
Abstract
The eye contains a circadian system that acts independently from the master circadian clock located in the brain. This circadian system regulates important physiological functions within the eye. Emerging experimental evidence also indicates that disruption of the ocular circadian clock, or its outputs, negatively affects the overall health of the eye. Although previous studies have investigated the effect of aging on the regulation of circadian rhythms, no study has investigated the effects of aging on the circadian rhythm in the ocular system. The aim of the present study was to investigate how aging affects the circadian rhythm of PER2::LUC bioluminescence in the retina, retinal pigment epithelium (RPE), and cornea. Our data suggest that among the 3 different ocular tissues investigated, the retina appears to be the most affected by aging whereas the RPE and cornea are less affected by aging. Our data, along with studies of other organs and tissues, suggest that reduction in the amplitude of rhythms is probably the most severe effect of aging on the circadian clock.
The mammalian eye contains a full circadian system that acts independently from the master circadian clock located in the brain (McMahon et al., 2014). This circadian system regulates important functions such as visual processing, susceptibility to light-induced photoreceptor damage, neurotransmitter synthesis and release, disk shedding and phagocytosis, corneal thickness, and intraocular pressure (see McMahon et al., 2014, for a recent review). Emerging experimental evidence also indicates that disruption of the circadian clock may negatively affect the overall health of the eye. Bmal1 knockout (KO) mice showed an increase in cataracts and corneal inflammation (Kondratov et al., 2006) and a significant reduction in the number of photoreceptors during aging (Baba et al., 2018). Clock mutant mice developed cataracts during aging (Dubrovsky et al., 2010), and Clock/Npas2 KO mice showed a significant reduction in the thickness of the photoreceptor layer (Baba et al., 2018).
Previous studies have shown that aging affects the circadian rhythms in locomotor activity, suprachiasmatic nucleus of the hypothalamus (SCN), and peripheral organs (Aujard et al., 2001; Yamazaki et al., 2002; Davidson et al., 2008; Sellix et al., 2012; Nakamura et al., 2015; Tahara et al., 2017). No previous study has investigated the effects of aging on the ocular circadian system. The aim of the present study was to investigate how aging affects the circadian rhythm of bioluminescence in the retina, retinal pigment epithelium (RPE), and cornea in PERIOD2::LUCIFERASE mice (PER2::LUC) (Yoo et al., 2004).
PER2::LUC mice raised at the Morehouse School of Medicine (water and food ad libitum, 12 h light and 12 h dark, fluorescent tubes 200-400 lux at cage level) were used in this study at the age of 3 and 12 months. Mice were sacrificed at ZT 8-10, eyes were removed (only one eye from each mouse was used in this study) and placed in iced Hank’s salt solution; then the retina, the RPE-choroid, and the cornea were carefully separated under a dissecting microscope (see Baba et al., 2010; 2015). The retina and eye cup containing the RPE-choroid were flattened by 4 radical incisions and placed on the culture membrane (Millicell-CM, Millipore, Billerica, MA) in a 35-mm Petri dish with 1.2 mL of 199 medium (Cambrex, Walkersville, MD) containing 0.1 mM D-Luciferin K salt (Molecular Imaging Products, Bend, OR), 10 mm HEPES (pH 7.2), penicillin/streptomycin cocktail (2.5 mL/L, Gibco, Grand Island, NY), and B27 (2%; Gibco) (see Baba et al., 2010 and 2017 for details about the specific culture conditions for each tissue). Corneas were prepared and cultured in the same manner as previously described (Baba et al., 2015). Dishes were sealed and kept at 37 °C. Because culture preparation of ocular tissues can be affected by the time of the day at which the cultures are prepared (Evans et al., 2015), all of the cultures were prepared under fluorescent light between ZT 8 and 10. All the procedures were performed in accordance with the National Institutes of Health Guide on the Care and Use of Laboratory Animals and the ARVO Statement for the Use of Animals in Ophthalmic Vision Research and were approved by the Institutional Animal Care and Use Committees of Morehouse School of Medicine. The bioluminescence emitted from the cornea, retina, and RPE-choroid was measured for 1 min at 10-min intervals using a Lumicycle (Actimetrics, Wilmette, IL). Bioluminescence recordings were detrended using a 24-h moving average-subtraction method and smoothed by a 2-h moving average. Daily peaks were identified by Origin software (Origin Lab, Northampton, MA), and the period was calculated from the slope of a linear regression line fitted to circadian peak phases (Baba et al., 2010). The daily amplitude of the circadian rhythm was calculated from the peaks to the median line of wave up to 7 days. The value of rhythmic power was calculated by the fast Fourier transform (FFT) with Lumicycle analysis program (Ruan et al., 2012), which was used to determine the dominant periodicity in the data fit by the cosine function with the highest amplitude (Herzog et al., 2014).
A circadian rhythm in PER2::LUC bioluminescence was observed at both ages and in all the tissues investigated (Fig. 1A), but all tissues from old (12-month-old) mice displayed a significant decrease in the amplitude of the PER2::LUC bioluminescence rhythm (p < 0.001 for all tissues, 2-way ANOVA, n = 6-10). The phase of PER2::LUC rhythm in the retinas of old mice was substantially advanced (Fig. 1B) (CT 10.52 ± 0.32 h, p < 0.01, t test, n = 8) relative to young (3-month-old) mice (CT 19.10 ± 0.14 h, n = 9). The phase of RPE bioluminescence circadian rhythm in old mice was slightly but significantly delayed (approximately 1.5 h) with respect to what was observed in the young mice (3 months CT 13.83 ± 0.23 h vs. 12 months CT 15.23 ± 0.48 h; p < 0.05, t test, n = 6-8). No phase change was observed in cornea (n = 6-10). The period of the PER2::LUC bioluminescence rhythm in the retinas of old mice was approximately 2 h longer than in young mice (Fig. 1C) (3 months 23.98 ± 0.16 h vs. 12 months 25.78 ± 0.27 h). Interestingly, the power spectrum analysis indicated that the rhythmicity of PER2::LUC rhythm in the cornea was less robust in old mice than in young mice (Fig. 1D) (p < 0.01, t test).
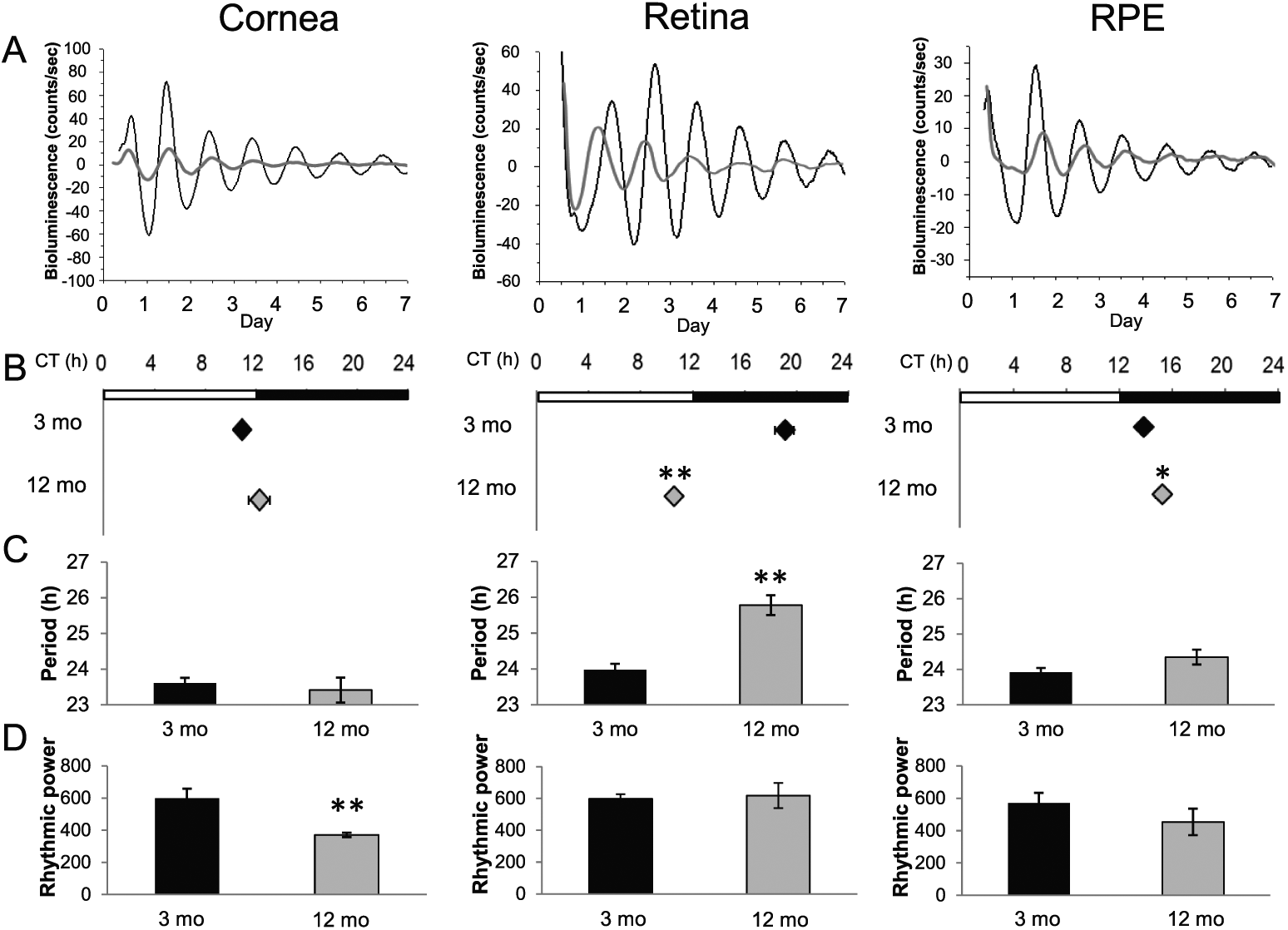
(A) Representative PER2::LUC bioluminescence rhythm in the cornea, retina, and RPE dissected from young (3 months old; black lines) and old (12 months old; gray lines) mice. (B) PER2::LUC rhythm of retinas in old mice showed an approximately 8.5-h phase advance, and RPE showed a slight delay in old mice. Black diamonds represent data obtained from young mice and gray diamonds represent data obtained from old mice (mean ± SEM, n = 6-10, *p < 0.05, **p < 0.01, t test). White and black bars above the plots indicate the light-dark cycle to which mice were exposed before preparation of organ cultures. (C) The period of PER2::LUC rhythm recorded from retinas in old mice was approximately 2 h longer than that observed in young mice (mean ± SEM, n = 6-10, **p < 0.01, t test). Black bars represent young mice and gray bars represent old mice. (D) The cornea was the only tissue in which the rhythmic power of PER2::LUC circadian rhythm was affected by aging (mean ± SEM, n = 6-10, **p < 0.01, t test). Black bars represent young mice and gray bars represent old mice.
Overall, we found that aging affects the circadian rhythms of ocular tissues. The most notable effects can be summarized as follows: (1) The amplitude of PER2::LUC bioluminescence circadian rhythm showed a significant decrease in all three tissues investigated; (2) the retina and the RPE showed a significant change in the phase of the circadian rhythm; (3) in the retina, the period of the circadian rhythm was longer in old mice; and (4) the circadian rhythm was less robust in corneas obtained from old mice.
Our data agree with previous studies which reported that amplitudes of circadian rhythms in the SCN and other tissues decline during aging (Nakamura et al., 2011; Nakamura et al., 2015; Aujard et al., 2001). However, progressive desynchronization of oscillators among SCN neurons, not a reduction in clock function, is thought to be the underlying cause of age-dependent reductions in amplitude (Nakamura et al., 2015). A decline in amplitude of the bioluminescence rhythm of xiphoid cartilage explanted from 20- to 24-month-old PER2::LUC mice and intervertebral disk from 12-month-old PER2::LUC mice also showed a damped circadian amplitude (Gossan et al., 2013; Dudek et al., 2017). Interestingly, the amplitude of the bioluminescence rhythm of xiphoid cartilage could be restored to the level observed in young mice by treatment with dexamethasone, thus suggesting that aging may affect the mechanism mediating the synchronization among the cells (Gossan et al., 2013). An additional study showed that cultured fibroblasts obtained from young and old human subjects did not show any changes in the phase and period of Bmal1::LUC rhythm. However, the Bmal1::LUC circadian rhythm of fibroblasts obtained from young individuals was shorter and phase-shifted when cells were treated with serum obtained from old individuals (Pagani et al., 2011). Hence, these studies may suggest that aging does not affect the capability of the cells to produce a circadian oscillation, but rather aging may prevent the capability of the cells within a tissue to remain synchronized. However, we must be cautious with this interpretation since it is possible that the age-related decrease in the amplitude of bioluminescence rhythms could be due to age-dependent decline in the efficiency of luciferase enzyme.
We found that the period and phase of the retinal circadian PER2::LUC rhythm were significantly affected by aging, with a longer period and a phase advance in the old mice. While this period-phase relationship would not be predicted from classic circadian entrainment theory, it is consistent with previous investigations in the mouse (Nakamura et al., 2015; Yamazaki et al., 2002; Sellix et al., 2012). In this context, it is important to note that these effects are observed in the peripheral oscillators and not in the SCN, in which aging causes a phase delay (Asai et al., 2001; Sellix et al., 2012; Davidson et al., 2008; Nakamura et al., 2015; Yamazaki et al., 2002). In contrast, aging did not affect the phase and period of PER2::LUC rhythm in the mouse cornea, consistent with data from rat corneas (Yamazaki et al., 2002). However, we found that aging increased the interculture variability in the PER2::LUC rhythm (Fig. 1D). Previous studies have reported age-related loss of corneal endothelial cells (Bourne and McLaren, 2004; Gipsen, 2013), and we have previously shown that in the cornea the PER2::LUC bioluminescence signal is mostly emitted by the corneal epithelium and endothelium (Baba et al., 2015). Hence, it is possible that the increased variability of the PER2::LUC rhythm in this tissue is due to changes in the PER2::LUC rhythmicity between the corneal epithelium and endothelium. Finally, we observed that the effect of aging on the RPE is modest, thus suggesting that the circadian clock in this tissue is less sensitive to the aging process.
Previous investigations have shown that many retinal functions are affected by aging. In the mouse, visual processing steadily decreases during aging (Gresh et al., 2003; Baba et al., 2012), photoreceptor viability is also reduced (Gresh et al., 2003; Baba et al., 2009; Gianesini et al., 2016), and dysfunction in the circadian clock leads to photoreceptor loss (Baba et al., 2018). Therefore, it is possible to speculate that the changes in the retinal circadian rhythmicity that occur during aging may negatively influence photoreceptors’ viability during aging.
In conclusion, our study suggests that among the 3 different ocular tissues investigated, the retina appears to be the most affected by aging, whereas the RPE and cornea are less affected by aging. Our data are consistent with previous studies in other organs and/or tissues (Davidson et al., 2008; Sellix et al., 2012; Gossan et al., 2013; Nakamura et al., 2015; Dudek et al., 2017), which have shown that reduction in the amplitude of the rhythms is probably the most severe effect of aging on the circadian clock.
Footnotes
Acknowledgements
This work was supported by grants from the National Institutes of Health: EY022216, EY026291, and T-32 HL103104 to G.T.; GM116760 to K.B.; and 5U54NS083932, S21MD000101, G12-RR03034, and U54RR026137 to the Morehouse School of Medicine. Thanks are also due to Dr. Zach Hall for helpful comments and suggestion on the previous version of the manuscript.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.