
Abstract
Like two dancers, the circadian clock and cell cycle are biological oscillators engaged in bidirectional communication, resulting in circadian clock–gated cell division cycles in species ranging from cyanobacteria to mammals. The identified mechanisms for this phenomenon have expanded beyond intracellular molecular coupling components to include intercellular connections. However, detailed molecular mechanisms, dynamics, and physiological functions of the circadian clock and cell cycle as coupled oscillators remain largely unknown. In this review, we discuss current understanding of this connection in light of recent findings that have uncovered intercellular coupling between the circadian clock in Paneth cells and the cell cycle in intestinal stem cells via WNT signaling. This extends the impact of circadian rhythms regulating the timing of cell divisions beyond the intracellular domain of homogenous cell populations into dynamic, multicellular systems. In-depth understanding of the molecular links and dynamics of these two oscillators will identify potential targets and temporal regimens for effective chronotherapy.
Molecular Mechanisms of Circadian Rhythms
Most organisms exhibit circadian rhythms, reflecting an internal biological clock with an approximate period of 24 h, to anticipate and align their physiological functions with daily environmental changes. Autonomous oscillations of circadian rhythms are generated at single-cell level by transcriptional-translational feedback loops (TTFLs), which consist of positive and negative elements (Hurley et al., 2016; Lowrey and Takahashi, 2011) (Fig. 1A). In mammals, the positive elements consist of heterodimeric basic helix loop helix-Per/Arnt/Sim (bHLH-PAS) transcription factors, Circadian Locomotor Output Cycles Kaput (CLOCK) and Brain and Muscle ARNT-Like 1 (BMAL1), which regulate the expression of negative elements, Period (Per1, Per2, and Per3) and Cryptochrome (Cry1 and Cry2) genes. PER and CRY proteins form complexes and directly inhibit CLOCK/BMAL1 transcriptional activity and repress their own mRNA expression forming a time-delayed negative feedback loop. In addition, CLOCK/BMAL1 controls the expression of nuclear receptors, Retinoic acid–related Orphan Receptor (ROR) and Rev-erbα, whose proteins activate and repress the expression of Bmal1, respectively, forming additional positive and negative feedback loops.
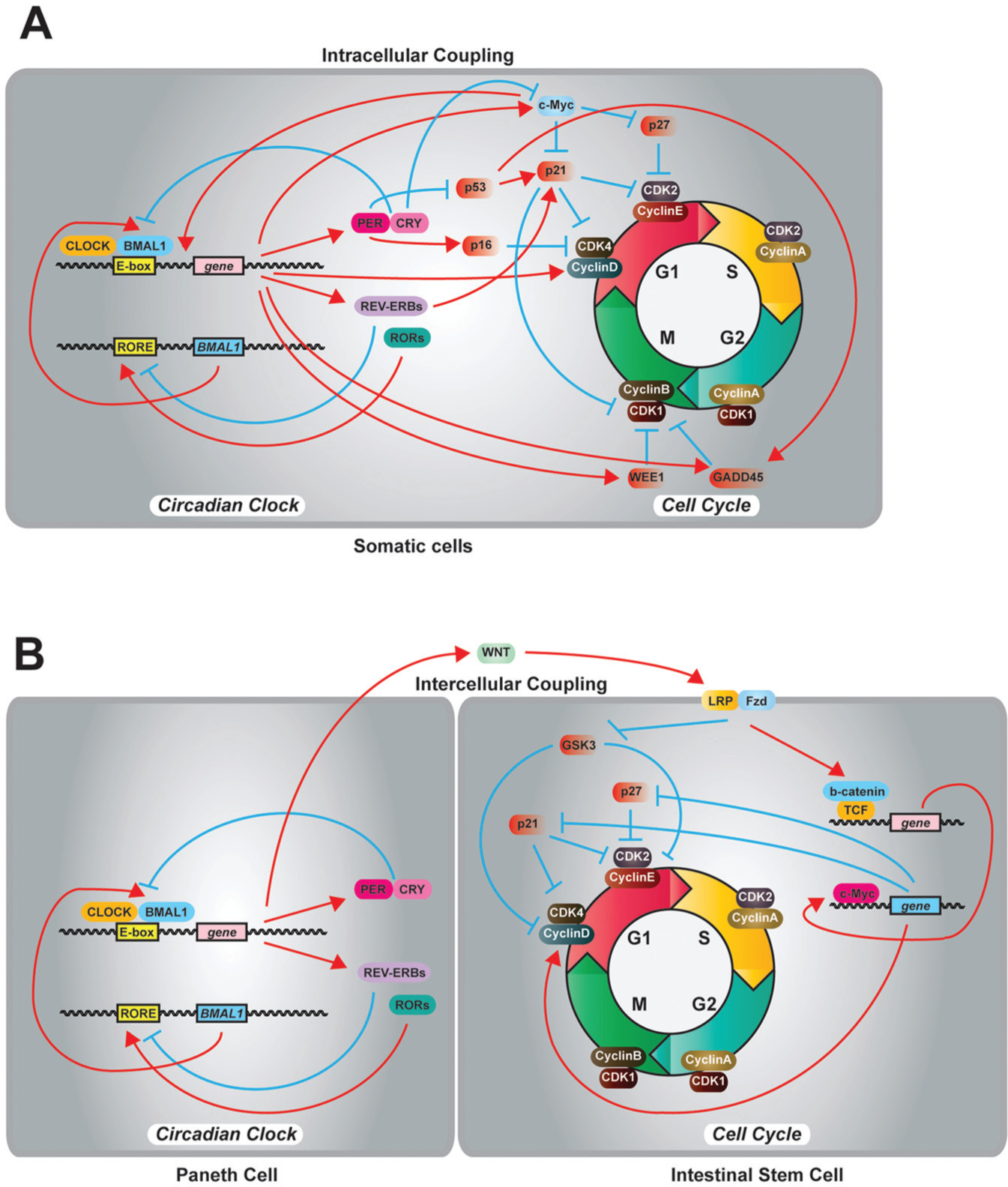
Molecular connections between the circadian clock and the cell cycle. (A) In mammalian somatic cells, the circadian clock gates the cell cycle by intracellular molecular connections. (B) The circadian clock in Paneth cells regulates the timing of divisions in intestinal stem cells and progenitor cells through rhythmic secretion of WNT.
Efficient operation of the above interlocked feedback loops that sustain autonomous circadian oscillations requires posttranslational regulation of the core clock proteins. Casein Kinase 2α (CK2α) rhythmically phosphorylates BMAL1, which determines the rhythmic accumulation of CLOCK/BMAL1 in the nucleus (Kondratov et al., 2003; Tamaru et al., 2009). Casein Kinase 1ε (CK1ε) phosphorylates CRY1, CRY2, and BMAL1, and CKIε-dependent phosphorylation of BMAL1 positively regulates its transcriptional activity (Eide et al., 2002). Importantly, the negative elements undergo extensive phosphorylation that regulates the function and degradation of these proteins. PER2 in mammals and its analogous protein, FREQUENCY (FRQ) in Neurospora crassa, have over 21 and 75 phosphorylated residues, respectively, which regulate the proteins’ half-life, subcellular localization, and circadian period (Baker et al., 2009; Vanselow et al., 2006). Specifically, CK1ε and CK1δ phosphorylate PER proteins and regulate subcellular localization and subsequent ubiquitin-mediated degradation (Akashi et al., 2002; Lee et al., 2009; Vanselow et al., 2006). Phosphorylated PER proteins by CK1ε/δ become targets of the E3 ubiquitin ligase complex that contains beta-transducin repeat–containing protein 1 (β-TrCP1) (Reischl et al., 2007). F-Box and Leucine Rich Repeat Protein 3 (FBXL3) forms a Skp1-Cul1-F-box protein (SCF) E3 ligase complex that polyubiquitinates the CRY proteins for degradation by the proteasomes (Busino et al., 2007; Godinho et al., 2007; Siepka et al., 2007). In contrast, FBXL21, a paralog of FBXL3, forms a SCF E3 ligase complex and stabilizes CRY by protecting it from FBXL3-mediated degradation in the nucleus while destabilizing it in the cytoplasm (Hirano et al., 2013; Yoo et al., 2013). REV-ERBα is ubiquitinated by two E3 ligases, ADP-Ribosylation Factor Binding Protein 1 (ARF-BP1) and Protein Associated with MYC (PAM), which leads to subsequent degradation of REV-ERBα (Yin et al., 2010). BMAL1 is also ubiquitinated by another E3 ligase, Ubiquitin Protein Ligase E3A (UBE3A), and RNA interference (RNAi) of UBE3A disrupts circadian rhythmicity in NIH3T3 cells (Gossan et al., 2014). These findings demonstrate that core clock protein-specific ubiquitin-mediated degradation is important for circadian rhythms. Intriguingly, recent findings from the Neurospora circadian clock have shown unexpected circadian oscillations in the absence of F-box and WD40 repeat-containing protein 1 (FWD-1), an ortholog of β-TrCP1, which determines the stability of the negative element, FRQ (Larrondo et al., 2015). This revealed that there are distinct phosphorylation events that determine the circadian period independent of half-life of FRQ. For more information on detailed molecular underpinnings of circadian rhythms, we refer to comprehensive reviews (Hurley et al., 2016; Lowrey and Takahashi, 2011).
Intracellular Molecular Links between the Cell Cycle and the Circadian Clock
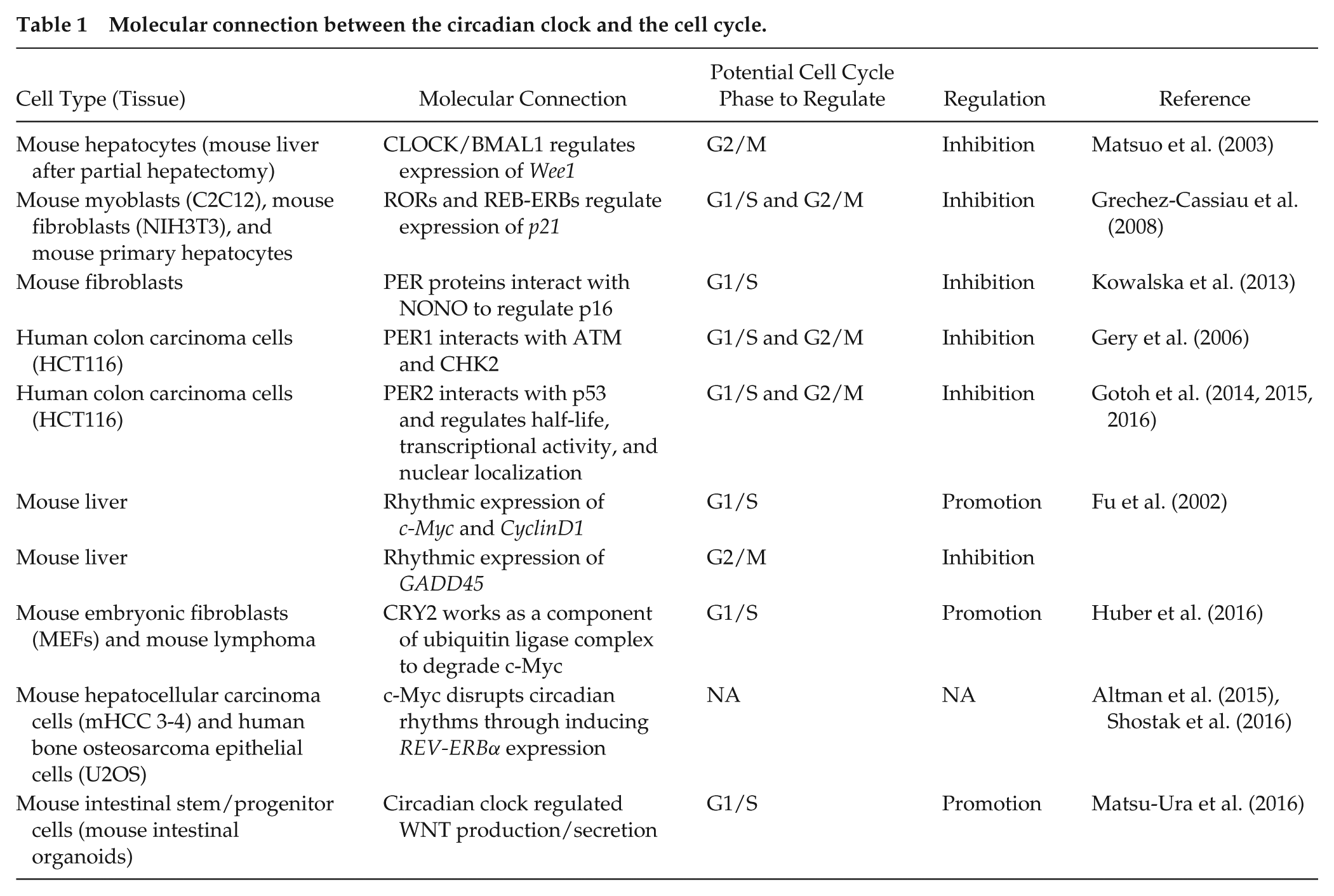
Previously, several circadian clock–regulated cell cycle components have been identified. Matsuo et al. (2003) reported that the expression of a G2/M checkpoint kinase, Wee1, is directly regulated by the heterodimeric circadian clock transcription factor, CLOCK/BMAL1, resulting in rhythmic gene expression and WEE1 kinase activity, which are abolished in Cry-deficient mice. In contrast, RORs and REV-ERBα/β antagonistically regulate the rhythmic expression of p21, a cyclin-dependent kinase inhibitor, which demonstrates that the circadian clock regulates G1/S transition in addition to G2/M (Grechez-Cassiau et al., 2008). Circadian gene expression of p21 is abolished in Bmal1-deficient mice with increased abundance of p21 and delayed cell proliferation (Grechez-Cassiau et al., 2008). Furthermore, PER proteins interact with a nuclear protein, NONO, to regulate rhythmic binding and activation of another cyclin-dependent kinase inhibitor, p16, which shows an additional link to the G1/S transition (Kowalska et al., 2013). Intriguingly, PER1 physically interacts with the checkpoint kinase proteins, Ataxia Telangiectasia Mutated (ATM) and Checkpoint Kinase 2 (CHK2), and overexpression of PER1 leads to a reduction of cell growth and increased susceptibility to DNA damage–induced apoptosis, which reveals a connection between circadian rhythms and DNA damage response (Gery et al., 2006). However, recent findings indicate that the cell cycle machinery also influences the circadian clock. Bieler et al. (2014) demonstrated that the cell division cycle influences period and phase of the circadian clock in NIH3T3 fibroblasts, and DNA replication is necessary for rhythmic access of the frq promoter via the histone chaperon, FACT complex, in N. crassa(Liu et al., 2017).
Importantly, core circadian clock elements regulate key components that control cell proliferation and tumorigenesis. Gotoh et al. reported a series of studies of PER2 interaction with a tumor suppressor, p53. The studies revealed that the physical interaction between PER2 and p53 results in (1) stabilization of p53 from Murine Double Minute-2 (MDM2)–mediated ubiquitination and degradation (Gotoh et al., 2014), (2) inactivation of the transcriptional activity of p53 (Gotoh et al., 2015), and (3) nuclear translocation of p53 in human colon cancer HCT116 cells (Gotoh et al., 2016). A proto-oncogene, c-Myc, and its downstream genes, Cyclin D1 and a tumor suppressor, Growth Arrest and DNA-Damage-Inducible 45 (GADD45), show rhythmic gene expression in mouse liver (Fu et al., 2002). The expression of these genes was altered in the Per2 mutant mouse (mPer2m/m) and showed dramatically increased expression of c-Myc mRNA, which correlates with radiation-induced tumorigenesis in mPer2m/m mice (Fu et al., 2002). In addition, CRY2 regulates the stability of c-Myc by promoting the ubiquitination and degradation of c-Myc (Huber et al., 2016). To add to this complexity, overexpression of c-Myc disrupts circadian rhythms by inducing REV-ERBα, which reduces the expression of Bmal1 (Altman et al., 2015; Shostak et al., 2016), establishing a bidirectional communication between circadian rhythms and cell proliferation. Together, the aforementioned molecular connections (summarized in Table 1) orchestrate intracellular coupling of the circadian clock and the cell cycle in mammalian somatic cells.
Molecular connection between the circadian clock and the cell cycle.
Dysregulated cellular proliferation is a characteristic property of cancer. Oscillations of circadian clock genes were reported in cancer cell lines including osteosarcoma cells (U2OS) (Hughes et al., 2009), breast cancer cells (MCF10A) (Xiang et al., 2012), and colorectal cancer cells (HCT116 and Caco2) (Gotoh et al., 2016; Moore et al., 2014). In contrast, it has been shown that circadian clock–related genes are impaired in most human cancers, suggesting that cancer cells target the circadian clock machinery to achieve uncontrolled growth and proliferation (Davidson et al., 2006). In fact, the number of rhythmic genes is dramatically reduced in cancers and immortalized cell lines cultured in vitro (percentage of rhythmic genes: 1.5% in U2OS [Krishnaiah et al., 2017]; 2.6% in NIH3T3 [Menger et al., 2007]; and 1.9% in Rat-1 [Duffield et al., 2002]) compared with liver and other organs (10%-40%) (Panda et al., 2002; Vollmers et al., 2009; Zhang et al., 2014). Although the difference in the number of rhythmic genes between cell lines and mouse organs may be due to differences in conditions in vitro and in vivo, these results suggest a disruption of the molecular clockworks in cancer and immortalized cells. In 2007, the International Agency for Research on Cancer (IARC) categorized shiftwork that involves circadian disruption as carcinogenic to humans (Straif et al., 2007). Per gene expression is associated with tumor development in γ-irradiated mice (Fu et al., 2002) and human cancers (Zhao et al., 2014). BMAL1 induces apoptosis in pancreatic cancer cells via p53 signaling (Jiang et al., 2016), and downregulation of Bmal1 expression results in increased cell proliferation and tumor growth of colon cancer (Zeng et al., 2010). This evidence supports the view that defective circadian clock gene expression and disruption of circadian rhythms correlate with tumor development and tumor progression in various human cancers (Savvidis and Koutsilieris, 2012).
The Circadian Clock in Adult Stem Cells and Its Connection to the Cell Cycle
Many of the previous studies used cancer or immortalized cell lines to investigate molecular coupling between the circadian clock and cell cycle/proliferation in the mammalian system. However, it is important to note that adult stem cells are one of the major proliferating cell types in our body, and the circadian clock appears to operate differently depending on the context of stem cells. Therefore, it is critical to investigate the roles of circadian rhythms in stem cell regeneration and proliferation. Yagita et al. (2010) reported the lack of circadian rhythms in mouse embryonic stem cells and demonstrated the initiation of circadian rhythms during differentiation. In contrast, human epidermal stem cells possess canonical TTFL of clock components with similar robustness as differentiated keratinocytes, with stemness, differentiation, epidermal aging, and tumorization regulated by the circadian clock (Janich et al., 2011; Janich et al., 2013). The circadian clock gates the cell cycle in epidermal stem cells, which controls the timing of S-phase at around the late afternoon to provide protection against DNA damage from UV irradiation (Geyfman et al., 2012; Plikus et al., 2013). In addition, quiescent neural stem/progenitor cells in the subgranular zone of adult mouse hippocampus possess rhythmic Per2 gene expressing cells and demonstrate time-of-day-dependent proliferation (Bouchard-Cannon et al., 2013). Interestingly, both BMAL1 and PER2 levels are strongly suppressed in the stem cells during the cell cycle, which releases the stem cells from inhibitory effects of cell cycle progression by the circadian clock (see Table 1) (Bouchard-Cannon et al., 2013).
In contrast to the aforementioned direct/intracellular coupling and regulation of stem/progenitor cell proliferation by the circadian clock, adult stem cells are also regulated indirectly by circadian rhythms. Hematopoietic stem cells and progenitor cells (HSPCs) circulate in the bloodstream, and the release of HSPCs from bone marrow is rhythmically regulated by noradrenaline secretion from the sympathetic nervous system that is under the control of the master clock (Mendez-Ferrer et al., 2008). Drosophila intestinal stem cells (ISCs) have a circadian rhythm of PER protein expression, and RNAi of per gene abolishes the daily proliferation rhythm of ISCs in damaged intestine (Karpowicz et al., 2013). Importantly, the daily proliferation rhythm in Drosophila ISCs is disrupted by enterocyte-specific per gene knockdown by RNAi (Karpowicz et al., 2013), suggesting indirect regulation of the cell cycle in ISCs via enterocytes. Recently, we reported a novel intercellular coupling mechanism between the circadian clock and the cell cycle in mouse enteroids (intestinal organoids), which possess ISCs, progenitor cells (PCs), and differentiated epithelial cells including Paneth cells. First, we noticed that the circadian rhythm of PER2 expression is weak or absent in the dividing ISCs/PCs compared with terminally differentiated intestinal epithelial cells in mouse enteroids (Matsu-Ura et al., 2016). Then we discovered that the circadian clock in Paneth cells regulates rhythmic WNT expression and secretion, which controls the circadian clock–gated cell divisions of ISCs and PCs, establishing an intercellular connection between these two oscillators (Fig. 1B). These findings indicate that tissue-specific adult stem cells in different tissues are in different stages of circadian clock development, which results in distinct circadian clock phenotypes depending on the types of adult stem cells.
WNT Signaling in Adult Stem Cells and Its Interaction with the Circadian Clock
WNT signaling is known to regulate the self-renewal and differentiation of various adult stem cells. In the small intestine, secreted WNT from Paneth cells promotes self-renewal of ISCs (Clevers et al., 2014). The activity of WNT is meticulously controlled for tissue patterning in the stomach. The homeodomain transcription factor, BarH-Like Homeobox 1 (BARX1), is abundantly expressed in embryonic mesenchymal cells in the stomach and regulates the expression of WNT inhibitor, Secreted Frizzled Related Protein 1 (SFRP), resulting in the development of gastric epithelium (Kim et al., 2005). In addition, gastric and colon cancers are associated with abnormality of canonical WNT signaling (Polakis, 2000; Vogiatzi et al., 2007). The importance of WNT signaling for self-renewal of adult stem cells in the gastrointestinal tract is also highlighted by the requirement of exogenous WNT for the growth and maintenance of organoid cultures from mammalian taste bud (Aihara et al., 2015), stomach (Barker et al., 2010), intestine (Sato et al., 2011), and colon (Sato et al., 2011). As well, WNT signaling is critical for the niche of adult stem cells in other tissues. Adult rat hippocampal neural progenitor cells (AHPs) express various types of Wnt and Frizzled (Fzd) genes, and WNT signaling maintains multipotency of AHPs and controls their differentiation (Wexler et al., 2009). Identical to AHPs, epidermal stem cells self-renew by autocrine WNT (Lim et al., 2013). WNT signaling is essential for proper embryonic cardiac differentiation, progenitor expansion, and myocardial growth (Klaus et al., 2007). Modulation of WNT activity enhances directed cardiac differentiation of human pluripotent stem cells (Lian et al., 2013), and WNT signaling is suggested to increase cardiac repair after ischemic damage of myocardium and epicardium (Duan et al., 2012). These results suggest that the link between the circadian clock and cell cycle through WNT signaling has an extensive impact on various adult stem cells.
Interactions of the circadian clock and WNT signaling have been described in other cells, tissues, and organisms. Numerous genes in WNT signaling are under circadian regulation, which highlights the impact of circadian rhythms on WNT signaling. Approximately 50 genes that are involved in WNT signaling demonstrate rhythmic expression profiles in 39 microarray data sets from 19 distinct mammalian tissues (Sotak et al., 2014). ChIP-Seq analysis using mouse liver (Rey et al., 2011) revealed rhythmic occupancy of BMAL1 on the promoters of the canonical WNT signaling pathway including Wnt10a, β-catenin, Fzd5, Dishevelled Segment Polarity Protein 2 (Dvl2), and Transcription Factor 3 (TCF3) (Guo et al., 2012), which highlights a direct link between the circadian clock and WNT signaling. Furthermore, a negative WNT signaling regulator, Axis Inhibition Protein 2 (AXIN-2), and WNT antagonist, Dickkopf WNT Signaling Pathway Inhibitor 3 (DKK-3), show rhythmic gene expression in arcuate nucleus in the brain of female Djungarian hamster (Boucsein et al., 2016).
Genetic and physiological disturbances of circadian rhythms affect the expression of genes in WNT signaling. Xenografted cancer cells proliferate faster in nude mice exposed to a “circadian disrupted” constant light condition compared with nude mice exposed to a “normal” 12-h light-dark cycle, secondary to increased expression of WNT10A in dermal fibroblasts due to disruption of the circadian clock (Yasuniwa et al., 2010). Furthermore, overexpression of Bmal1 increased the expression of β-catenin and cell proliferation in NIH3T3 cells (Lin et al., 2013). In contrast, shRNA knockdown of Bmal1 decreased the expression of genes in the canonical WNT pathway, Axin2, Wnt10a, β-catenin, Fzd5, Dvl2, and TCF3, during adipogenesis of 10T1/2 mouse embryonic cells (Guo et al., 2012). Last, in zebrafish, mRNAs of wnt1, wnt3a, and wnt5a were down-regulated in neurons of per1 mutants in constant dark conditions (Huang et al., 2015). Together, these data indicate a robust connection between the circadian clock and WNT signaling, which widens the impact of circadian rhythms in cell proliferation, self-renewal and differentiation of stem cells, and tissue patterning.
Conclusion
Three and a half centuries following Christiaan Huygens’ classic description of coupled oscillations between two pendulum clocks in 1665 (Huygens, 1893), coupled oscillators continue to be identified throughout the natural world. In biology, coupled oscillators generate diverse dynamic behaviors such as the synchronous flashing of fireflies (Blair, 1915), circadian rhythms in the SCN (Barker et al., 2007), frog choruses (Aihara et al., 2014), myocardial cell contractions (Mirollo and Strogatz, 1990), polyrhythmic interactions among critical brain areas during working memory maintenance (Fujisawa and Buzsaki, 2011), and time-of-day proliferation of somatic cells including adult stem/progenitor cells. These observations indicate the ubiquitous nature of coupled oscillators, but molecular mechanisms that determine diverse dynamic behaviors remain largely unexplored. The circadian clock and the cell cycle use both intra- and intercellular coupling mechanisms to establish their connections, which may vary depending on the cell types and multicellular environment.
We recently demonstrated that the circadian clock in Paneth cells regulates rhythmic expression and secretion of WNT, which coordinates the timing of cell divisions of adult stem and progenitor cells resulting in different coupling ratios depending on the inherent frequency of cell cycle times (Matsu-Ura et al., 2016). Importantly, we identified rhythmic intestinal crypt formation following the coordinated cell division cycles, as well as a reduced number of crypts in the small intestine of circadian arrhythmic, Per1/2-double-knockout, mice. These data suggest that the circadian clock–dependent gating of cell division cycles affects proper timing and formation of intestinal crypts. Importantly, our data enable us to hypothesize that gastrointestinal stem and progenitor cells will likely respond differently to chemotherapeutic drugs such as 5-Fluorouracil depending on the time of administration due to clock-gated cell cycle progression.
Unintended damage to healthy stem and progenitor cells remains a major limitation of cancer chemotherapy (Davidson et al., 2006; Savvidis and Koutsilieris, 2012). Further, disruption of circadian rhythms is associated with a higher risk of various human cancers (Savvidis and Koutsilieris, 2012), and the circadian clock is dysregulated in most human neoplasms (Davidson et al., 2006). Therefore, it will be important to characterize cell cycle profiles of different adult stem cells and cancer cells with respect to the circadian cycle and determine whether those cells show time-of-day-specific responses to chemotherapy. This information could then be exploited to design novel treatment regimens to administer chemotherapeutic drugs at the trough of cell divisions of healthy stem/progenitor cells to reduce the risk of unintended damage to healthy cells, without reducing the efficacy of drugs to eliminate cancerous cells. Further investigation of the molecular coupling components and the intimate dance between the cell cycle and the circadian clock in both normal and cancer cells will be critical to effectively use circadian molecular signatures to treat cancer.
Footnotes
Acknowledgements
This work was supported by Department of Interior grant D12AP00005 (to S.R.M. and C.I.H.) and NIH grant 1U19AI116491 (to S.R.M. and C.I.H.). All authors contributed to writing this manuscript and generating figures.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.