
Abstract
ID2 is a rhythmically expressed helix–loop–helix transcriptional repressor, and its deletion results in abnormal properties of photoentrainment. By examining parametric and nonparametric models of entrainment, we have started to explore the mechanism underlying this circadian phenotype. Id2-/- mice were exposed to differing photoperiods, and the phase angle of entrainment under short days was delayed 2 h as compared with controls. When exposed to long durations of continuous light, enhanced entrainment responses were observed after a delay of the clock but not with phase advances. However, the magnitude of phase shifts was not different in Id2-/- mice tested in constant darkness using a discrete pulse of saturating light. No differences were observed in the speed of clock resetting when challenged by a series of discrete pulses interspaced by varying time intervals. A photic phase-response curve was constructed, although no genotypic differences were observed. Although phase shifts produced by discrete saturating light pulses at CT16 were similar, treatment with a subsaturating pulse revealed a ~2-fold increase in the magnitude of the Id2-/- shift. A corresponding elevation of light-induced per1 expression was observed in the Id2-/- suprachiasmatic nucleus (SCN). To test whether the phenotype is based on a sensitivity change at the level of the retina, pupil constriction responses were measured. No differences were observed in responses or in retinal histology, suggesting that the phenotype occurs downstream of the retina and retinal hypothalamic tract. To test whether the phenotype is due to a reduced amplitude of state variables of the clock, the expression of clock genes per1 and per2 was assessed in vivo and in SCN tissue explants. Amplitude, phase, and period length were normal in Id2-/- mice. These findings suggest that ID2 contributes to a photoregulatory mechanism at the level of the SCN central pacemaker through control of the photic induction of negative elements of the clock.
Keywords
Many aspects of biochemistry, physiology, and behavior are organized around a 24-h rhythm, driven by an endogenous circadian clock (Dunlap et al., 2004). Interlocked autoregulatory molecular transcriptional–translational feedback loops (TTFLs) underlie circadian organization in single cells. The positive loop is composed of basic helix–loop–helix (HLH)-PER-ARNT-SIM (bHLH-PAS) transcription factors CLOCK, NPAS2 and BMAL1; the negative loop of the period (per1, per2) and cryptochrome genes; and an interlocking loop of REVERB and ROR nuclear receptors (Papazyan et al., 2016). The master circadian oscillator in mammals is located within the hypothalamic suprachiasmatic nucleus (SCN), although many body tissues (e.g., heart, liver), as well as cell lines derived from them, harbor a circadian clock (Dunlap et al., 2004; Peirson et al., 2006; Dibner et al., 2010). Genomic analysis of one such line of mammalian fibroblasts identified a series of rhythmically expressed genes with a circadian periodicity, including Inhibitor of DNA binding 2 (Id2; Duffield et al., 2002). ID2 is an HLH transcriptional inhibitor and although involved in regulation of development (Wang and Baker, 2015), it has not been studied extensively in the context of the adult animal.
Our previous studies have revealed important roles for ID2 in the circadian system. First, ID2 interacts with CLOCK and BMAL1, core components of the clock mechanism, through their HLH domain and inhibits their transactivation potential by sequestering CLOCK and BMAL1 to the cytoplasm (Duffield et al., 2009; Ward et al., 2010). Second, the expression profile of clock-controlled genes in the liver of Id2-/-mice is altered, highlighting a role for ID2 in regulating the output pathways of the circadian clock (Hou et al., 2009). Recently, we revealed alterations in the nocturnal locomotor activity and feeding behavior profiles of Id2-/- mice (Mathew et al., 2013; Zhou et al., 2014a; Zhou et al., 2016).
The Id2 gene is expressed widely throughout the adult body and, when examined over 24 h, is rhythmically expressed at both the mRNA and protein levels (Duffield et al., 2002; Panda et al., 2002; Duffield et al., 2009; Hou et al., 2009; Hughes et al., 2009; Adachi et al., 2013). This 24-h rhythmic expression pattern includes the SCN, liver, and immortalized fibroblasts. Interestingly, Id2-/- fibroblasts show a ~2-fold higher per1 gene induction following serum stimulation as compared with stimulated wild-type (WT) cells (Ward et al., 2010), suggesting that ID2 may modulate the entrainment mechanism of the clock in peripheral tissues as well as in the master circadian clock in the SCN. In line with this hypothesis, Id2-/- mice show abnormally rapid entrainment in response to a large change in the photoschedule, corresponding increased magnitude of light-induced phase delays, and a delayed phase angle of entrainment (Duffield et al., 2009).
Although the putative role of Id2 and other members of the Id gene family in circadian entrainment has been explored (Duffield et al., 2009; Hou et al., 2009; Ward et al., 2010), the mechanism by which Id gene products affect photoentrainment remains unclear. An important feature of endogenous circadian clocks is that they require minor adjustments on a daily basis that ensure their correct phase relationship with the environment, a process known as entrainment. The daily alteration between light and dark is the major synchronizer, or zeitgeber, for the circadian clock in most organisms. The aim of our current study was to explore the mechanism by which these large light-induced phase shifts and rapid reentrainment are achieved in the Id2-/- mouse by examining parametric (continuous) and nonparametric (discrete) models of photoentrainment (Pittendrigh and Daan, 1976; Johnson et al., 2003; Dunlap et al., 2004). Parametric entrainment is achieved in response to exposure to continuous durations of light and is based on a tonic response of the clock to the luminance level, resulting in changes in the angular velocity of the clock and a change in the cycle length or circadian period (Aschoff, 1979; Winfree, 2001). Nonparametric entrainment is achieved in response to short-duration light pulses and is based on a circadian-dependent response to light—described by the phase-response curve (PRC)—resulting in discrete changes in the clock’s phase (Pittendrigh and Daan, 1976; Johnson et al., 2003; Dunlap et al., 2004).
In the current investigation, we used the Id2-null mouse to determine the extent to which the increased phase shifts and speed of entrainment in this animal model rely on parametric and/or nonparametric mechanisms. We examined the speed of resetting of the clock to light, as well as examined discrete light treatments of different durations and intensities. At the molecular level, we examined the induction of clock gene expression in response to a discrete light treatment. We also examined the anatomy of the retina and melanopsin-positive retinal ganglion cell (RGC) densities, pupillary responses to light treatments, and the amplitude of state variables of the clock as plausible explanations for the phenotype. The collective results suggest that the Id2-null mouse photoentrainment phenotype occurs during only the phase-delay portion of the PRC and is evident only under subsaturating light pulses. The phenotype is not explained by major differences in the shape of the PRC under saturating light pulses, in the speed of the resetting of the clock, in the anatomy and function of the retina or retinal pathways, or in the amplitude of state variables of the clock. Based on our prior studies (Duffield et al., 2009; Ward et al., 2010), we propose a model in which ID2 attenuates the circadian photic response by modulating signaling to the clock either at the gene promoter level or upstream of the intracellular signaling components that impinge on the period genes.
Materials and Methods
Animals
Id2-/- mice were generated and maintained as described previously (Duffield et al., 2009; Mathew et al., 2013). The Id2-null line of mice (129sv/C57BL6J/FBVN background) was separately crossed with the per1:luciferase (per1luc; CD1 background; Wilsbacher et al., 2002) as well as with the PER2::LUCIFERASE (PER2LUC; C57bl/6J background; Yoo et al., 2004) to allow for real-time bioluminescence reporting of gene and protein expression of mouse tissues, respectively. The per1:luc is a gene expression reporter construct, and the mouse line is generated from a multisite transgene approach. Firefly enzyme luciferase expression is driven by the mouse per1 promoter and 5′-UTR elements. Conversely, the PER2::LUC construct is a transgene fusion protein of the LUCIFERASE reporter to the terminal axon of the per2 locus, preserving proximal and distal regulatory elements, and where the transgene has replaced the native per2 gene using a targeted knock-in approach. Both transcriptional and posttranscriptional regulation of the PER2 protein is preserved, and as PER2 is translated in the presence of luciferin, its expression is monitored as bioluminescence. Mice were generated from in-house breeding at the University of Notre Dame (UND). Unless indicated otherwise, data shown were from experiments conducted on the Id2-null line. Age- and sex-matched littermate Id2+/+ mice were used as control mice in all procedures. An Id2+/- group was also included in the parametric entrainment experiments and in the CT14 saturating light pulse and multipulse phase-shifting experiments. Because no phenotype was identified in these or prior experiments (Duffield et al., 2009) for the Id2+/- group, an Id2+/- group was not included in several of the subsequent experiments. Food and water were available ad libitum. Animals were housed in a 12 h light:12 h dark (LD) regimen under climate-controlled conditions (19-21 °C, 50-65% humidity). All mice were entrained to the LD regimen for at least 3 weeks prior to experimentation. Experiments were conducted in accordance with the Institutional Animal Care and Use Committee at UND. The Id2-null mouse line was donated to the Mutant Mouse Resource & Research Centers and is available from Jackson Laboratory (Bar Harbor, ME; 41568-JAX; Id2tm1Mais/Mmjax).
Locomotor Activity Monitoring, Behavioral Manipulations, and Circadian Phenotype Analysis
Adult mice (≥3 months of age) were maintained in individual cages (29 × 11.5 × 13 cm) equipped with a running wheel (Actimetrics, Wilmette, IL) and a passive infrared movement detector (Slimline PIR motion detector, Smarthome, Irvine, CA). Mice were studied under a 12:12 LD cycle (150-400 lux, fluorescent lights, General Electric 36-W cool white), with lights on at 0700 h and off at 1900 h, or in constant DD (0 lux). All mice received sterile water treated with antibiotics (sulfamethoxalzole and trimethoprim oral suspension [Alpharma, Fort Lee, NJ] at a final dilution of 400 mg and 80 mg/L, respectively) and sterile food (see Duffield et al., 2009). Wheel-running and general activity, measured using a passive infrared motion detector, were monitored by a PC computer and using Clocklab hardware and software (Actimetrics). Actogram analyses were conducted on activity data using the Clocklab software, with data arranged into 6-min time bins. Lights-off was defined as zeitgeber time (ZT) 12. Circadian time (CT) 12 indicated activity onset under continuous darkness (DD) and was determined using standard methods (Duffield et al., 2009; von Gall et al., 1998; Duffield et al, 1998). Briefly, estimation of CT12 was determined by drawing a line by eye through at least 7 to 10 consecutive activity onsets on double-plotted actograms. Free-running periods were calculated from the slope of this fitted line. All lines were fitted by 2 individuals who were independent of the experimenter and who were blind to the treatments. In cases in which both wheel-running and general activity data were generated for an animal, the mean value was calculated. In most instances, there was little discrepancy between the 2-phase values; across experiments, the variability (range) between wheel-running and general activity values for each animal was 11 ± 2.1 min (mean ± SEM) or 6 min (median).
Continuous/Parametric Entrainment Experiments
In the reentrainment and the phase-delay experiments, the LD cycle was extended by 10 h so that lights-off changed from 1900h to 0500 h, and the lights-on in the following LD cycle changed from 0700 h to 1700 h. For the phase-advance reentrainment experiments, the light phase of the LD cycle was advanced by 6 h so that lights-on occurred earlier at 0100 h instead of 0700 h, and the lights-off in the following LD cycle changed from 1900 h to 1300 h. In the reentrainment experiments, the photoperiod before and after the shift remained as 12-h light and 12-h dark. The intensity of light during the extension of the light phase matched that of the preceding LD cycle and was 150 to 250 lux. Assessment of stable entrainment was determined from actograms and from profiles of activity/24 h. Phase shifts were determined by measuring the phase difference between eye-fitted lines through successive daily activity onsets, using observers blind to treatment.
Phase Angle Determination under Different Photoperiods
Mice were maintained on 12:12 (normal), 6:18 (short days), and 18:6 (long days) LD cycles, 30 days in each case. Using actograms, the phase angle of activity onset relative to ZT12 (lights-off) was calculated from 10 days of continuous activity in LD (the last 10-day segment in each respective photoperiod) and by drawing a line of best fit through 7 to 10 days of activity onsets.
Discrete/Nonparametric Entrainment and PRC Experiments
In the acute light pulse experiments, mice were entrained to a 12:12 LD cycle for 14 days and transferred to DD for ≥ 10 days of free-running activity. Mice were exposed to 30 min of light (800 lux) starting at CT12, CT14, CT16, CT18, CT20, CT22, CT0, CT4 and CT8 (after an initial 10 days in DD) and maintained for at least a further 10 days in DD. For the synthesis of the PRC, mice were reentrained to a 12:12 LD cycle before each subsequent DD light pulse treatment. The magnitude of the phase delays was determined by drawing a line of best fit through 7 to 10 days of activity onsets immediately before the light treatment and a second line through 7 to 10 days following the treatment, either immediately (for phase delays) or after the days showing transients (for phase advances). The phase shift was calculated as the difference between the actual onset of activity and that predicted by the pretreatment line on the last day of treatment. Data for CT4 and CT8 were combined for the purposes of statistical analysis. Experiments examining different intensities and durations of light (4 min at 10 lux, 1 min at 100 lux, 4 min at 100 lux, and 30 min at 800 lux) at CT16 were also undertaken in a similar manner (and after an initial 14 days in DD). Light intensities were established using neutral density filters (Lee Filters, Burbank, CA).
Cumulative Phase Shift Experiments
Mice were exposed to 1 of 4 treatment protocols before transition to DD conditions: (1) a 10-h delay of the LD cycle by a 10-h treatment with continuous light (250 lux) as described above or a series of discrete 15-min light pulses at 250 lux interspaced over 10 h by presenting light either (2) every 1 h, (3) 1.5 h, or (4) 2 h. The total accumulated light exposure during the night phase was (1) 10 h, (2) 2.75 h, (3) 2.25 h, and (4) 1.5 h, respectively. After treatment, mice were maintained in DD for 10 days, and the magnitude of the resultant phase delay was determined from actogram analysis as described above.
In Situ Hybridization
In situ hybridization (ISH) for per1 gene expression was conducted as previously described (de la Iglesia, 2007; Han et al., 2012) using a radiolabeled cRNA probe for per1 (nucleotide positions 538-1752; kindly provided by Dr. H. Okamura; Takumi et al., 1998). Autoradiographic images were generated by exposing slides to Ultramax film (Kodak, Rochester, NY). Images were scanned at high resolution, and hybridization intensities were determined with ImageJ software (National Institutes of Health, Bethesda, MD) using templates for the ventral SCN (for photic-induction analysis) or whole SCN (for rhythm analysis) (Han et al., 2012).
Retinal Histology and Antimelanopsin Immunohistochemistry
Animals were killed by cervical dislocation and eyes frozen and embedded in tissue tech on dry ice and stored in a −80 °C freezer prior to sectioning on a cryostat. Coronal sections (16 µm) were collected on adhesive-coated slides (Thermo Fisher Scientific, Pittsburgh, PA) and treated with 4% paraformaldehyde for 5 min and 2 washes in 1 × PBS. Slides were rehydrated in a descending series of alcohols, 100%, 95%, and 70%, for 3 min for each solution. Sections were briefly dipped in dd H2O and immersed in Cresyl Violet (Sigma-Aldrich, St. Louis, MO) for 10 to 20 min. Sections were dehydrated in an ascending series of alcohols, 70%, 95%, and 100%, for 30 sec each and cleared in xylene. Slides were coverslipped with DepexR (VWR International, West Chester, PA). Cresyl Violet (Nissl)–stained sections were viewed under brightfield microscopy and images digitally captured (Diagnostic Instruments, Sterling Heights, MI). Retinal layers were measured in micrometers and RCGs counted using SPOT software (Diagnostic Instruments, Sterling Heights, MI).
Melanopsin immunostaining in the retina was performed on alternate 16-µm retinal sections. Sectioned slides were washed in 1 × PBS for 20 min and incubated for 1 h in blocking buffer (1 × PBS/5% normal goat serum/0.3% Triton X-100/1% dimethyl sulfoxide). Antimelanopsin antisera (UF006; AB-N38, Advanced Targeting Systems, San Diego, CA) 1:5000 in blocking buffer (Provencio et al., 2002) was added and incubated overnight at 4 °C. The slides were washed 3× in 1 × PBS/0.1% TWEEN-20 for 10 min. Secondary antibody (Alexa Fluor 488 Invitrogen Goat anti-rabbit IgG) 1:500 in blocking buffer was applied for 1 h at room temperature, and the slides were washed as above. A final wash with 1 × PBS for 10 min was used, and the slides were mounted with Vectashield (Vector Laboratories, Burlingame, CA). Immunofluorescence images were collected on a Leica DM500B microscope. RGCs and antimelanopsin RGCs were counted.
Pupillometry
Pupil constriction response to 60-sec white light was assessed using the methods outlined in Van Gelder (2005) and Hattar et al. (2003). Mice were subjected to 1 min of light at ZT8.5-11 to test the response and magnitude of pupil constriction. Mice were dark-adapted for at least 2 h before the test. Treatments consisted of 10 lux and 100 lux of white light. The pupil responses of the mice were recorded on a handheld video camera and analyzed by measuring the pupil diameter in relation to cornea diameter. The pupil:cornea ratio was normalized to the dark-adapted aperture from time 0 sec to provide a measurement of relative pupil aperture (normalized pupil area = 1.0).
Real-time Gene and Protein Expression Analysis
per1luc or PER2LUC mice were sacrificed by cervical dislocation at 2 to 4 h into the light phase of the LD cycle (ZT 2-4). Brain tissue was quickly removed and placed in ice-cold Hanks Balanced Salt Solution (Cellgro, Corning, Tewksbury, MA). Coronal sections of the brain (300-µm thick) were cut on a manual vibroslice (NVSL, World Precision Instruments, Inc., Sarasota, FL). Brain regions were identified under a dissecting microscope, and the SCN was isolated as a square tissue (~1.5 mm2), containing either a rostral or caudal aspect of the SCN. Explants were cultured, using an established method, on a permeable membrane, creating an interface between culture medium and humidified air and housed within sealed culture dishes (Yamazaki and Takahashi, 2005; van der Veen et al., 2012; Zhou et al., 2014b). SCN tissues were cultured on culture plate inserts (Millicell, Millipore, MA), placed in individual 35-mm translucent Petri dishes with 1.2 mL of culture media composed of Dulbecco’s Modified Eagle’s Medium (Sigma-Aldrich), 19 mM D-glucose, 4.2 mM NaHCO3, 10 mM HEPES, 25 U/mL penicillin and streptomycin, 5% fetal bovine serum, and 100 mM luciferin (Luck-100, Gold Biotechnology, St. Louis, MO) and sealed with vacuum grease and glass coverslips (Wilsbacher et al., 2002; Yamazaki and Takahashi, 2005; van der Veen et al., 2012; Zhou et al., 2014b). Tissues were placed in a light-tight box at 36 °C, and bioluminescence was measured using a photomultiplier tube (LumiCycle, ActiMetrics, IL) every 10 min.
LumiCycle output data were analyzed for amplitude, period, and phase. The average amplitude and period length were calculated by fitting a sin damp curve to the length of data that displayed rhythms with the LumiCycle Analysis software (Actimetrics) for the first 3 rhythms after the initial peak. Phase was calculated by determining the initial peak in local time, converting the value to ZT time, and then converting to numerical values.
Results
Id2-/- Phase Angle Is Delayed under Variable Photoperiods
Previous work on the circadian system of Id2-null mice revealed a small difference in the phase angle of activity onset in the Id2-/- that was delayed relative to littermate control mice (Duffield et al., 2009; Mathew et al., 2013). According to the nonparametric model of entrainment, the phase angle of entrainment is the result of the daily phase shift that the circadian system needs to achieve to compensate for the difference between the endogenous period length (τ) and the period length of the zeitgeber (T), that is, 24.0 h. In turn, according to the parametric model of entrainment, τ changes predictably with exposure to light. In organisms that combine both models of entrainment, parametric and nonparametric, the phase angle of entrainment will depend on τ, which in turn will depend on the photoperiod length. We examined mice at different photoperiods, initially on a standard LD 12:12, then switching for 30 days to a short photoperiod of LD 6:18, and finally transitioning to a long photoperiod of LD 18:6 for 30 days (Fig. 1A). The phase angle of the activity rhythm was measured under the three different photoperiods and was consistently delayed in Id2-/- mice relative to the WT group (2-factor analysis of variance [ANOVA]: effect of photoperiod, F2,73 = 7.45, p < 0.01; effect of genotype, F1,73 = 9.34, p < 0.01; interaction F2,73 = 1.47, n.s.). Consistent with previous findings (Duffield et al., 2009; Mathew et al., 2013), Id2-/- mice exhibited a small delay in the phase angle under LD 12:12 relative to WT controls, in this case 20 min (post hoc test, p < 0.05). Under short photoperiods, the change in phase angle was especially large, being 2 h delayed relative to WT controls (p < 0.05; Fig. 1B). Under the long photoperiod, no significant difference was detected.
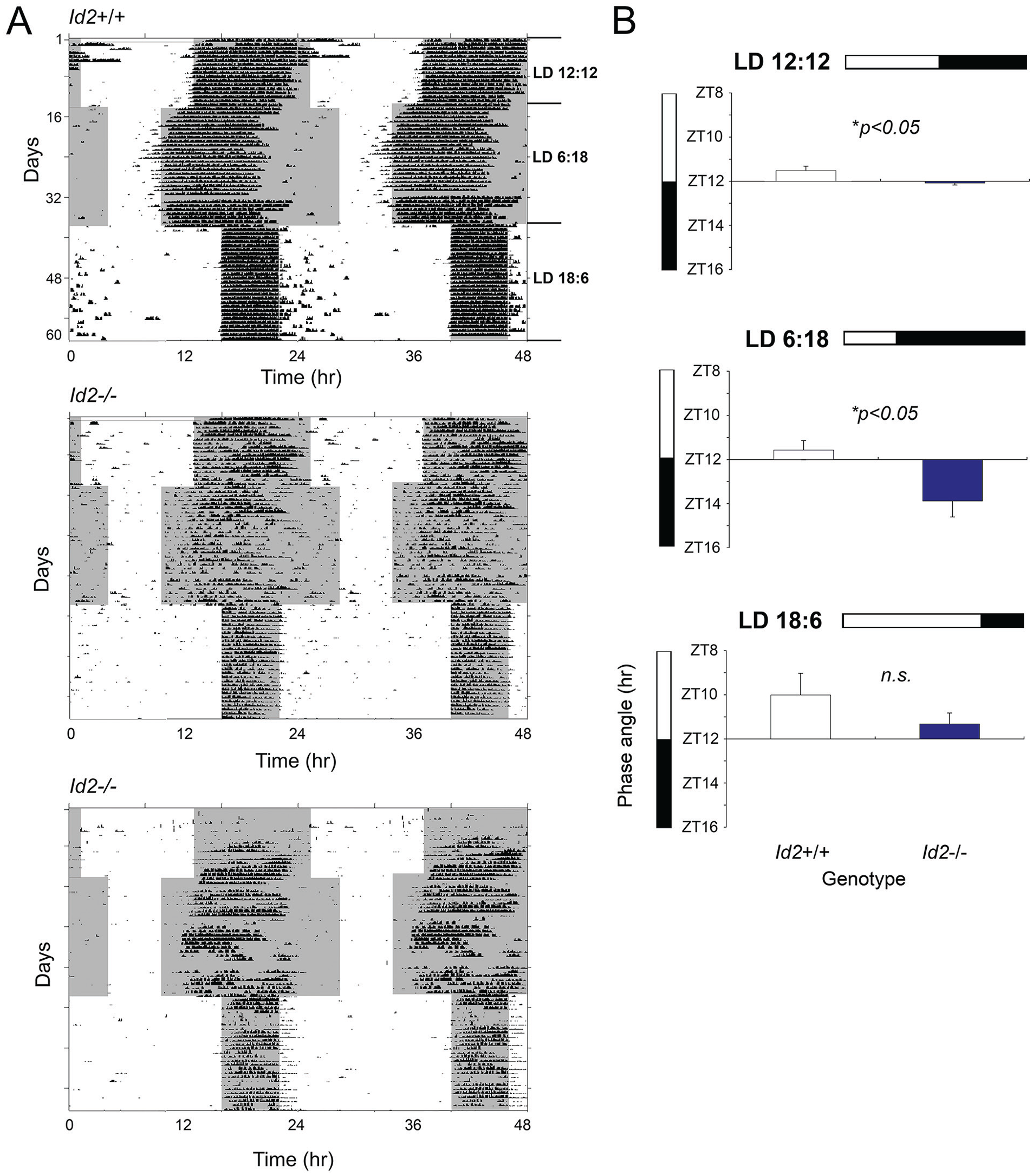
Id2 mutant mice exhibit a delayed phase angle of activity onset relative to lights-off under standard and short photoperiods. (A) Locomotor activity records (wheel-running) of representative wild-type (Id2+/+) and Id2-/- mice under 12:12 (standard), 6:18 (short), and 18:6 (long) LD cycle conditions are shown in double-plotted format. Each horizontal line represents a 48-h period, and the second 24-h period is plotted to the right and below the first. Vertical bars represent periods of wheel-running activity. (B) Phase angle of activity onset to lights-off (zeitgeber time 12, ZT12) of wild-type and Id2-/- mice on 12:12, 6:18, and 18:6 LD cycles. Values are group means ± SEM for wild-type (n = 13) and Id2-/- (n = 14) mice. Significant differences between genotypes were detected (analysis of variance, followed by post hoc t tests, *p < 0.05). Color version of the figure is available online.
Id2-/- Mice Exhibit Abnormally Large Phase Delays But Normal Phase Advances in Response to Continuous Light
One of the most striking findings of our earlier studies was that the number of days required to entrain to a new LD cycle was considerably reduced in Id2-/- mice when exposed to an abrupt 10-h delay of the LD cycle (Fig. 2A; Duffield et al., 2009). Furthermore, when mice were exposed to 10 h of continuous light starting at ZT12 but then transferred into constant darkness to ascertain the magnitude of the resultant phase shift of the clock, Id2-/- mice exhibited phase shifts that were almost twice the size of control mice (Duffield et al., 2009). In the current study, this specific LD to DD transition experiment was repeated on a new cohort of mice, and consistent with the earlier work, Id2-/- mice exhibited larger phase delays (150 lux: Id2+/+, 2.26 ± 0.46 h; Id2-/-, 4.63 ± 0.44 h; t28 = −3.7, p < 0.001); that is, corresponding phase shifts of the activity rhythms were twice as large in Id2-/- mice. To explore this finding further and determine whether such responses could be achieved under phase advancing parametric entrainment conditions, we exposed mice to a large 6-h advance of the photo schedule. Under these conditions, mice of all 3 genotypes showed similar responses to the protocol, taking an average of 6 days to achieve stable entrainment (1-factor ANOVA, F2,33 = 0.22, n.s.; Fig. 2B).
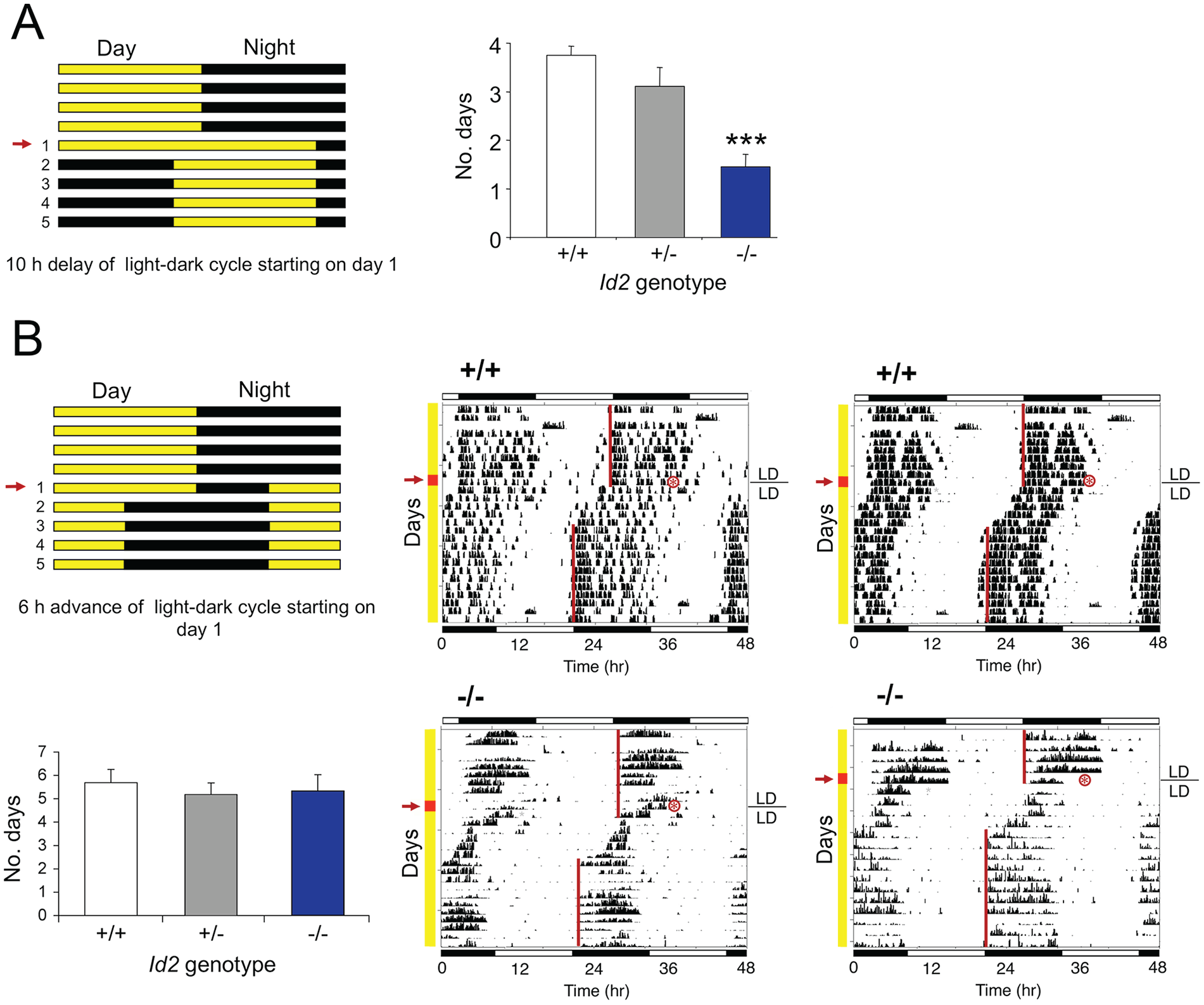
Id2 mutant mice show abnormally rapid entrainment to a delay but not to an advance of photo schedule. (A) Wild-type and Id2-/- mice exposed to a 10-h delay of the LD cycle by a 10-h extension of the light phase on day 1 of the LD cycle. Mice were maintained on an LD cycle for at least 14 days and transferred to a new LD cycle. Protocol (left) and mean ± SEM numbers of days required for stable entrainment to the new photoschedule for wild-type (n = 11), heterozygote (n = 9), and mutant (n = 11) mice (right). Numbers on the left of the protocol indicate the number of days after the transition to the new LD cycle, and the arrow indicates the day of treatment (day 1). Values marked by asterisks are statistically significant; analysis of variance followed by Dunnett’s post hoc tests (***p < 0.001). Data in the histogram are reproduced from Duffield et al. (2009). (B) Wild-type and Id2-/- mice exposed to a 6-h advance of the LD cycle by shortening the light phase by 6 h on day 1 of the LD cycle. Protocol (top left) and representative locomotor activity records (wheel-running) of 2 wild-type and 2 Id2-/- mice (right). Numbers on the left of the protocol indicate the number of days after the transition to the new LD cycle, and the arrow indicates the day of treatment (day 1). The timing of the LD cycles on the actogram is indicated by the white/black bars above and below the records, and the yellow line on the left of the vertical axis shows the days before and after the transition to the new LD cycle. The red line and arrow on the left indicates the day of treatment (day 1), and the actual mid-time of treatment is marked by a red asterisk. A line is fitted to the phase of activity onset for several days before and after the shift of the LD cycle. Mean ± SEM numbers of days required for stable entrainment to the new photoschedule for wild-type (n = 12), heterozygote (n = 11), and mutant (n = 13) mice (bottom left). No significant difference was detected between genotypes (analysis of variance, n.s.).
No Genotypic Differences in Phase Delays Produced by Discrete Saturating Light Pulses (Nonparametric Entrainment)
To further explore the response of the Id2-/- mice to the phase-delay portion of the circadian cycle, we tested mice under nonparametric entrainment conditions. Mice were exposed to a discrete pulse of light during early subjective night, predicted to generate a phase delay of the clock. Mice were maintained under constant darkness for 10 days before being treated with an 800-lux 30-min light pulse at CT14 (Suppl. Fig. S1A). To our surprise, no difference was detected between genotypes in the magnitude of the phase shifts produced by this discrete light pulse treatment (F2,32 = 2.24, n.s.).
Id2-/- Light PRC
To further explore the Id2-/- mice responses to photic stimuli under nonparametric conditions of entrainment, a standard entrainment protocol using discrete exposure to pulses of bright white light was used and allowed for the generation of a photic PRC (Schwartz and Zimmerman, 1990; von Gall et al., 1998). We generated a PRC for free-running Id2-/- and Id2+/+ mice treated with a 30-min saturating light pulse delivered at specific times across the circadian 24 h (CT0, CT4-8, CT12, CT14, CT16, CT18, CT20, and CT22), and the resultant phase shifts quantified (Suppl. Fig. S1B). The magnitude of the phase shifts and the shape of the PRC were similar between genotypes (2-factor ANOVA: effect of time, F6,154 = 53.33, p < 0.0001; effect of genotype, F1,154 = 0.29, n.s.; interaction, F1,154 = 1.60, n.s.).
Speed of Clock Resetting Is Similar between Genotypes
In WT mice and hamsters, the SCN clock is reset by discrete light treatments within 1 to 2 h after the light pulse, with 1.0 h being insufficient time for a shift to be resolved. This principle can be tested by applying discrete pulses of light at differing intervals after an initial pulse and ascertaining the accumulative shift of the clock (Best et al., 1999). These double-pulse experiments reveal that the resulting phase of the circadian system after an abrupt delay of the circadian system can represent a composite of parametric effects of light and the cumulative effects of light pulses that stimulate the phase-delaying portion of the PRC. Similarly, multiple-pulse protocols reveal similar properties for phase advances of the clock (Best et al., 1999). We tested the hypothesis that the more rapid reentrainment to abrupt delays of the LD cycle in Id2-/- mice could be due to a shortening of the time it takes for the clock to reset to the first pulse, which would lead to cumulatively larger phase delays and quicker reentrainment. To test this, we exposed Id2+/+, Id2+/-, and Id2-/- mice to fixed durations of light (15 min, 250 lux) interspaced with varying intervals of dark (45 min, 75 min, and 105 min; Fig. 3A-C) over a 10-h period, after which the animals were released into constant darkness. The cumulative phase shifts produced were then compared against the effect of the 10-h continuous light protocol as previously detailed (250 lux; Fig. 1; Fig. 3D). If delays were occurring substantially more rapidly, the serial pulses would continue to fall in the delay portion of the PRC. Alternatively, if resetting was slower, the later pulses would fall around and beyond the crossover point to which phase advances begin to occur. Therefore, a more rapid resetting of the clock would predict a larger accumulated phase delay for Id2-/- animals than WT animals in the group with pulses separated by 1 h but not in the group with pulses separated by 2 h, as both genotypes should have completed the delay in response to the first pulse after 2 h. However, no significant difference was observed between genotypes in the size of accumulative phase shifts produced by any treatment protocol, this being in the order of a ~3-h average shift for each group (Fig. 3E). Only in the continuous light treatment group and only in the Id2-/- mice was there an increase in the magnitude of the phase shift (average shift was ~6 h; ANOVA, F11,73 = 3.32, p < 0.05; Dunnett’s post hoc tests, p < 0.05). This difference between genotypes is consistent with our current findings described above and our earlier reports (Duffield et al., 2009).
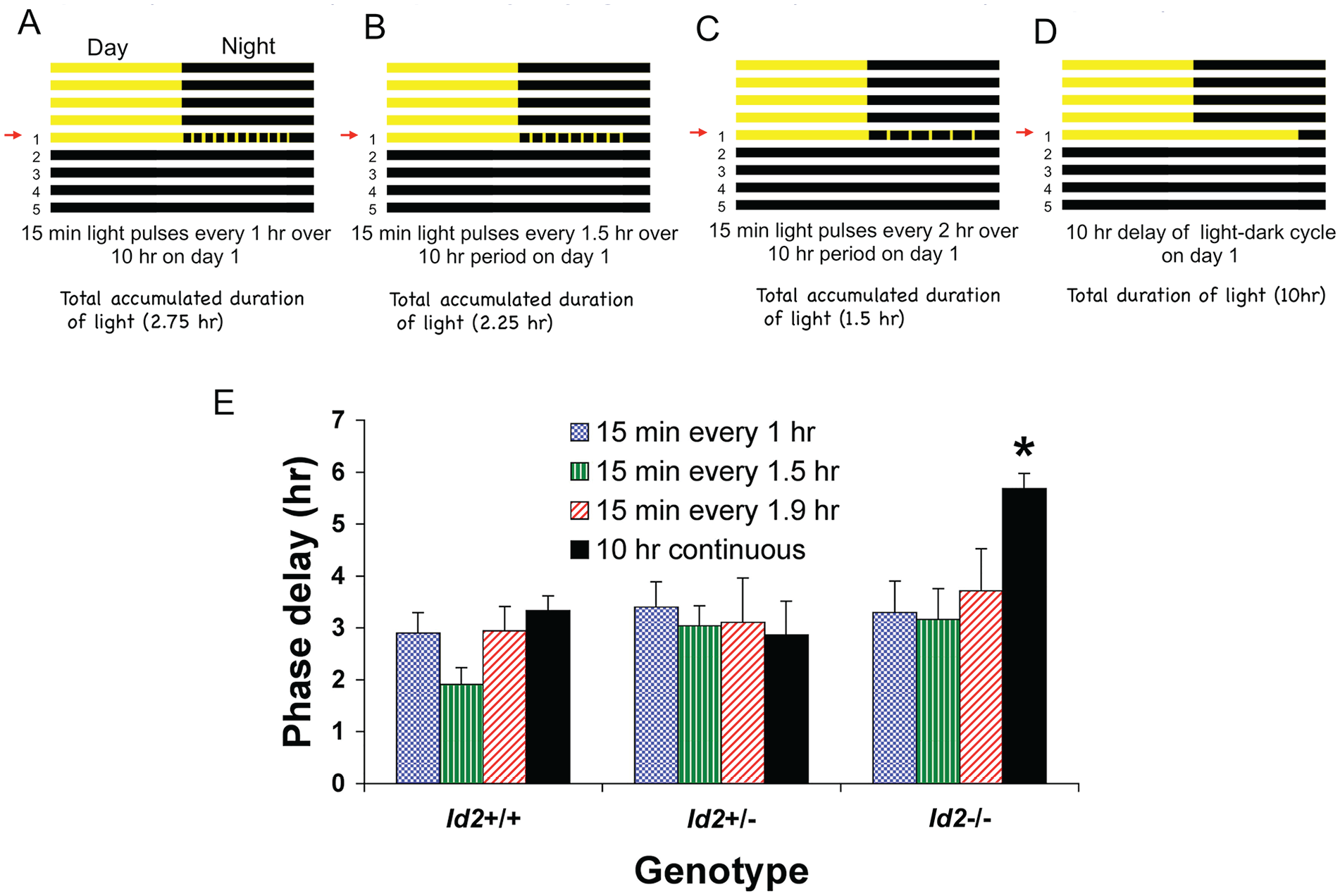
Speed of circadian clock resetting is similar between Id2-/- and wild-type mice. Mice under free-running conditions in DD were subjected to 1 of 4 treatment regimens: (A) a series of 15-min pulses of light delivered every 1 h over a 10-h period starting at ZT12, (B) a series of 15-min light pulses delivered every 1.5 h over a 10-h period starting at ZT12, (C) a series of 15-min light pulses delivered every 2 h over a 10-h period starting at ZT12, or (D) a 10-hour delay of the photocycle with an extension of the LD cycle with continuous light starting at ZT12. Numbers on the left of the protocol indicate the number of days following the transition to the new LD cycle, and the arrow on the left indicates the actual day of treatment (day 1). (E) Mean ± SEM magnitude of the phase delay for wild-type (n = 10), heterozygote (n = 5), and Id2-/- mutant mice (n = 12). No difference was observed between genotypes in the size of accumulative phase shifts in experiments testing the photo-refractory duration with a series of discrete light pulses (15 min, 250 lux) interspaced by varying time intervals (1, 1.5, and 1.9 h). Only the 10-h continuous light Id2-/- group showed a difference, consistent with previous experiments (see Fig. 2A). Analysis of variance followed by Dunnett’s post hoc tests, *p < 0.05 against all other groups. Color version of the figure is available online.
Id2-/- Mice Show Larger Phase Shifts in Response to Low-Illuminance/Short-Duration Light Pulses But Not to a Saturating Light Pulse
Id2-/- mice tested with a discrete saturating light pulse at CT16 (30 min, 800 lux), a phase of the mouse PRC that shows predictably large phase delays (Suppl. Fig. S1B), showed phase shifts with a size comparable to that of WT mice (Fig. 4A). Because of the absence of a genotypic difference, we then tested mice at the same circadian phase for responses to a light pulse of differing combinations of low illuminance and short duration, specifically 8 lux for 4 min versus 100 lux for 1 min versus 100 lux for 4 min. This experiment revealed differential responses in the Id2-/- mice (2-factor ANOVA: effect of treatment [light intensity × duration], F3,79 = 7.37, p < 0.001; effect of genotype, F1,79 = 8.14, p < 0.01; interaction, F3,79 = 2.9, p < 0.05). The 4-min 8-lux treatment resulted in a 1.9-fold increase above that of WT controls in the magnitude of the resultant Id2-/- phase shift (p < 0.01; Fig. 4B). A similar differential response was observed in Id2-/- mice treated for a shorter duration but increased illuminance of 1 min at 100 lux (WT, 29 ± 8 min; Id2-/-, 66 ± 15 min; mean ± SEM phase shift; p < 0.05). However, this difference in the magnitude of the phase shift was lost when both the pulse duration and intensity were increased to 4 min at 100 lux and when the magnitude of the shifts approached saturation (WT, 94 ± 10 min; Id2-/-, 88 ± 11 min; mean ± SEM phase shift; n.s.; for comparison, see the 30 min at 800 lux treatment; Fig. 4A).
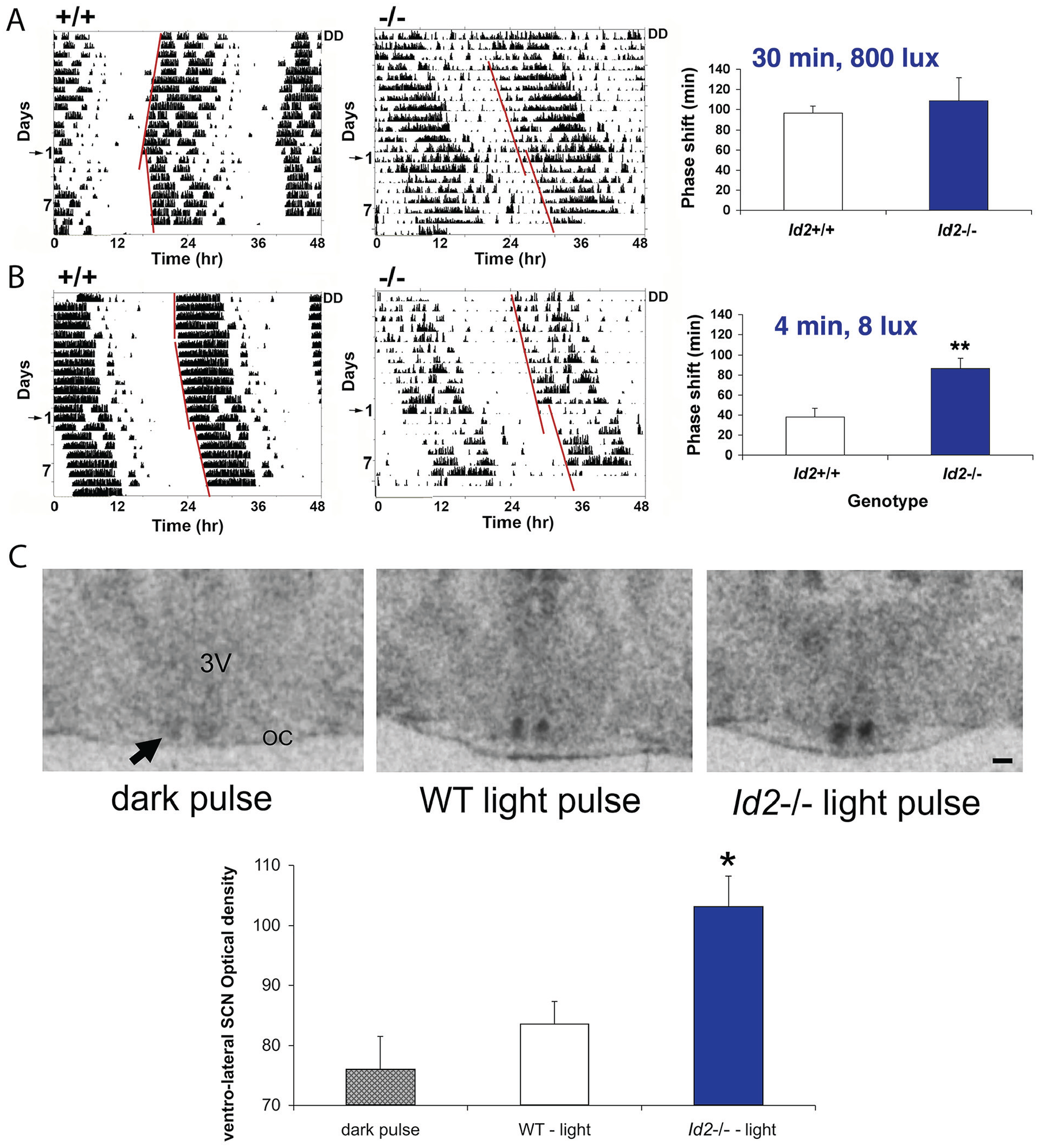
Id2-/- mice treated with a single subsaturating discrete light pulse produces a larger phase shift of the clock and an enhanced level of period 1 gene expression in the suprachiasmatic nucleus (SCN). (A) A single saturating discrete light pulse at CT16 (30 min, 800 lux) results in a comparable size phase shift between genotypes. Wild-type and Id2-/- mice (n = 20 and 16, respectively) were maintained in DD, and wheel-running activity and general activity (passive infrared motion detector) were recorded. Representative wild-type (WT) wheel-running activity (left) and Id2-/- general activity, as measured by a passive infrared motion detector (right), are presented. Numbers on the left indicate the number of days following the light treatment, with day 1 marked by the arrow on the left, being the day of treatment. A line is fitted to the phase of activity onset before and after the light treatment, and the time difference between the 2 lines is the measured phase delay of the free-running rhythm. No difference was observed between genotypes in the magnitude of phase shifts (analysis of variance [ANOVA] followed by post hoc t tests, n.s.), consistent with data shown in Supplemental Figure S1. (B). A single subsaturating discrete light pulse at CT16 (4 min, 8 lux) produced a 1.9-fold increase in the magnitude of the phase shift. Representative WT wheel-running activity (left) and Id2-/- wheel-running activity (right) are presented. Treatment of wild-type (n = 15) and Id2-/- mice (n = 10) with short duration/low light resulted in a significant difference in phase response between genotypes (ANOVA followed by post hoc t tests, **p < 0.01). (C). Light induction of per1 gene expression is increased in the Id2-/- mouse suprachiasmatic nucleus following a subsaturating discrete light pulse at CT16 (4 min, 8 lux). Wild-type and Id2-/- mice treated with 4 min of light (8 lux) at CT16 and brain harvested 1 h later; per1 gene expression assessed by in situ hybridization using a radiolabeled probe, n = 2 to 4, 12-µm coronal sections per animal (WT/Id2-/- dark pulse, n = 7; WT light pulse, n = 6; Id2-/- light pulse, n = 6). 3V, third ventricle; OC, optic chiasm. The arrow marks the SCN region. Scale bar = 100 µm. Quantification of expression in the ventrolateral SCN. A template of the light-induced zone was applied to all sections, and data were normalized to signal intensities in the corpus callosum. WT and Id2-/- dark pulse controls, with relative expression values of 78.3 ± 9.3 and 74.5 ± 7.5 (mean ± SEM), respectively, were combined for the purposes of statistical analysis. Significant elevation observed in per1 expression in Id2-/- relative to WT mice (ANOVA, followed by Dunnett’s post hoc tests, *p < 0.05). Color version of the figure is available online.
Light Induction of per1 Gene Expression Is Increased in Id2-/- SCN in Response to a Low-Illuminance/Short-Duration Light Pulse
Because per1 is a state variable of the clock, and it is induced by light as part of the mechanism by which the clock is shifted/entrained (Albrecht et al., 1997; Shearman et al., 1997; Shigeyoshi et al., 1997; Sun et al., 1997; Travnickova-Bendova et al., 2002; Dibner et al., 2010; Hughes et al., 2015), we investigated whether the gene induction response to light in the Id2-/- mouse was enhanced. We specifically tested animals with light at a subsaturating dose matching that which revealed a differential phase shift response in the Id2-/- mice (i.e., 4 min, 8 lux; Fig. 4B). Animals were treated at CT16, tissue harvested 1 h later, and per1 gene expression assessed by ISH. The expression of per1 in the ventrolateral SCN in light-treated Id2-/- was significantly greater than that detected for WTs (1-factor ANOVA, F3,15 = 3.3, p < 0.05; Dunnett’s post hoc tests, p < 0.05; Fig. 4C), a result consistent with an increased magnitude of the behavioral phase shift (Fig. 4B).
Retinal Anatomy and Pupil Constriction Responses Are Similar between Id2-/- and WT Mice
It is plausible that the Id2-/- photoentrainment phenotype could be explained by anatomical and/or functional changes to the retina and/or retinal hypothalamic tract (RHT), critical components of the photic input pathway to the SCN. To test this possibility, we examined the anatomy of the retina between genotypes by histological analysis. The qualitative images and the quantitative data revealed no significant differences between the retinal layers of Id2+/+ and Id2-/- mice (t tests, n.s.; Suppl. Fig. S2A-B). Furthermore, there was no difference in the number of RGCs between genotypes (t8 = 0.96, n.s.; Suppl. Fig. S2B), including when examined at the subregional level (left-lateral, right-lateral, and medial retina; data not shown). Because the melanopsin-positive intrinsically photoreceptive (ip) RGCs represent an important contribution to accessory visual functions, namely, circadian entrainment and pupillary response (Hattar et al., 2002; Hughes et al., 2015), we examined this population of cells. A representative image of fluorescent-immunostained melanopsin shows where ipRGCs are located in the retina and their low population density (Suppl. Fig. S2C), consistent with other reports (Hattar et al., 2002; Hughes et al., 2015). No significant difference in the quantity of the ipRGCs, as counted by the number of cell soma, was detected between Id2+/+ and Id2-/- mice (t test, n.s.), this being an average of ~2% of the RGC population. To test the retina at the function level, and based on the fact that components of the retina and retinal pathway, namely, projections of the ipRGCs and the RHT/retinothalamic tract, are shared by the circadian and pupillary systems (Hattar et al., 2002; Berson, 2003; Hattar et al., 2003; Hughes et al., 2015), we tested pupil constriction responses in dark-adapted Id2-/- mice at 2 different intensities of white light, 10 and 100 lux. Based on the qualitative data of the images and the quantitative data of the comparison of relative pupil apertures over a period of 1 min (Suppl. Fig. S2D), there was no difference in the rate or magnitude of pupil constriction in response to light between Id2+/+ and Id2-/- mice at either light intensity (Bonferroni multiple measures corrected t tests, n.s.).
Amplitude of per1 and PER2 Rhythms is Normal in Id2-/- SCN
One explanation for the photoentrainment phenotype is that the amplitude of state variables of the clock is lower. A limit cycle can be used to model this aspect of the circadian clock and explain its responses to clock resetting and the magnitude of phase shifts (Jewett et al., 1991; Winfree, 2001; Johnson et al., 2003; Dunlap et al., 2004; Vitaterna et al., 2006). A prediction is that a lower amplitude pacemaker would allow the same intensity of a zeitgeber stimulus to result in a larger shift of the clock. To test this hypothesis, we examined the expression of the clock genes per1 and per2, both state variables of the clock, in the SCN by ISH and by luminometry in SCN tissue explants. Tissues were harvested every 4 h over the circadian day, and ISH analysis revealed an expected rhythm in per1 gene expression that peaked at CT4-CT8. These expression profiles are comparable with other studies (Shearman et al., 1997; Sun et al., 1997). However, no difference was observed in the expression levels between genotypes at any of the circadian times (2-factor ANOVA: effect of time, F5,28 = 23.8, p < 0.0001; effect of genotype, F1,28 = 0.49, n.s.; interaction, F5,28 = 0.60, n.s.; Suppl. Fig. S3). Similar results were obtained for the SCN studied ex vivo in cultured tissue explants harvested from both PER2::LUC and per1-luc mice (Fig. 5). In both cases, robust rhythms in protein and gene expression, respectively, were detected for both Id2-/- and WT tissue (Fig. 5A). Moreover, consistent with the ISH data, we observed no difference in the peak-to-trough amplitude for either PER2 protein (t14 = 0.81, n.s.) or per1 gene expression (t13 = 0.58, n.s.; Fig. 5D). In addition, we observed no difference in the period length of the clock in the cultured Id2-/- SCN (PER2::LUC, F2,18 = 1.22, n.s.; per1-luc, F2,24 = 0.27, n.s.; Fig. 5B), a finding consistent with our previous data from the analysis of behavioral locomotor activity in the intact animal (Duffield et al., 2009). Finally, there was no difference in peak phase between the Id2 genotypes of both PER2::LUC (F2,22 = 0.42, n.s.) and per1-luc mice (F2,28 = 0.88, n.s.; Fig. 5C), both occurring at predicted phases of the circadian/diel cycle (Wilsbacher et al., 2002; Yoo et al., 2004): PER2::LUC peaked between ZT11.5 and ZT11.8, whereas per1-luc peaked between ZT6.5 and ZT6.63.
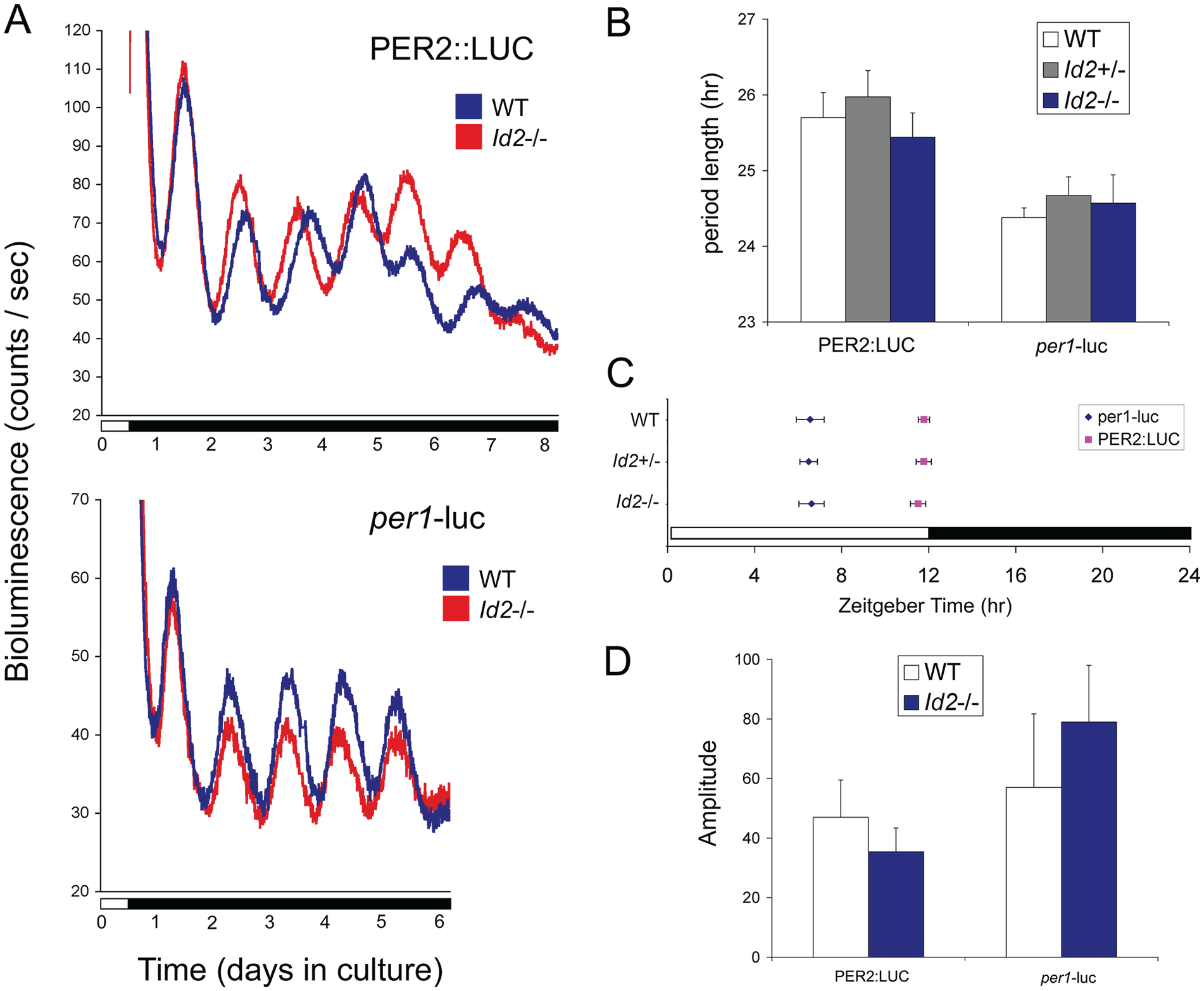
Period length, phase, and amplitude of suprachiasmatic nucleus (SCN) PER2::LUCIFERASE and per1-luciferase rhythmic expression are comparable between Id2-/- and wild-type mice. (A) Representative raw bioluminescent time-course data from 300-µm SCN tissue explants from PER2::LUC (top) and per1-luc (bottom) adult mice crossed with Id2-null mice. Wild-type and Id2-/- SCN. (B) Mean ± SEM period length determinations. No significant differences in period lengths were found between genotypes Id2+/+, Id2+/-, or Id2-/- (analysis of variance, n.s.). SCN tissue was derived from PER2::LUC or per1-luc wild type (n = 5, 6), Id+/- (n = 8, 15), and Id2-/- (n = 8, 6) mice and prepared between ZT8 and ZT11.5 from mice on a 12:12 LD cycle. (C) Mean ± SEM phase determinations. There was no difference in phase between genotypes for PER2::LUC (analysis of variance [ANOVA,[ n.s.; squares) or per1-luc (ANOVA, n.s.; diamonds) mice. Peak phase was highly conserved between samples: PER2::LUC peaked in the first full cycle at ~ZT12.0 and per1-luc at ~ZT6.5. PER2::LUC or per1-luc wild type (n = 7, 6), Id+/- (n = 9, 18), and Id2-/- (n = 9, 7) SCN. (D) Mean ± SEM amplitude determinations. The mean amplitude of the first 1 days of rhythms displayed no significant difference between wild type and Id2-/- for both PER2::LUC and per1-luc mice (t test, n.s). Amplitude was defined as the mean acrophase-nadir counts per second (cps) from the first 3 cycles. PER2::LUC or per1-luc wild type (n = 7, 7) and Id2-/- (n = 10, 8) SCN. Color version of the figure is available online.
Crucial in the analysis of the Id2-/- circadian system is the preservation of a key feature of the photoentrainment phenotype in the line of mice crossed between the original Id2-/- and PER2::LUC colonies. Importantly, we specifically tested these mice using the parametric entrainment protocol shown in Figure 3D and revealed that Id2-/- mice on the PER2:LUC genetic background continued to exhibit the photoentrainment phenotype (phase shift response to 10 h continuous light, 150 lux: Id2+/+/PER2::LUC, 2.24 ± 0.34 h; Id2-/-/PER2::LUC, 3.92 ± 0.28 h; t14 = −3.8, p < 0.01). Therefore, the Id2 entrainment phenotype persists in this genetic background, but still no amplitude effect was observed in the rhythm of PER2 protein expression within the SCN.
Discussion
ID2 is an HLH transcriptional repressor, and earlier investigations have revealed a contribution of ID2 in circadian regulation at the 3 fundamental levels of the clock, namely, its input (including entrainment), core clock function, and output (Duffield et al., 2009; Ward et al., 2010; Adachi et al., 2013; Hou et al., 2009; Mathew et al., 2013). In this study, we have explored the photoentrainment phenotype of the Id2-null mouse, using protocols based on both parametric and nonparametric models of entrainment. We show distinct alterations in the entrainment properties of the clock in Id2-/- that are unidirectional (delays and not advances), that are evident only when the circadian system is probed with subsaturating light pulses, and that likely explain the delayed phase angle of entrainment under different photoperiods and the increased rate of phase-delay reentrainment of the clock to abrupt phase shifts of the LD cycle (Fig. 6A). Further, the increased magnitude of light-induced phase delays in response to subsaturating light is accompanied by an increase in SCN regional per1 gene expression (Fig. 6B). Anatomical and functional (pupillometry) aspects of the retina were examined and found to be normal in the Id2-/- mouse, suggesting that the photoentrainment phenotype is not due to a developmental abnormality in the retina or RHT. Finally, using Id2-/-/per1 and Id2-/-/PER2 luciferase mice to assess ex vivo clock gene expression or ISH to assess expression in vivo, we confirmed that the circadian phase, period length, and the amplitude of the clock’s state variables in the SCN were normal, highlighting a stable clock in the Id2-null mouse. Together, our findings suggest that the augmented delays observed in Id2-null mice likely reflect increased nonparametric responses that are revealed only under low-intensity and short-duration pulse conditions but not to changes in the parametric responses of the clock to light (Duffield et al., 2009; Adachi et al., 2013; Mathew et al., 2013).
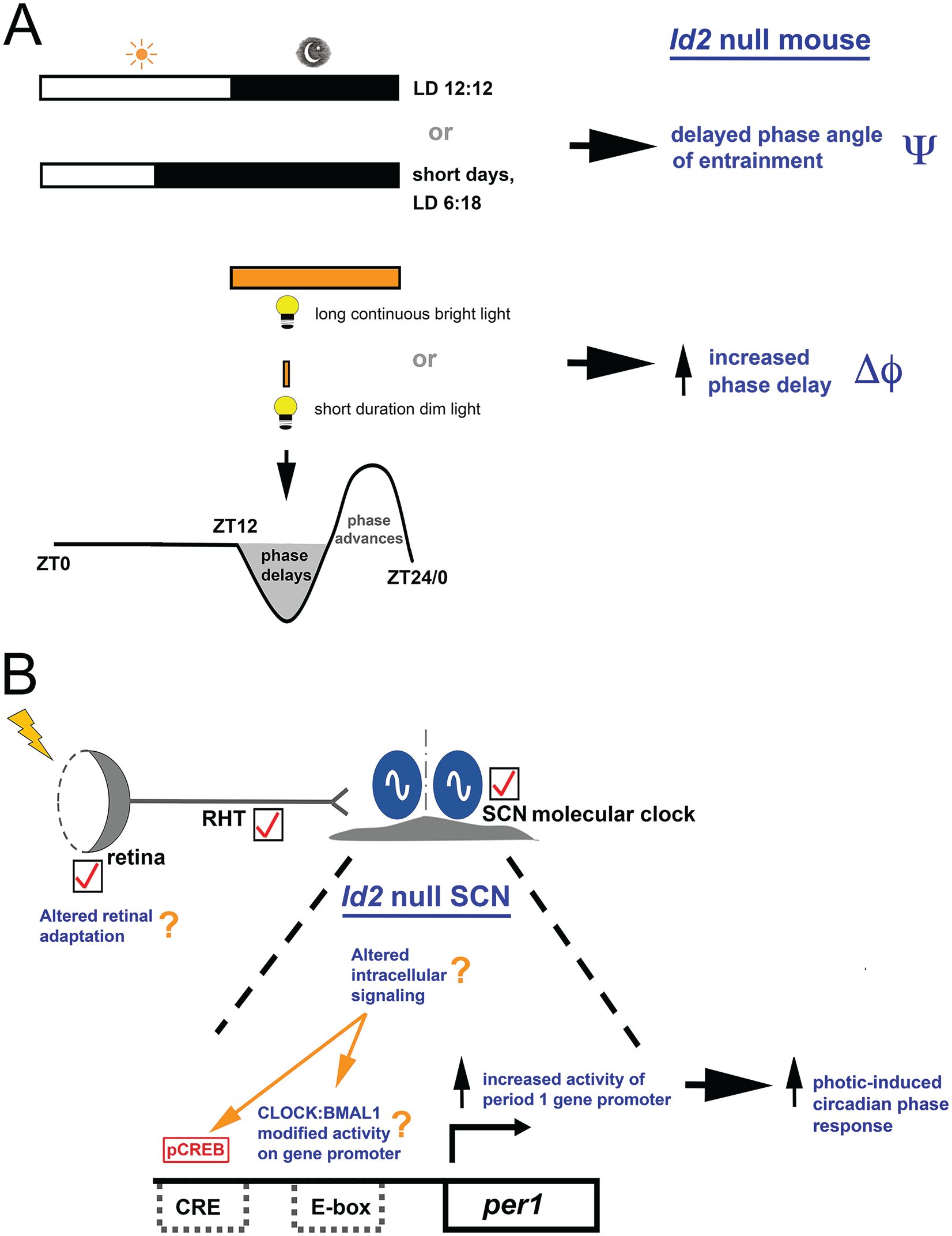
Absence of ID2 results in an altered photic responsiveness of the circadian system. (A) Id2-/- mice, under conditions of 12:12 LD or a short-day photoperiod exhibit a delayed phase angle of entrainment (Ψ) relative to wild-type (WT) mice. Long bright-light exposure during the delay portion of the phase-response curve (PRC) (parametric entrainment) or a short duration/dim light (nonparametric entrainment) produces a larger phase delay of the clock (Δφ) and faster entrainment. These properties of the clock cannot be explained by standard models of parametric and nonparametric entrainment, tested in various ways. The timing of the LD cycles is indicated by the white/black bars. The delay portion of the PRC is highlighted in gray to signify the phase of the cycle with a change in photic response. (B). The SCN is the likely locus of the Id2-/- photic responsiveness phenotype. The retina and retinal hypothalamic tract (RHT) structure/function are normal in Id2-/- mice, as is the phase, period length, and amplitude of SCN clock gene expression. The tick mark signifies a “normal” anatomy or function in the photoentrainment–circadian clock pathway, although a retinal contribution to the phenotype cannot be entirely excluded. In the Id2-/- mouse, short duration/dim light activates the SCN cells and increases activity of the period 1 clock gene promoter, as demonstrated by the elevation of light-induced per1 expression compared with WT mice. Likely targets for ID2 modulation of photoentrainment are CLOCK and BMAL1 via direct protein interaction with and inhibition by ID2 (Ward et al., 2010) and/or altered intracellular signaling that culminates in E-box and/or Ca2+/cAMP response element (CRE) activation at the per1 gene promoter. As per1 is a light-inducible state variable of the clock, a change in per1 expression produces a phase shift of the clock (Duffield et al., 2009; Ward et al., 2010). Thus, an enhanced responsiveness of this input pathway in Id2-/- mice results in increased photic-induced phase changes of the circadian pacemaker. CLOCK:BMAL1, heterodimer of bHLH-PAS transcription factors; pCREB, phosphorylated Ca2+/cAMP response element-binding protein. Color version of the figure is available online.
Phase Angle of Entrainment
Our earlier studies revealed a phenotype characterized by a delayed phase angle of entrainment. However, this was examined under a single photoperiod, specifically LD 12:12, and the differences reported were small (~30 min). In the current study, we examined the relationship of activity onset of locomotor activity under 3 different photoperiods. The phase angle was delayed relative to WT controls in LD 12:12 and short photoperiods. Furthermore, the phase angle relative to lights-off in Id2-null mice was ~0 h under LD 12:12, but under LD 6:18, it deviated by as much as 2 h. The difference in phase angle between WT and Id2-/- mice could be explained by a different parametric response to light in the different genotypes; that is, different photoperiods could induce different changes in period length (τ; Pittendrigh and Daan, 1976; Jones et al., 1999; Dunlap et al., 2004; Gronfier et al., 2007). However, this is unlikely, because the locomotor rhythms of Id2-null mice have a similar circadian period length as WT controls in DD and LL (Duffield et al., 2009; Adachi et al., 2013; Mathew et al., 2013), and ex vivo Id2-/- SCN clock gene expression rhythms have similar period lengths to controls (herein). This makes the phase angle phenotype curious, as it cannot be easily explained by current models of the clock. In other animal and human studies, an increased deviation of endogenous period length away from the environmental T cycle (e.g., 24-h LD cycle) or a decreased strength of the zeitgeber (e.g., light intensity) often result in marked increases in the phase angle (Pittendrigh and Daan, 1976; Jones et al., 1999; Dunlap et al., 2004; Gronfier et al., 2007). However, more recent genetic studies of the Neurospora clock question the assumed predictive/correlative relationship between period length and phase angle (Lee et al., 2017), and so the observation that Id2-/- mice express a normal period length but an altered phase angle is not a complete aberration. The phase angle difference in Id2-/- mice (smaller and delayed relative to WT mice under LD 12:12 conditions) is observed not only with wheel-running activity and by monitoring general activity within the cage but also in their profiles of feeding activity (Mathew et al., 2013), suggesting that this behavioral phase reflects the phase of the central pacemaker. A possible explanation that would be consistent with the findings of the current study is that the fixed light intensity used in the experiments effectively has a greater zeitgeber strength in the Id2-null circadian system, thereby decreasing the phase angle and delaying it relative to WT controls. This phenotype is also exaggerated when Id2-/- mice are maintained on a short LD 6:18 photoperiod, but the phase angle difference, although still delayed relative to WTs, deviates further from the phase of activity onset than that of the WTs (Id2-/-, delayed 2 h versus WT, advanced 20 min). It is therefore reasonable to consider that the different phase of entrainment can be explained by more powerful delaying effects in Id2-/- mice, which would move the early subjective night away from the evening light.
It is important to note that all phase estimates in the study were based on the activity onset of locomotor activity and were determined from both wheel-running and general activity (using a passive infrared motion detector), an approach used previously (Duffield et al., 2009; Mathew et al., 2013). Where both records were available for each animal, phase values were based on an average of the two, providing additional confidence in the phase determination. Importantly, the phase calculations were concordant, revealing little variability between the 2 assessments of circadian phase.
Phase Delays But Not Phase Advances Are Modified
In the current study, we explored the directionality of the photoentrainment phenotype. Our previous report revealed a significant increase in the speed of entrainment in response to a 10-h delay of the LD cycle (Duffield, 2009), resulting in an increase from 3.5 days in WTs to 1.5 days for Id2-/- mice. The clock can be reset in 1 of 2 directions, delays and advances, with light usually generating delays during the early night and advances during the late night. A large 6-h advance of the LD cycle falling in the predicted phase advance portion was applied to experimental mice. In all genotypes, the average number of days to reach stable entrainment was 5.5. Therefore, these data reveal a unidirectional phenotypic response in the Id2-/- mice to photic entrainment under continuous/parametric conditions.
Phase-Response Curve
To understand how the Id2-/- phenotype could be explained by nonparametric models of entrainment, we exposed mice to discrete treatments of light under free-running DD conditions. The initial investigation focused on testing the phase of the circadian cycle responsible for producing large phase delays of the clock, specifically at CT16. Here, we found no differences between genotypes, in all cases eliciting large ~2-h average phase delays. To try to explain the discrepancy between the results of the parametric and nonparametric entrainment experiments, we generated a photic PRC by testing mice at 2-h intervals across the circadian night. Surprisingly, we found little genotypic difference, and all genotypes exhibited a predictable photic PRC (Schwartz and Zimmerman, 1990; von Gall et al., 1998).
Speed of Clock Resetting
The circadian clock of wild-type rodents can be reset by light within 2 h but not 1 h (Best et al., 1999). This principle can be tested by applying discrete pulses of light at differing intervals after an initial pulse and ascertaining the accumulative shift of the clock. We hypothesized that the speed of resetting of the clock in Id2-/- mice was faster, thereby allowing for a larger size of accumulative phase shifts to occur. We challenged mice with a series of discrete light pulses (15 min, 250 lux) interspaced with varying time intervals. No differences were detected between genotypes in the magnitude of composite phase shifts produced in response to any of the 3 different interval protocols. If Id2-/- mice have a profoundly shorter clock-resetting time, an increased composite phase delay is predicted for the 1-h and/or 1.5-h interval protocol groups. These results suggest that the speed of resetting of the circadian system in Id2-/- mice is normal and indistinguishable from WT or heterozygous mice. Only in the 10-h continuous light treatment group and only in the Id2-/- mice was there an increase in the magnitude of the phase shift; this difference between genotypes was consistent with our earlier findings and our previous reports (Duffield et al., 2009). These data reveal no obvious difference in the speed of clock resetting in Id2-/- mice but do highlight the unique feature of continuous light exposure in producing enhanced phase delays in only the mutant animals. This is a profoundly different response compared with WT and heterozygote controls, in which the magnitude of the phase shift is similar despite the 4 protocols exposing them to differing durations of accumulated light during the night (i.e., 1.5 h, 2.25 h, 2.75 h, or 10 h). A possible limitation of the experiment is that the testing of differences in resetting speed was based on protocols that changed the interspacing of the start of the pulses by 30 min, as in 1, 1.5, and 2 h. It is plausible that this interval is too large to allow for discrimination between resetting speeds should these genotypic differences be small.
Photoresponsiveness
A change in relative delay and advance portions or their magnitudes within the PRC or an increased speed of resetting of the clock would provide a mechanism to explain the large phase delays and rapid entrainment produced by continuous light. However, we did not observe such differences in the Id2-/- mice. We therefore explored whether light treatments of shorter duration and lower intensity might differentially affect Id2-/- mice. Unlike a discrete light pulse response at CT16 using a pulse of long duration (30 min) and high illuminance (800 lux), Id2-/- mice treated with a single subsaturating light pulse at the same circadian phase exhibited an almost 2-fold increase in the magnitude of the phase shift above that of WTs. Responses were similar whether the conditions were 4 min at 10 lux or 1 min at 100 lux, but the effect was absent at 4 min at 100 lux. These observations correlate with our earlier finding of a 1.8-fold larger mean phase delay with a treatment of 10 h of continuous light.
Phase adjustments of the mammalian clock by light have been shown to exhibit different temporal dynamics depending on the length of exposure. The relationship between the duration of the light pulse and the magnitude of the resultant phase shift follows a linear/log trajectory during the first hour after light onset. However, after 1 h, it switches to a linear/linear relationship (Kronauer et al., 2019; Nelson and Takahashi, 1991; Comas et al., 2006). In other words, the circadian system is highly responsive early on to exposure to the light and especially during the first 15 min, but after 1 h, it becomes less responsive. However, the system does remain responsive for the duration of a long exposure (e.g., 10 h). Thus, the normal circadian system undergoes a change(s) in its state of photosensitivity or responsiveness over the course of a photic stimulus. With this background, it is surprising that the exaggerated phase responses of the Id2-/- mice are revealed when challenged by a short (1 min or 4 min [10-100 lux]) or long treatment (10 h [150-300 lux]) but not with a pulse of intermediate duration (30 min [800 lux]). Irradiance may also be an important independent variable in contributing to the phenotype (Nelson and Takahashi, 1991; Kronauer et al., 2019), and separating the duration and intensity factors will be important in future investigations. Thus, the enhanced phase responses are revealed at the extremes of photic stimuli, which in WT animals are known to be in different physiological states of responsiveness. It is plausible that the Id2-/- circadian system can be driven to a state of heightened responsiveness, suggesting a role for ID2 in reducing phase adjustment responses of the circadian system when challenged with extremes of light exposure (dim short exposure versus bright long exposure) and to ensure correct integration of photic information but contributing little to regulating the system during moderate exposure. This may explain why genotypic differences were revealed only during these specific extreme conditions and that are otherwise masked when challenged with a standard 30 min/800 lux exposure.
To better understand the underlying physiology of this response, we challenged mice with 4 min at 10 lux at CT16 and examined the induced levels of expression of a state variable of the clock (i.e., a clock gene). Consistent with the increased magnitude of the phase shift produced under these conditions of low illuminance and short duration, a significant elevation of light-induced per1 expression was observed in the Id2-/- ventrolateral (retinorecipient) SCN. These results are also consistent with the observation that Id2-/- fibroblasts in vitro, when serum stimulated, show ~2-fold higher levels of induced per1 expression (Ward et al., 2010).
Retinal Pathways
There are 2 ways that the SCN could respond differentially to a fixed treatment of light, determined by circadian phase of exposure, and by dose (light intensity × time): the clock within the SCN could be more sensitive to a fixed signal from the retinal afferents, or the retina or the RHT produce an elevated signal. The observations in Id2-/- fibroblasts (Ward et al., 2010) would suggest the former is more likely and that the SCN clock at the cellular level responds more to the zeitgeber signal. To test the latter possibility and the fact that Id genes are implicated in developmental processes (Wang and Baker, 2015), we examined retinal anatomy and undertook functional assessment of the accessory visual system by pupillometry. We found no differences between genotypes in retinal anatomy, including the population size of RGCs or numbers of melanopisin-positive RGCs. Pupillometry responses of dark adapted Id2-/- mice to light exposure showed normal responses both in the speed of the response and in the extent of pupil constriction. Therefore, these observations suggest that Id2-/- mice have normal retinal function and that the photoentrainment phenotype is not due to major aberrations in the structure/function of the retina or RHT.
Despite the normal gross anatomy and pupillometry responses, it is possible that a difference in the Id2-/- circadian system still might occur at the retinal level (Comas et al., 2006; Wong et al., 2005). As Id2-/- pupillary responses are normal when tested at 10 or 100 lux, it is unlikely that an abnormal retinal function underpins the enhanced short-duration/low-intensity photic phase shift responses and their SCN molecular correlates. However, it is possible that an altered Id2-/- retina process, such as adaptation, contributes to generating the abnormal circadian responses observed with photic exposure of greater duration and illuminance. If a factor, it likely involves a complex contribution from different classes of photoreceptor cells, namely, rods (especially for dim light responses), cones, and ipRGCs (especially for higher irradiance responses), and their signal integration (Butler and Silver, 2010; Lall et al., 2010). An important area of future work will be to further address any retinal contribution to the phenotype or to determine that the anatomical locus of the phenotype resides entirely within the SCN.
Limit Cycle Property of the Clock
Assuming that the locus of the phenotype is within the circadian clock cells of the SCN, one possible explanation for large phase shifts in Id2-/- mice is a difference in the amplitude of the rhythms of state variables of the clock: in theory, the smaller the molecular oscillations relative to the magnitude of the zeitgeber, the larger the resultant phase shift, in accordance with the limit cycle model of the clock (Jewett et al., 1991; Winfree, 2001; Johnson et al., 2003; Dunlap et al., 2004; Vitaterna et al., 2006). Our earlier results raised this as a possibility because a proportion of Id2-/- mice maintained in DD expressed lower-amplitude wheel-running locomotor activity rhythms (Duffield et al., 2009; Mathew et al., 2013). This mechanism has also been used to explain the phenotype of the Clock mutant mouse (Vitaterna et al., 2006). To test whether the amplitude of the molecular clock is smaller in Id2-/- mice, we examined the rhythmic profiles of the negative loop elements of the clock, per1 and per2, in constant dark conditions and ex vivo. Together, the ISH and tissue explant analyses of clock gene expression in the Id2-/- SCN reveal robustly rhythmic profiles, with no difference in the amplitude of the molecular clock. Therefore, the limit cycle property of the Id2-/- clock does not explain the photoentrainment phenotype. In addition, we found no difference in circadian phase or period length of rhythmic per1 or PER2 expression in SCN tissue explants. These results for the SCN are similar to the observations of clock gene rhythms in the Id2-/- liver, in which per1, per2, and bmal1 exhibit normal rhythmic expression profiles with no obvious change in amplitude or phase (Hou et al., 2009). Further, the Id2-/- mouse exhibits a normal free-running period length in locomotor activity in DD and in LL (Duffield et al., 2009; Adachi et al., 2013), indicating that ID2 is not required for clock function or molecular rhythmicity.
Molecular Redundancy
Noting that other members of the Id gene family, namely, Id1, Id3, and Id4, (1) are expressed rhythmically in the SCN, (2) share similar peak phases, (3) have the potential to interfere with the formation of a functional CLOCK:BMAL1 complex, and (4) reduce its capacity to transactivate downstream genes (Duffield et al., 2009; Ward et al., 2010), we propose a model in which multiple ID proteins act in concert. Given this potential redundancy, it is perhaps unsurprising that the Id2 null mouse circadian phenotype is somewhat subtle. Redundancy is not a new feature of the mammalian clock and is present for many of the canonical clock components, for example, cry1, cry2; per1, per2, per3; Clock, Npas2; Reverbα, Reverbβ; and Dec1, Dec2 (Dunlap et al., 2004; Liu et al., 2008; Erzberger et al., 2013; Papazyan et al., 2016).
Cellular-Molecular Mechanism
Some mechanistic insight into the Id2-/- phenotype can be found from the clock-resetting pathway that involves the CLOCK-BMAL1 complex (Shearman and Weaver, 1999; Jung et al., 2003; Shim et al., 2007; Lee et al., 2010; Lee et al., 2015). A current model poses that the protein kinase C–activated CLOCK becomes partnered with sumoylated BMAL1 together with CREB binding protein, which, as a complex, results in the activation of the per1 gene promoter via its E-box elements (Lee et al., 2015). Importantly, this event is achieved independently of the classical pCREB activation of the CRE of the per1 promoter (Travnickova-Bendova et al., 2002; Lee et al., 2010; Jagannath et al., 2013). This suggests that the most likely explanation for the Id2-/- entrainment phenotype is through the disturbed balance in available CLOCK and BMAL1 heterodimer and its entry into the nucleus (Ward et al., 2010), which in turn might disrupt the normal operation of this potent clock-resetting mechanism. Under normal physiological conditions, the time-specific expression of ID2 protein, as predicted from its circadian rhythm in gene expression and protein abundance (Duffield et al., 2002; Duffield et al., 2009; Hou et al., 2009; Adachi et al., 2013), would therefore be predicted to gate the nuclear entry and synthesis of functional CLOCK:BMAL1 heterodimer. This in turn would culminate in per1 gene promoter activation. The absence of ID2 would therefore result in an artificially greater activation of this pathway, presumably resulting in an enhancement of the inducible per1 expression and resultant increased magnitude of the circadian phase shift (Fig. 6B). In line with this model, zeitgeber-induced per1 expression is indeed elevated in the Id2-/- SCN after photic stimulation (herein) and in serum-stimulated Id2-/- fibroblasts, and in vitro assays demonstrate the ID2 interference of CLOCK:BMAL1 transactivation of the per1 promoter (Duffield et al., 2009; Ward et al., 2010). Furthermore, Adachi et al. (2013) observed elevated levels of per1 expression in Id2-/- SCN following transfer of mice from LD to LL.
Another possibility is that ID2 deletion results in an altered state of intracellular signaling in SCN cells, thereby becoming more responsive to the photic signal. Such a change might result in enhanced Ca2+ signaling and/or phosphorylation of CREB, thereby culminating in increased activation of the CRE of the per1 gene promoter and resulting in increased per1 expression. Future investigations will focus on elucidating such possible molecular-cellular mechanisms in SCN cells.
What the current in vivo studies suggest, however, is that the unique responsiveness of the Id2-/- circadian system is conditional on the intensity of the signal and the circadian phase of its presentation (early night with phase delays but not late night with phase advances). More specifically, these data reveal that with a saturating signal, the effect of the knockout phenotype is abolished, and only a “weaker” subthreshold discrete stimulus or a prolonged stimulus result in an increased phase shift response.
The hypothesis that this phenotype reflects an absence of what would be a normal action of ID2 upon CLOCK:BMAL1 heterodimer activity and/or modulation of intracellular signaling in the SCN central pacemaker (Fig. 6) needs to be tested empirically. However, the synthesis of prior in vivo and in vitro studies (Duffield et al., 2009; Hou et al., 2009; Ward et al., 2010; Adachi et al., 2013) and the Id2-null mouse experiments conducted herein suggest this to be a likely mechanism. Importantly, the current study reveals that the enhanced entrainment phenotype cannot easily be explained by the absence of ID2 resulting in a gross abnormality in the retina or RHT, changes in the PRC or in the speed of clock resetting (Best et al., 1999), or as having an effect on the amplitude of state variables of the clock that would result in an altered limit cycle (Jewett et al., 1991; Vitaterna et al., 2006). Furthermore, as the period length under constant DD or LL conditions is normal (Duffield et al., 2009; Adachi et al., 2013), the phenotype cannot be explained through an altered velocity of the clock, as might be predicted from the parametric model for entrainment (Johnson et al., 2003; Dunlap et al., 2004).
A Role for ID2 in Photoadaptation?
A component of the molecular clock comparable with ID2 can be seen in Neurospora, in which the VIVID protein contributes a TTFL that closes outside the core oscillator but that affects aspects of circadian timing (Heintzen et al., 2001; Hunt et al., 2010; Schafmeier and Diernfellner, 2011; Gin et al., 2013). VVD is a negative regulator of the White Collar Complex of the clock (homologous to CLOCK:BMAL1) and regulates the photic phase setting and responses to changing intensities of light (i.e., photoadaptation). Similar to ID2, rhythmic vvd expression is clock controlled, and VVD feeds back to regulate input and output gene pathways not required for circadian rhythmicity. Thus, the contribution of ID2 to the molecular circadian clock in the mammal may be an evolutionary convergent mechanism (Pando and Sassone-Corsi, 2001; Schafmeier and Diernfellner, 2011), controlling the photic response (functionally limiting the magnitude of phase shifts in low-light or continuous-light conditions) while also acting as a mediator of clock output.
Therefore, it is plausible that ID2 contributes to a photoadaptation mechanism to tune the circadian system to differences in light intensity, such that it responds more appropriately to the state of the environmental photic conditions. Clearly, further experimentation is required to explore this putative function for ID2 in the mammal, including specific contributions by the SCN and retina. However, the precedent for such a mechanism in rodents is apparent in the responses to changes in light intensity observed during differing durations of twilight transitions and photoperiod, conditions that depend on season and latitude (Boulos and Macchi, 2005; Comas and Hut, 2009). This process might also be relevant in correcting for spurious exposure to light during the night (e.g., moonlight or artificial light), environmental perturbations that might otherwise result in inappropriate responses of the clock.
Supplemental Material
Duffield_Supplemental_Material_JBR_Sept_2020 – Supplemental material for Inhibitor of DNA binding 2 (Id2) Regulates Photic Entrainment Responses in Mice: Differential Responses of the Id2-/- Mouse Circadian System Are Dependent on Circadian Phase and on Duration and Intensity of Light
Supplemental material, Duffield_Supplemental_Material_JBR_Sept_2020 for Inhibitor of DNA binding 2 (Id2) Regulates Photic Entrainment Responses in Mice: Differential Responses of the Id2-/- Mouse Circadian System Are Dependent on Circadian Phase and on Duration and Intensity of Light by Giles E. Duffield, Sung Han, Tim Y. Hou, Horacio O. de la Iglesia , Kathleen A. McDonald, Kirk L. Mecklenburg, and Maricela Robles-Murguia in Journal of Biological Rhythms
Footnotes
Acknowledgements
This work was supported by grants awarded to G.E.D. from the National Institute of General Medical Sciences (R01-GM087508) and from the University of Notre Dame (UND), and to H.O.D. from the National Institute of Neurological Disorders and Stroke (R01-NS094211). K.A.M. was a recipient of a UND College of Science Undergraduate Summer Research Fellowship. We thank Theresa Sikorski and staff at the Freimann Life Science Center, UND, for assistance with animal care; Drs Ryan Thummel, Samer Hattar, and Russell Van Gelder for helpful technical advice on retinal histology and pupillometry, respectively; and the anonymous reviewers for helpful suggestions. The authors declare no competing financial interests.
Author Contributions
G.E.D. designed the research; G.E.D., S.H., H.O.I., T.Y.H., K.A.M., K.L.M., and M.R.M. performed the research; G.E.D., K.A.M., and M.R.M. analyzed the data; G.E.D. wrote the article.
Conflict of Interest Statement
The authors have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.