
Abstract
The increasing demand for improved agricultural production will require more efficient breeding for traits that maintain yield under heterogeneous environments. The internal circadian oscillator is essential for perceiving and coordinating environmental cues such as day length, temperature, and abiotic stress responses within physiological processes. To investigate the contribution of the circadian clock to local adaptability, we have analyzed circadian period by leaf movement in natural populations of Mimulus guttatus and domesticated cultivars of Glycine max. We detected consistent variation in circadian period along a latitudinal gradient in annual populations of the wild plant and the selectively bred crop, and this provides novel evidence of natural and artificial selection for circadian performance. These findings provide new support that the circadian clock acts as a central regulator of plant adaptability and further highlight the potential of applying circadian clock gene variation to marker-assisted breeding programs in crops.
Plants exhibit a remarkable capacity to adapt to changing environments. Their immobility creates a natural selective pressure to acquire a repertoire of finely tuned responses that modify their physiology in response to the environmental conditions. The challenge is exceedingly high for a plant; they need to respond to a multitude of stresses and environmental conditions while maximizing growth and fitness. Not surprisingly, each stress response elicits a cascade of signaling events that are all coordinated with the daily light-dark and warm-cold cycles on earth. This coordinated response to daily and seasonal changes brought on by the rotation and axial tilt of the earth is facilitated by an internal biological clock. An endogenous circadian clock is present in all three domains of life: eubacteria, archaea, and eukaryotes (Edgar et al., 2012).
The widespread occurrence of endogenous circadian clocks suggests that there is a fitness advantage in entraining internal processes to the external environment. For example, strains of cyanobacteria with circadian periods that match the light/dark cycle will out-compete other strains with altered periods, which demonstrates a fitness advantage when the circadian oscillator resonates with the environment (Ouyang et al., 1998). In the model plant Arabidopsis thaliana, proper clock-regulation of growth confers a fitness advantage as compared with mutants whose endogenous periods do not resonate with the environmental cycle (Dodd et al., 2005; Resco et al., 2009). Because of this benefit of synchrony with the environment, it is predicted that the evolution of circadian clock performance is linked to daily and seasonal fluctuations in the environment. Given the changes in the annual patterns of temperature and photoperiod with latitude (Hut et al., 2013), similar clines in circadian timing would be expected to properly match behavioral or physiological responses. For example, Drosophila melanogaster delays its afternoon activity under warmer temperatures to avoid desiccation, a response that is dependent on altered splicing of the clock gene period (per) (Majercak et al., 1999). In 2 predominant splice forms of D. melanogaster per, latitudinal clines alter temperature compensation of the clock, providing additional buffering to the local climatic environment (Costa and Kyriacou, 1998; Rosato et al., 1997; Sawyer et al., 1997). Similarly, allelic variants that differ in an important transcriptional activation domain of the mammalian circadian rhythm gene Clock vary in relative frequency along a latitudinal cline in nonmigratory blue tit (Cyanistes caeruleus) birds and Chinook salmon (Oncorhynchus tshawytscha) (Johnsen et al., 2007; O’Malley and Banks, 2008). Evidently, divergent selection on circadian clock traits contributes to local adaptation in a variety of organisms.
Studies using natural accessions of Arabidopsis have uncovered associations between hypocotyl growth, light sensitivity, and flowering time with geographical parameters (Maloof et al., 2001; Stenøien and Fenster, 2002; Stinchcombe et al., 2004). However, although circadian period has been significantly associated with altitude, only suggestive correlations of period with latitude and longitude have been reported, possibly due to limited sample size or distribution (Edwards et al., 2005; Michael et al., 2003). A recent study in tomato uncovered both a phase delay and period lengthening of the circadian clock in domesticated varieties that correlate with expanded cultivation at higher latitudes (Muller et al., 2016). Such trait variation among populations across a geographic gradient is often an indicator of natural selection (Endler, 1986). Based on early findings in Arabidopsis, we hypothesized that plants subjected to natural or artificial selection would exhibit variation in circadian clock period across geographical and environmental gradients. To test this, we selected natural populations of Mimulus guttatus (monkeyflower) and elite cultivars of Glycine max (soybean) and assessed circadian period variation using automated leaf movement analyses.
Materials and Methods
Plant Growth Conditions
All plants were grown in Sunshine Redi-earth. M. guttatus was grown under ~90 μmol s-1 m-2 light and soybean under ~300 μmol s-1 m-2 light. A day after transferring plants to the imaging chamber, they were watered once with a 20-20-20 (NPK) fertilizer. Plants were watered daily to prevent any movement due to water loss. Soybean seeds were sown onto damp soil in 2.25” square pots and grown in cycles of 12 h light at 25 °C and 12 h dark at 18 °C. Following emergence of the first trifoliate leaves, plants were transferred to constant light at 25 °C for imaging. Soybeans were tied to stakes just under the cotyledons to keep them upright during imaging. We used a randomized block design with representatives from each maturity group imaged in each run to test for experimental replicate effects. M. guttatus seeds were stratified in the dark at 4 °C in water for 1 week, planted in ½” 1.27-cm polyvinyl chloride (pvc) coupling tubes and grown in cycles of 12 h light at 20 °C and 12 h dark at 16 °C. Plants were moved into the imaging room at the emergence of the first set of true leaves and imaged in constant light at 20 °C. All annual populations were imaged in one experiment and the perennial populations were imaged in one experiment with 2 overlapping annual populations included in the perennial experiment to test for experimental effects and allow us to compare across experiments. Seed source and number of lines imaged for each population can be found in Supplemental Table S1.
Circadian Period Estimation
The camera and imaging platforms and circadian period estimation using TRiP (Suppl. Tables S1-S2) were described previously (Greenham et al., 2015).
Statistical Analysis
To examine the relationship between circadian period and latitude in M. guttatus, we first performed a separate analysis on annuals and perennials. We used a mixed model approach in SAS (PROC MIXED, SAS v9.3) for annual and perennial populations alone using the following model: Period = Population + Line (Population) + latitude + error. We treated latitude as a fixed effect, population and line within population as random effects (Suppl. Table S3). To further detect interactions between habits (annuals vs. perennials) and latitude, we applied the following model to the entire dataset: Period = habits + population (habits) + line (Population) + latitude + habits * latitude + error. We treated population and line as random effects and habits and latitude as fixed effects (Suppl. Table S3). The annual and perennial M. guttatus lines were imaged in 2 independent experiments with 2 annual populations imaged each time. We ran an ANOVA to test for a repeat effect between experiments and did not detect a significant difference. A t-test was used to compare the annual and perennial average period data. To examine the association with climatic variables we ran a PCA analysis using prcomp() in R (RC Team, 2015) (Suppl. Table S4). For the soybean analysis, we performed a generalized linear model in SAS (PROC GLM, SAS v9.3); genotype was treated as a fixed effect and experimental repeat as random effects. We examined the effect of excluding the outliers in Figure 2D using SAS PROC REG and Cook’s distance output values. The Cook’s D for the very short period cultivar is 0.58 and we excluded this line from the regression and still found a significant correlation (r2 = 0.2384, p = 0.0003). Therefore, we decided to keep the outlier in the model to present the full data set. A similar test was performed for the data in Figure 2E and the results were also significant (r2 = 0.22, p = 0.0007).
Results
Annual Populations of Mimulus guttatus Exhibit a Latitudinal Cline in Circadian Period
Mimulus species are an exceptional system for studying ecological adaptations because of their breadth of evolved physiological and developmental responses to diverse habitats, such as grasslands, coastal cliffs, sand dunes, and heavy metal soils (Wu et al., 2008). M. guttatus can be found from northern Mexico to Alaska, with multiple coastal and inland populations that span altitudinal ranges throughout the latitudinal gradient (Kiang and Hamrick, 1978; Wu et al., 2008). We examined the circadian phenotype of M. guttatus populations spanning a wide latitudinal range to address whether the clock is contributing to local adaptation. To preclude variation in altitude (Edwards et al., 2005) from confounding our analysis, we considered only populations occurring at elevations less than 650 m (Fig. 1A; Suppl. Table S1). Plants were entrained to daily cycles of light and temperature (see Materials and Methods) and, following emergence of true leaves, were moved to continuous light and constant temperature for determination of free-running circadian period by leaf movement (Greenham et al., 2015) (Fig. 1B and 1C; Suppl. Table S2). The populations included both perennial and annual forms of M. guttatus classified either by physiological study or confirmed by a chromosomal inversion polymorphism shown to contribute to this life-history transition (Lowry and Willis, 2010; Twyford and Friedman, 2015). Using a mixed model analysis, we found a significant correlation between period and latitude among the annual populations (r2 = 0.36, p < 0.0001; Fig. 1D, Suppl. Table S3). Given the genetic heterogeneity within these populations due to the outcrossing nature of this species, the significance of this correlation is striking. Northerly populations tended to have longer periods than the southern populations (Suppl. Table S2). Interestingly, we did not observe a latitudinal cline in period among perennial populations (Fig. 1F). The overlapping geographic distributions of interfertile annual and perennial ecotypes have led to extensive gene flow between these populations, which are genetically differentiated only by a large chromosomal inversion (Twyford and Friedman, 2015). Reciprocal transplant experiments have shown that traits (e.g., days to flower, survival to flower, flowers produced) contributing to local adaptations that differ between annuals and perennials map to this inversion and a strong selection between inversion loci and the environment is found in near isogenic lines (Hall and Willis, 2006; Lowry and Willis, 2010). Phylogenetic analysis also shows that neighboring populations of M. guttatus are more similar regardless of their life history, and latitudinal variation is associated significantly with the genetic composition of these populations (Twyford and Friedman, 2015). That the latitudinal cline was observed for circadian period among annuals and not perennials suggests that this trait may colocalize with or depend on the inversion and contributes to local adaptation among annuals.
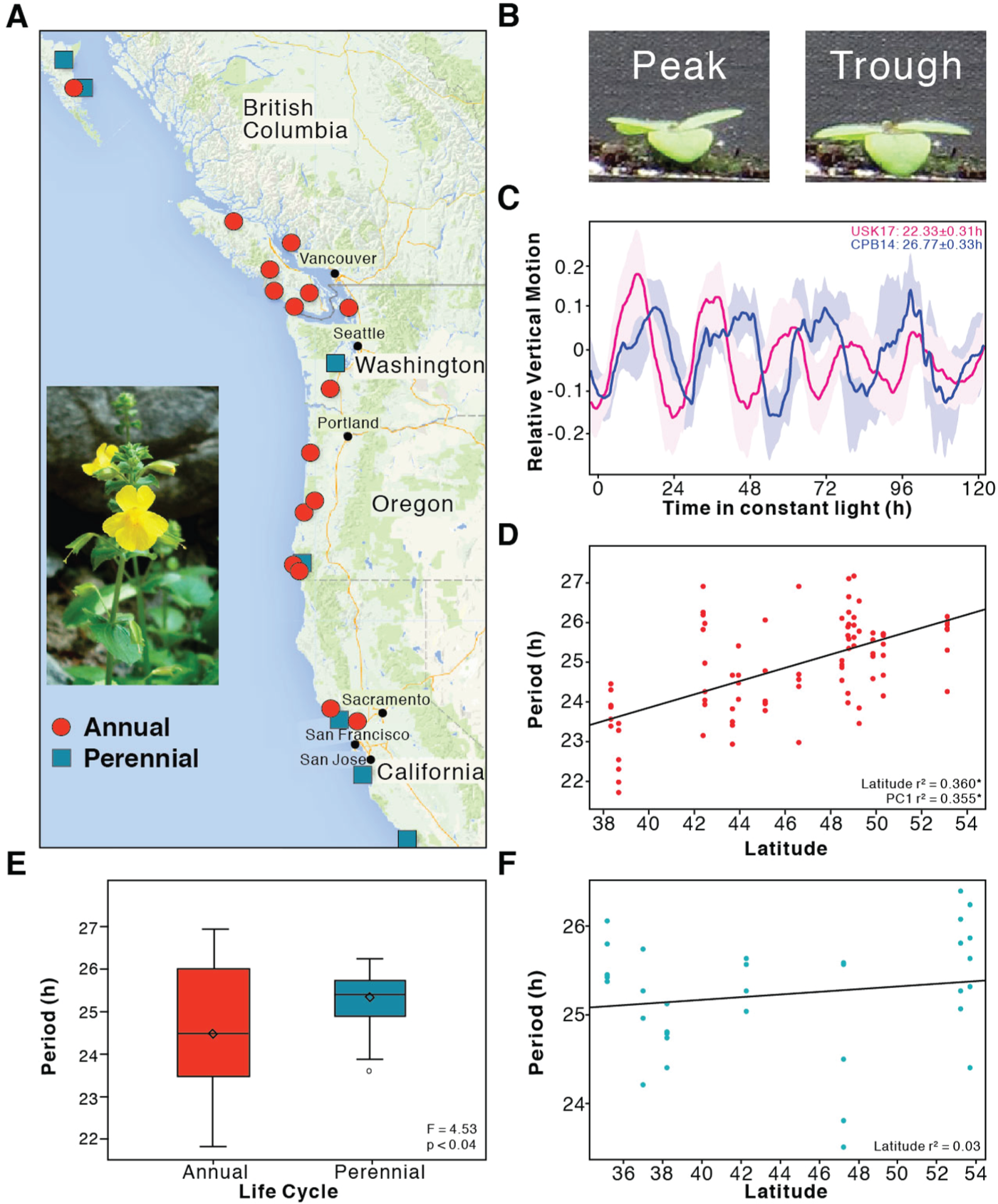
Circadian period is significantly correlated with latitude in annual populations of M. guttatus. (A) Geographic origin of the M. guttatus populations. Red circles represent annual populations and blue squares represent populations with perennial life cycles. (B) Images showing true leaves at peak (left) and trough (right) positions. (C) Examples of leaf movement traces from 2 annual populations. Traces are presented as mean (indicated by the line) and SEM (indicated by the shading). (D) Circadian period is positively correlated with latitude for annual populations growing at <650 m elevation. Data points represent the mean of each line. Solid line represents the regression line. Asterisks indicate statistical significance at p < 0.0001. (E) Effect of life cycle on circadian period. Data are the average period among 5 annual (red) populations (CAV, USK, GBS, RFA and CPB) and 7 perennial (blue) populations (ORO, DAV, WTB, OPB, HOC, WLB, and PCL) at similar latitudes (Suppl. Table S1). (F) Circadian period does not correlate with latitude in perennial populations of Mimulus guttatus. Data points represent the mean of each line. Solid line represents the regression line.
In addition, a significant difference in period was found between annuals and perennials growing at similar latitudes, with a longer period in perennials (Fig. 1E). Perennial M. guttatus populations typically grow in regions with high year-round soil moisture and therefore invest more energy in vegetative growth and flower later than annuals (Hall and Willis, 2006; Lowry and Willis, 2010). One consequence of lengthening the period, seen as a lagging phase under light-dark cycles, could be to increase the critical photoperiod and so delay flowering, consistent with the late flowering among perennials (Lowry and Willis, 2010). In contrast, annual populations of M. guttatus are found in areas with limited soil moisture that are prone to early seasonal drought (Wu et al., 2008). The leading phase associated with a short period could decrease the critical photoperiod and so accelerate flowering as a mechanism to avoid drought. Drought stress can also accelerate flowering once the critical photoperiod is reached, a process that requires the circadian clock gene GIGANTEA (Riboni et al., 2013; Riboni et al., 2014). Rapid life cycle completion allows many annual populations of M. guttatus to survive in such habitats (Hall and Willis, 2006; Lowry et al., 2008). The relationship between circadian period and flowering time may have necessitated a greater range of periods among annuals to enable sexual reproduction under diverse conditions. This is consistent with previous work showing less variation in the critical photoperiod among perennial compared with annual populations (Friedman and Willis, 2013).
The link between flowering time and circadian clock function is complex and, given the breadth of pathways guided by the circadian clock, other environmental factors, such as photoperiod, temperature and precipitation, are likely important contributors to the variation observed in the annual populations. However, latitude, precipitation, temperature, and other climatic measures covary, making it difficult to disentangle them and establish clear causal relationships without extensive additional experimentation. To test for associations of circadian period with climate, we selected 19 climatic variables from the WorldClim dataset at 10 min resolution (Hijmans et al., 2005) and applied Principal Component Analysis (PCA). PC1 explained 62% of the variation with opposing associations between precipitation and temperature (Suppl. Table S4). Although temperature is a well-established environmental cue entraining the circadian clock (Greenham and McClung, 2012), we know little about the influence of precipitation on clock function. The strong correlation of circadian period with precipitation is particularly enticing, since low soil moisture is a constant threat to annual populations. As precipitation increases with latitude so does period, suggesting that annuals at higher latitude that receive more annual precipitation could increase fitness through later flowering; extension of the vegetative phase would increase biomass accumulation and seed production. Thus, precipitation may be an important driver of circadian period variation among these populations. Clearly, the evolutionary response to drought can be quite rapid. For example, 2 natural populations of the annual Brassica rapa underwent rapid adaptive evolution in response to selection imposed by a series of dry years with severe drought late in the growing season (Franks et al. 2007). Descendants flowered earlier than the ancestors, indicating rapid adaptive evolution in the rates of development and flowering, which allowed the descendants to escape the late-season drought (Franks et al., 2007; Franks et al., 2008; Franks, 2011; Franks et al., 2016).
Elite Cultivars of Soybean Exhibit a Latitudinal Cline in Circadian Period
Our leaf movement analysis provides new support for the hypothesis that fine-tuning the circadian clock is one component of the local adaptability of wild populations of M. guttatus. If the clock contributes to local adaptation and plant performance, we would expect to see similar trait variation in modern crop species that have undergone extensive selective breeding for geographically optimized performance. Several studies have implicated the circadian clock in regulating growth and flowering in soybean. Eight maturity loci (E1 to E8) that alter the time required to reach the final stage of development, several of which are photoperiod and light sensitive, are important targets for yield improvement (Xia et al., 2012). The circadian clock gene GIGANTEA is responsible for a flowering quantitative trait locus corresponding to E2 (Watanabe et al., 2011). In addition, a recent examination of photoperiodic flowering in soybean revealed a correlation between the abundance of the circadian regulated cryptochrome1 blue light receptor GmCRY1a and latitude (Zhang et al., 2008). However, none of these studies have directly assessed the relationship of circadian period—a direct and robust measurement of circadian clock function—with latitude.
To directly assess the relationship of circadian period with latitude in a cultivated crop, we obtained 60 genotypes of soybean from a collection of elite cultivars from 6 distinct maturity groups (Fig. 2A). While M. guttatus populations have been naturally selected for performance, elite soybean cultivars have been artificially selected for optimal yield in specific maturity zones along a latitudinal gradient from southern Louisiana to North Dakota. Soybean cultivars are adapted to the photoperiod in defined latitudinal ranges of 160-240 km, with maturity group designations of 00-000 grown in the north and VIII grown in the south (McWilliams et al., 1999). Each maturity group is further subdivided into early (0.1-0.3), middle (0.4-0.6) and late (0.7-0.9) based on time to physiological maturity. Most soybean cultivars can be classified as either of indeterminate and determinate growth habit; indeterminate genotypes continue vegetative growth throughout reproductive development whereas determinate genotypes cease vegetative growth at the onset of reproductive development (Tian et al., 2010). We limited our analysis to indeterminate types because determinate types are only grown in the most southern maturity zones.
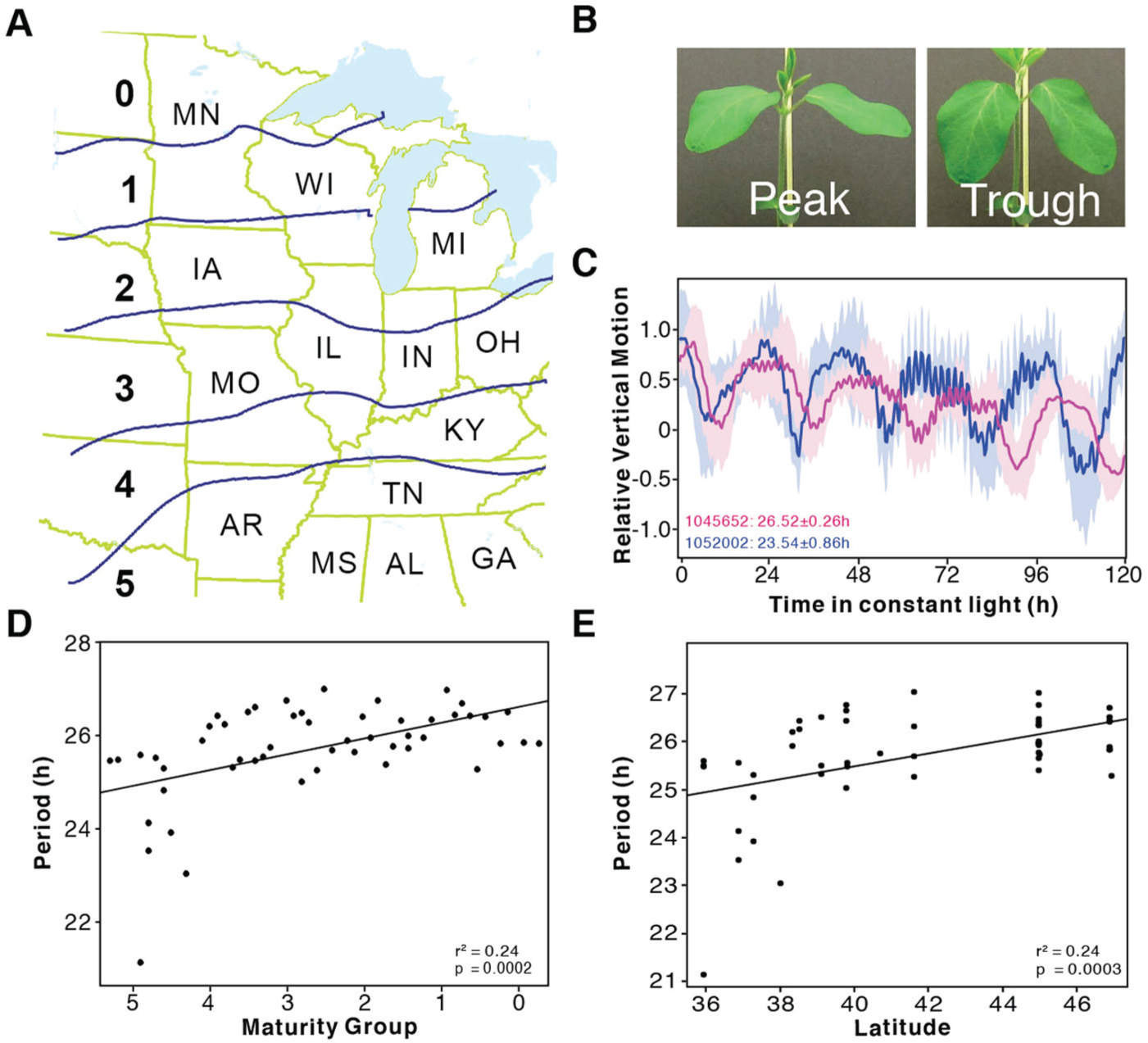
Circadian period is significantly correlated with maturity group in soybean (Glycine max). (A) Map of cultivation ranges of soybean relative maturity (RM) groups. (B) Images showing leaves at peak (left) and trough (right) positions. (C) Examples of leaf movement traces from 2 cultivars. Traces are presented as mean (indicated by a line) and SEM (indicated by shading). (D) Circadian period is significantly correlated with RM group plotted from southern to northern RMs along the x-axis. Solid line represents the regression line. (E) Circadian period is significantly correlated with latitude. Latitude was estimated based on the geographic center of RM plantings. Solid line represents the model fit. All data points are the mean of each line.
Plants were entrained to daily cycles of light and temperature (see Materials and Methods) and, following emergence of the first trifoliate leaves, plants were transferred into constant light and temperature for circadian period estimation by leaf movement (Fig. 2B-C). Consistent with our observations in M. guttatus, the period of these soybean cultivars is significantly correlated with maturity group (r2 = 0.24, p = 0.0002; Fig. 2D, Suppl. Table S5). Because the maturity groups vary along a latitudinal gradient, we also observed a significant positive correlation of period with latitude (defined as the midpoint latitude for the maturity group plantings) (Fig. 2E). This provides strong evidence that circadian clock function varies systematically among maturity groups and contributes to the optimization of fitness of these cultivars along the latitudinal gradient.
Discussion
Results from this study reinforce the role of the circadian clock as a central regulator translating environmental signals into coordinated growth responses, and add to previous studies suggesting latitude-specific selection effects on circadian properties (Hut et al., 2013). In Drosophila, circadian period is correlated with latitude in locomotor activity, eclosion rhythms, and oviposition (Hut et al., 2013; Joshi and Gore, 1999). Among Japanese populations of D. auraria, the amplitude of the oscillator and the phase-response curve both varied with latitude (Pittendrigh and Takamura, 1989). In D. littoralis, the rate at which the circadian rhythm damps out and eclosion becomes arrhythmic increases with latitude (Pittendrigh et al., 1991). A latitudinal cline in allele-length at the OtsClock1b locus was found in kokanee salmon populations in both North America and Russia (Lemay and Russello, 2014). A study of 52 human populations revealed a significant correlation between single nucleotide polymorphisms in circadian-regulated genes and latitude-derived difference between maximum and minimum photoperiod; this suggests that, following migration out of Africa, humans adapted to different latitudes in part through adjustments to their circadian clocks (Forni et al., 2014).
Our data add to these studies, firmly establishing latitudinal clines in circadian period in both wild annual populations of M. guttatus and domesticated and highly bred populations of soybean. By breeding for favorable productivity traits, breeders have been inadvertently selecting for circadian parameters that likely contribute to improved performance in distinct latitudinal zones, as recently found in tomato. Allelic variation of the tomato homolog of Arabidopsis EID1 is responsible for the phase delay, and the EID1 genomic region shows the signature of selection. The EID1 allele of domesticated varieties enhances performance (specifically chlorophyll content) in long days, and this was encountered during cultivation at increasing latitude (Muller et al., 2016). Similarly, natural selection has modified circadian period across the latitudinal range of annual populations of M. guttatus. The lack of a latitudinal cline in perennial populations of M. guttatus supports the model that annual and perennial plants are under divergent selection due to altered fitness trade-offs (Friedman and Rubin, 2015). Annuals must complete their life cycle in one growing season. Annual populations of M. guttatus grow in a range of local environments, including cliff faces and steep hillsides, and frequently experience seasonal droughts, with growing seasons varying from 6 weeks to 5 months. Annuals tend to flower early, which restricts their vegetative growth and therefore limits their seed production. Extensive genetic variation in these populations results in morphological and physiological traits that vary along latitudinal gradients, such as plant height, trichome number and leaf shape (Kooyers et al., 2015). Many of these patterns are discordant with altitudinal trends (Kooyers et al., 2015), and reveal the challenges associated with linking environmental factors and local adaptability. In contrast, if conditions become unfavorable, perennials can reduce their risk by increasing vegetative growth, flowering later, or missing a seed production year (Friedman and Rubin, 2015). Genetic divergence in reproductive timing among M. guttatus populations provides an essential adaptation to their local growing season, as seen with low performance when populations are grown in nonnative sites (Hall and Willis, 2006; Lowry et al., 2008).
The pressures faced by annual populations of M. guttatus are comparable to the challenges associated with the geographic expansion of crop growth during domestication, a process that often results in modifications to photoperiodic flowering (Nakamichi, 2014). The pressure to flower and produce a high yielding seed deposit in a growing season leads to altered circadian clock performance across latitudinal clines through natural and artificial selection. The shared correlation between the circadian period and latitude in natural populations of M. guttatus and soybean highlights the importance of continued investigation of the diversity and genomic basis of adaptation within and among natural populations. Identification of genomic loci in these and other populations of M. guttatus that enable survival in harsh conditions are excellent candidates for marker-assisted breeding in crop plants, such as soybean. We have also described life-history–dependent variation in the circadian clock, which may be used by annual populations of M. guttatus to adapt to harsh conditions. On a broader level, this study adds support to the view that integrating circadian trait information into current breeding programs has the potential to expand the performance of crops in the field by fine-tuning the circadian system for targeted geographic regions. This is especially critical for soybean, where weather trend data from 1994-2013 of in-season temperature and precipitation variation has estimated yield suppression of 30% in the US, with region-based variations from -22% to +9% (Mourtzinis et al., 2015). These findings suggest that local adaptation strategies are essential for continued crop improvement.
Footnotes
Acknowledgements
This work was supported by grants from the National Science Foundation to K.G. (IOS-1202779), J.R.P. (IOS-1202778), and C.R.M, (IOS-0923752 and IOS-1025965) and by a grant from the NSF LiT to J.H.W. (IOS-1024966). We thank Monsanto Company for providing the soybean cultivars and Miya Howell, Shunhong Dai, and Sasha Preuss for helpful discussions.
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.