that in MZ pairs the twin who believed the world to be more positive reported higher well-being, lower feelings of loneliness and less social isolation—the pattern of results we would expect if the associations were potentially causal. However, associations were attenuated compared to the full sample. Utilizing bivariate twin modeling revealed that approximately half of the covariation between the traits was explained by genetic confounding, further highlighting that the part that could be explained by a potential causal effect is substantially smaller—approximately half—than what would be expected by just exploring phenotypic relations. Nevertheless, the unique environmental estimates—the parts which could be attributable to causal effects (direct or indirect)—also include measurement error (Neale & Maes, 2004; Rijsdijk & Sham, 2002), and reverse causation cannot be ruled out (Gonggrijp et al., 2023; McGue et al., 2010). It could equally be true that higher well-being, less loneliness, and less social isolation have a positive effect on world beliefs. Moreover, bi-directional effects of causality or a third unmeasured environmental variable causing the associations are plausible. For example, a person harboring negative world beliefs could be less motivated to engage in social situations and invest less in nurturing close relationships, leading to increased loneliness. This is supported by previous findings showing that negative world beliefs are indirectly associated (i.e., moderated by strong-tie relationship quality) with loneliness over time (Lemay et al., 2024). However, it is also plausible that people who are lonelier and participate less in social activities may develop more negative world beliefs, since their everyday life will be less stimulating. To address the question of causal directionality, longitudinal—ideally, longitudinal genetically informative—samples are needed (e.g., Castro-de-Araujo et al., 2023; De Moor et al., 2008; Heath et al., 1993; Minică et al., 2018).
Expectedly, we found that genetic influences explain a large part of the covariance in above associations in line with previous research demonstrating that all examined phenotypes are considerably heritable (Bartels, 2015; Boomsma et al., 2005; Matthews et al., 2016; Nes & Røysamb, 2015; Perizonius et al., 2024; Plomin et al., 2016; Polderman et al., 2015). The significant genetic correlations between primals and well-being, loneliness or social isolation entail that partly similar biological mechanisms play a role in these phenotypes. To investigate the nature of such underlying genetic structures further, molecular genetically informative studies are needed (Uffelmann et al., 2021). Finding that genetic effects influence not only the phenotypes themselves but also their associations highlights that future studies analyzing the relationship of positive world beliefs with other psychological phenotypes need to consider the potential confounding of genetic influences.
Sensitivity analyses confirmed that being more neurotic leads to lower well-being, more feelings of loneliness, and a less positive worldview (Buecker et al., 2020; Clifton et al., 2019; Kokko et al., 2015; Perizonius et al., 2024; Stahlmann et al., 2020). Scoring higher on extraversion was associated with less social isolation, and more positive world beliefs, replicating previous results (Buecker et al., 2020; Clifton et al., 2019; Perizonius et al., 2024; Stahlmann et al., 2020). Controlling for these personality differences showed that part of the phenotypical associations between primals and well-being, loneliness or social isolation is explained by these differences. Also genetically, personality differences explained part of the covariation, as demonstrated by diminished bivariate heritabilities and lower genetic correlations when adjusting for neuroticism or extraversion. For loneliness, only believing the world to be Meaningful remained associated with feeling less lonely when adjusting for genetic confounding and neuroticism. This aligns with previous research showing that feelings of loneliness and neuroticism are phenotypically and genetically closely related (Abdellaoui et al., 2018; Buecker et al., 2020; Schermer & Martin, 2019). While the effect sizes largely diminished within MZ twin pairs for well-being and social isolation, most associations remained evident. Bivariate twin modeling revealed that even after controlling for personality differences the covariations between primals and well-being, loneliness or social isolation were partly genetically explained, leaving the remaining unique environmental contribution, albeit significant, relatively small.
Although the present findings do not rule out a causal effect of primals on the respective outcomes, they highlight that such an effect would be much smaller than previously anticipated. This, in combination with the uncertainty of the directionality of causal effects, raises the question whether interventions targeting primals to increase well-being and social connectivity would be an effective strategy to improve life outcomes. While primals intervention studies are still in their infancy, a recent study showed that temporarily changing one’s Enticing world belief was possible (Hämpke et al., 2024). Yet, receiving the primals intervention had no significant direct effect on well-being and the effect of changing one’s Enticing world belief was non-existent two weeks after the intervention. Thus, future work is essential that shows why targeting primal world beliefs specifically is preferable to directly targeting any given outcome to ensure better life outcomes.
Limitations of the present study should be noted. Attenuated effect sizes in co-twin control analyses might reflect measurement error in the exposure (Frisell et al., 2012; Gonggrijp et al., 2023; Gustavson et al., 2024; McGue et al., 2010). We accounted for this by having a relatively large twin sample, by ensuring reasonable reliability of the exposure and moderate MZ twin correlations in the exposure (Frisell et al., 2012; Gustavson et al., 2024; Perizonius et al., 2024). We further addressed this issue by estimating bivariate heritabilities and genetic correlations. The lack of significant D and C effects, moreover, could be due to a lack of power (Keller et al., 2010; Posthuma & Boomsma, 2000) or due to the fact that the influence of C decreases with age (Bergen et al., 2007). Another limitation is using self-rated and translated, sometimes adapted and/or different, questionnaires than the ones used previously. Not only do self-rated questionnaires introduce rater biases (e.g., Podsakoff et al., 2003), but different measurements in different cultures can lead to different outcomes. The lower effects sizes reported here could be due to this. Although we used long to moderately long versions for most questionnaires, we used a short version to assess personality differences, which could lead to lower effect sizes. While it should be noted that Clifton et al. (2019) used a longer personality questionnaire and still found, similar to us, only small overlaps with primals, longer versions of questionnaires could be advantageous, as they provide a more representative measure of a personality dimension, by comprehensively covering all its sub-facets (Soto & John, 2017b). Similarly, while the WHO 10 items well-being scale is robust and measures general well-being using multiple items, it does not allow for multidimensional testing of well-being (e.g., VanderWeele & Johnson, 2025; Zhang et al., 2024). Future research should explore if the present findings hold when longer, multidimensional measures are used. While our findings substantially contribute to our knowledge of the nature of the relationships between primals and loneliness, well-being as well as social isolation, future studies are needed to replicate the present findings.
In summary, three main conclusions can be drawn from the present study: Firstly and most importantly, approximately half of the phenotypic associations of positive world beliefs with higher well-being, lower feelings of loneliness, and less social isolation are due to genetic confounding. Secondly, associations are substantially explained by differences in the personality traits neuroticism and extraversion. Lastly, most associations between primals and social well-being remained within MZ twin pairs even after controlling for personality differences, showing a potential for causal effects. However, these effects are much smaller than reported in the past once genetic confounding was adjusted for. Altogether, this study underscores the importance of considering genetics and personality in future research on primals and related traits.
Supplemental material
Supplemental Material—How positive beliefs about the world relate to well-being and social connections—A genetically informative approach
Supplemental Material for How positive beliefs about the world relate to well-being and social connections—A genetically informative approach by Sophie Perizonius, Laura W Wesseldijk, Fredrik Ullén, and Miriam A Mosing in European Journal of Personality
Footnotes
Acknowledgments
We thank all Swedish twins that participated and the Swedish Twin Registry for providing the data.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The project was partly supported by Templeton Religion Trust (TRT0293).
Open science statement
The present study and analyses were preregistered (https://aspredicted.org/r5hv-vvvj.pdf—note that the study proposed in the pre-registration has been split into two separate studies, with the present study focusing on the self-reported outcomes only, for example, see point 3, b (1)). As sensitive data were analyzed, data are only available through application (https://ki.se/en/research/swedish-twin-registry-for-researchers). All analyses scripts and further supplemental material are accessible at ![]() .
.
Ethical considerations
This study conforms to the Declaration of Helsinki and was approved by the Swedish Ethical Review Authority (Dnr 2020-02575, 2021-02014, 2022-00109-02).
Consent to participate
The participants gave informed consent before participation and participant are aware that they can withdraw from the registry at any time. See also ethical approval above.
Supplemental material
Supplemental material for this article is available online.