
Abstract
In mammals, the circadian system is composed of a principal circadian oscillator located in the suprachiasmatic nucleus (SCN) and a number of subordinate oscillators in extra-SCN brain regions and peripheral tissues/organs. However, how the time-keeping functions of this multiple oscillator circuit are affected by aberrant lighting environments remains largely unknown. In the present study, we investigated the effects of chronic light exposure in the middle of the night on the circadian system by comparing the mice housed in a 12:4:4:4-h L:DLD condition with the controls in 12:12-h L:D condition. Daily rhythms in locomotor activity were analyzed and the expression patterns of protein products of clock genes Period1 and Period2 (PER1 and PER2) were examined in the SCN and extra-SCN brain regions, including the dorsal striatum, hippocampus, paraventricular nucleus (PVN), and basolateral amygdala (BLA). Following 2 weeks of housing in the L:DLD condition, animals showed disturbed daily rhythms in locomotor activity and lacked daily rhythms of PER1 and PER2 in the SCN. In the extra-SCN brain regions, the PER1 and PER2 rhythms were affected in a region-specific pattern, such that they were relatively undisturbed in the striatum and hippocampus, phase-shifted in the BLA, and abolished in the PVN. In addition, mice in the L:DLD condition showed increased anxiety-like behaviors and reduced brain-derived neurotropic factor messenger RNA expression in the hippocampus, amygdala, and medial prefrontal cortex, which are brain regions that are involved in emotional regulation. These results indicate that nighttime light exposure leads to circadian disturbances not only by abolishing the circadian rhythms in the SCN but also by inducing misalignment among brain oscillators and negatively affects emotional processing. These observations serve to identify risks associated with decisions regarding lifestyle in our modern society.
Virtually all organisms, including humans, have evolved a circadian timing system to adapt to the daily fluctuations in the environmental light condition (Klein et al., 1991). In mammals, the circadian system is composed of a principal circadian oscillator located in the suprachiasmatic nucleus (SCN) of the hypothalamus and subordinate oscillators in extra-SCN brain regions and peripheral tissues and organs (Mendoza and Challet, 2009; Dibner et al., 2010; Mohawk et al., 2012; Menaker et al., 2013). The self-sustained circadian oscillation is driven by feedback loops containing circadian clock genes, such as Period1 (Per1) and Per2, and their protein products (Reppert and Weaver, 2002; Partch et al., 2014). The output signals from the SCN keep the extra-SCN oscillators synchronized with each other and with the environment light/dark (LD) cycle, ensuring that each physiological, biochemical, and molecular event throughout the body occurs at the right time of the day (Kalsbeek and Buijs, 2002; Holzberg and Albrecht, 2003).
Recent urbanization has greatly changed our light environment by introducing artificial illuminations at night (Hölker et al., 2010). Exposure to irregular LD cycles disrupts the time-keeping functions of the circadian system, which is often associated with detrimental health consequences, including mental health issues (Evans and Davidson, 2013; LeGates et al., 2014). It has been well documented in animal models that aberrant lighting conditions, such as shifted LD cycles, non-24-h LD schedules, constant light, or dim-light exposure at night, result in circadian disturbances as well as anxiety- or depression-like behaviors (Fonken et al., 2009; Fonken et al., 2010; Bedrosian et al., 2011; Fonken et al., 2012; LeGates et al., 2012; Tapia-Osorio et al., 2013; Fonken and Nelson, 2013; Moriya et al., 2015). In addition, human shift workers, who frequently experience nighttime illumination, have also been shown to have a higher risk of developing mood or anxiety disorders (Scott, 2000; Vogel et al., 2012; Morrissette, 2013; Caruso, 2014). Thus, a better understanding of how aberrant lighting conditions affects the circadian system and emotional regulations can provide insights into the mechanisms underlying the health issues associated with shift work.
In the present study, we used a 12:4:4:4-h L:DLD paradigm to explore how chronic light exposure in the middle of the night affects time-keeping functions and emotional responses in mice. Daily patterns of the general locomotor activity and expression of circadian clock components in the SCN and several extra-SCN brain regions (i.e., the paraventricular nucleus [PVN], striatum, hippocampus, and basolateral amygdala [BLA]) were compared between the L:DLD and the control 12:12-h L:D groups. To assess the effects of L:DLD housing on emotional state, anxiety-like behaviors were examined following the analysis of brain-derived neurotrophic factor (Bdnf) messenger RNA (mRNA) expression in brain regions that have shown an association between decreased BDNF and anxiety, including the hippocampus, amygdala, and medial prefrontal cortex (mPFC) (Pandey et al., 2006; Uysal et al., 2011; Sharma et al., 2012; Yu et al., 2015). The results revealed that chronic nighttime light exposure induces a unique pattern of daily rhythm disturbances and negative emotional responses.
Materials and Methods
Animals
Male CD1 mice (28-34 days old) were purchased from Charles River Laboratory (Spencerville, OH) and group-housed (4-5 animals per group) in 18 × 28 × 12-cm Plexiglas cages with Aspen Chip (Nepco, Warrensburg, NY). Room temperature was kept around 22 °C with a humidity level at 50%. The animals were first entrained in 12:12-h light:dark (L:D, 300 lux:1 lux) conditions for 2 weeks before assigned into the experimental and control groups. The experimental group received 4-h light exposure (300 lux) at the middle of the dark phase every night and thus was housed under 12:4:4:4-h L:DLD conditions, while the control group remained in the initial L:D condition that they were entrained into. Food (Prolab 2000 #5P06; PMI Nutrition LLC, St. Louis, MO) and water were available ad libitum throughout the experiment. All procedures used were approved by the Institutional Animal Care and Use Committee of Michigan State University.
General Activity Recording
For recording of general locomotor activity, animals were singly housed in either L:D (control, n = 9) or L:DLD (n = 19) condition, with a motion sensor mounted on top of each cage. General locomotor activities were recorded in 5-minute bins using VitalView (Starr Life Sciences Corp., Oakmont, PA) for at least 2 weeks. Actograms and periodograms were created using ClockLab (Actimetrics, Inc., Wilmette, IL).
Histological Analysis
The mice were group-housed in either L:D (n = 30) or L:DLD condition (n = 30) for 2 weeks. The animals were euthanized with sodium pentobarbital (200 mg/kg; Vortech Pharmaceutical, Ltd., Dearborn, MI) and perfused transcardially using 20 mL saline followed by 50 mL 4% paraformaldehyde in 0.1 M phosphate buffer at 4-h intervals over a daily cycle (n = 5/time point/group), beginning at zeitgeber time 2 (ZT2, the light onset time was defined as ZT0). The brains were postfixed for 12 to 18 h and then cryoprotected overnight in a 20% sucrose solution. Two alternate sets of 40-µm sections were collected from each brain using a cryostat.
Immunocytochemistry
The sections were incubated in the primary antibody against either mouse PER1 or PER2 (raised in rabbit, 1:5000, AB2201, AB2202; Millipore, Billerica, MA) and then incubated in biotinylated goat anti-rabbit IgG secondary antibody (1:200, BA-1000; Vector Laboratories, Burlingame, CA). The sections labeled for PER1 were processed using the avidin-biotin-immunoperoxidase technique with 3,3′-diaminobenzidine tetrahydrochloride as the chromogen (DAB; Sigma-Aldrich, St. Louis, MO). Sections incubated in a PER2 antibody were processed with avidin-biotin-immunoperoxidase technique using DAB enhanced with 4% nickel sulfate as the chromogen. To deal with potential variability in independent immunohistochemistry (ICC) runs, animals that were compared directly were processed together with unique cut to mark each brain. Following the ICC reaction, the sections were mounted on slides, rinsed with ethanol and xylene, and coverslipped with Permount (Fisher Scientific, Fair Lawn, NJ).
ICC data analysis
Images of sections were captured using a light microscope (Zeiss, Gottingen, Germany) and a CCD video camera (CX9000; MBF Bioscience, Williston, VT). The PER1- or PER2-immunoreactive (ir) nuclei were counted in the SCN, PVN, striatum, hippocampus, and BLA using National Institutes of Health (NIH) Image J program as described previously (Yan and Silver, 2004; Shuboni and Yan, 2010; Yan, 2011). For the SCN, PER1- or PER2-ir nuclei were counted bilaterally in 3 mid-SCN sections. The boundary between the ventrolateral “core” and dorsomedial “shell” subregions was determined based on the template from a previous study (Yan and Silver, 2004). For the PVN, PER1- or PER2-ir nuclei were counted bilaterally in 3 sections containing magnocellular and parvocellular nuclei of the PVN and averaged. For the dorsal striatum and BLA, 2 or 3 sections were used and PER1- or PER2-ir nuclei were counted within the area, which was set consistently across the sections in the right hemisphere and calculated as cell counts per 104 µm2. For the hippocampus, PER1- and PER2-ir nuclei were counted in different subfields, the CA1 and CA3 areas, of the right hemisphere in 3 sections of the dorsal rostral hippocampus. Immunoreactive nuclei were counted along the pyramidal cell layer and calculated as counts per 104 µm2.
Open Field Test
After 2 weeks of group housing in the L:DLD or L:D condition, animals (n = 8/group) were subjected to the open field test at ZT5 to 7 using a box at 45 × 45 × 60 cm in a testing room with light intensity ~30 lux. The arena was divided to 3 × 3 grids, and the central square (15 × 15 cm) was defined as the center. Animals were place individually in the center of testing arena and allowed to explore freely for 5 min. Trials were recorded and manually scored for the amount of time spent in the center by observers who were unaware of the experimental condition of the animals. After the 5-min trial, the number of fecal boli on the arena was counted. The area was wiped with a 70% ethanol solution between the tests for each animal. The testing was conducted in 2 arenas simultaneously, and no testing order effect was observed.
Tissue Collection, mRNA Isolation, and Real-Time Polymerase Chain Reaction
Two weeks after the open field test, brains were collected from the animals and Bdnf mRNA expression was examined in the hippocampus, amygdala, and mPFC. Animals (n = 8/group) were euthanized with sodium pentobarbital (200 mg/kg) and rapidly decapitated at ZT6. Brains were removed and placed in RNAlater (Ambion, Austin, TX). The hippocampus and mPFC were dissected from the brain according to Spijker (2011). Briefly, the entire hippocampus was gently separated from the cortex and removed. Next, coronal slices (at 1 mm) were made, and mPFC was dissected using a razor blade. The amygdala containing the BLA and central nucleus of amygdala was also dissected based on the brain atlas in Paxinos and Watson (2007). Total RNA was isolated using Trizol (Life Technologies, Carlsbad, CA), and RNA concentrations and purity were checked using a GeneQuant 100 Spectrophotomerter (GE Healthcare, Little Chalfont, UK). DNA in RNA samples was digested with deoxyribonuclease I, Amplification Grade (Life Technologies). Complementary DNAs (cDNAs) were synthesized from 2 µg of total RNA using a High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). For real-time polymerase chain reaction (PCR) analysis, 1% of the cDNA was used at a final concentration of 1× Power SYBR Green PCR Master Mix (Life Technologies) and 0.05 µM of each primer using an ABI 7000 Real-Time PCR System (Life Technologies). Each reaction was performed in duplicates. Forward and reverse primer sequences were 5′-AGA AAG CTG CTT CAG GAA ACG C-3′ and 5′-TTA CGC AAA CGC CCT CAT TCT G-3′, respectively, for Bdnf, and 5′-ATC CAC TGG TGC TGC CAA G-3′ and 5′-CCG TTC AGC TCT GGG ATG AC-3′, respectively, for Gapdh, which was used as a reference gene. In all the reactions, the generation of only a single expected amplicon was confirmed by melting analysis. Quantification of cDNAs was performed by the standard curve methodology.
Statistical Analysis
The statistical analyses were conducted using the program, R (R CoreTeam, 2013). The general activity was analyzed by the 2-way repeated-measure analysis of variance (ANOVA; time × lighting conditions). The Student t test was used to further evaluate group differences at each ZT. Expression of PER1 and PER2 was examined by 1-way ANOVA or 2-way ANOVA (time × lighting conditions), and the Student t test was used to further evaluate group differences at each ZT. Dunnett’s test was used to compare PER1 and PER2 expression at each ZT to those at ZT with the lowest expression (i.e., ZT2 for the SCN and ZT14 for the extra-SCN regions). The results of the open field test and Bdnf mRNA expression were analyzed using the Student t test. Mean differences were considered statistically significant when p < 0.05.
Results
Chronic Nighttime Light Exposure Altered the General Activity Pattern
Animals in the control L:D condition showed robust nocturnal rhythms in general locomotor activity (Fig. 1A). In the L:DLD condition, although the majority of the activity occurred during the night phase, the behavioral bouts were fragmented (Fig. 1B). Two of 19 L:DLD animals became arrhythmic at the third week in the L:DLD condition (data not shown). The averaged activity for the first 2 weeks in the L:D or L:DLD condition is shown in Figure 1C. In both conditions, animals increased their activity around dark onset (ZT12) and light onset (ZT0). However, while an increase of the activity in L:D animals preceded the lights-off by 30 min (ZT11.5), the activity of L:DLD animals reached the peak 30 min after dark onset (ZT12.5) (effect of lighting conditions, F1,26 = 2.66, p = 0.12; effect of time, F12,312 = 19.94, p < 0.01; interaction, F12,312 = 7.00, p < 0.01) (Fig. 1D). Similarly, while the activity of L:D animals reached the peak at ZT22 and decreased before light onset, L:DLD animals increased their activity at light onset (ZT0) (effect of conditions, F1,26 = 5.82, p = 0.023; effect of time, F12,312 = 4.02, p < 0.01; interaction, F12,312 = 2.59, p < 0.01) (Fig. 1E). There was no difference in the total activity within 24 h between L:D and L:DLD condition (t26 = 0.11, p = 0.92) (Fig. 1F).
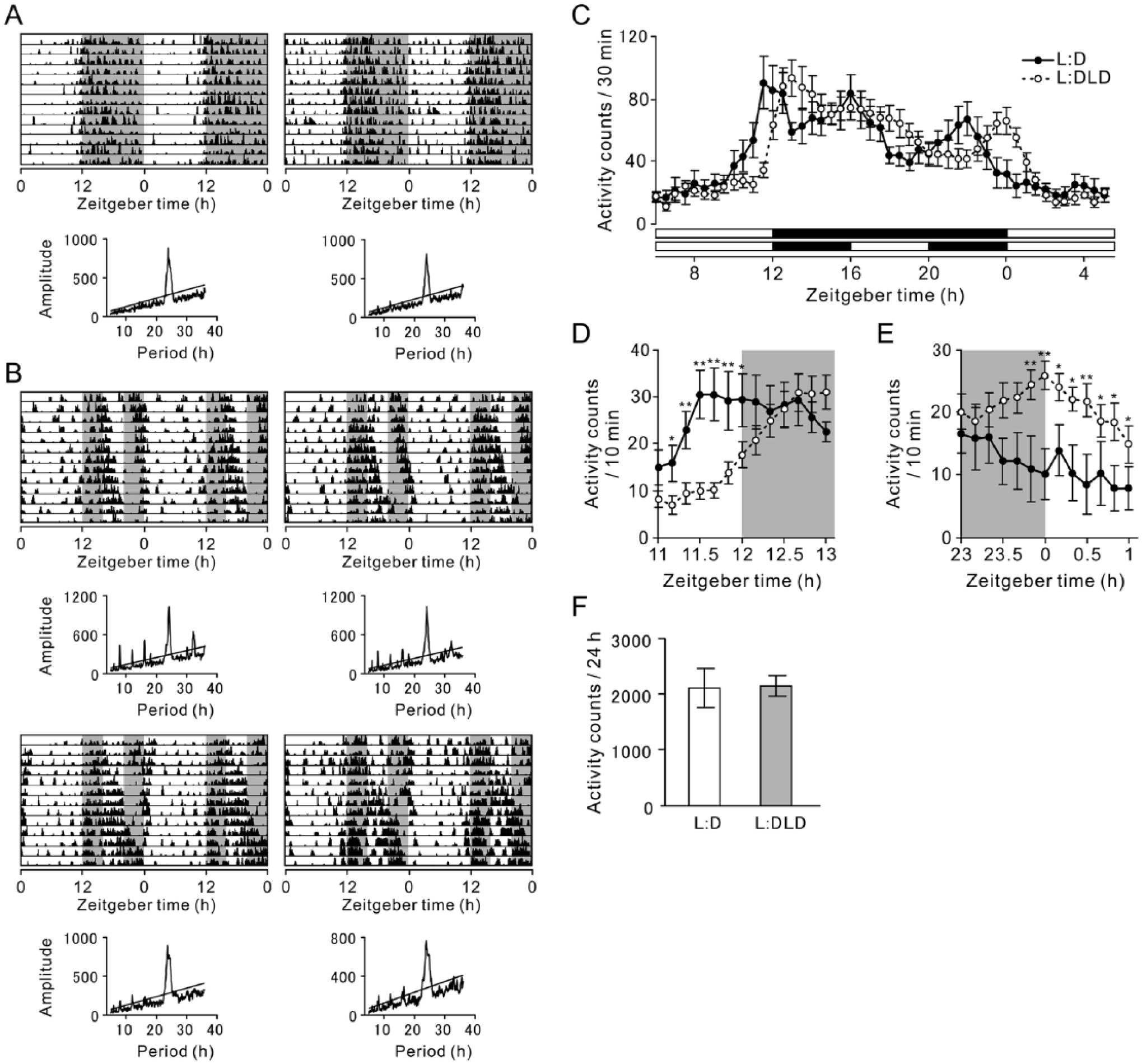
General activity rhythms in L:D and L:DLD conditions. Representative double-plotted actograms and corresponding periodograms of mice in (A) L:D and (B) L:DLD conditions are shown. (C) Activity counts for 30 min were averaged (L:D, n = 9; L:DLD, n = 19). Mean activity counts for 10 min around (D) light offset and (E) light onset. (F) Averaged activity counts for 24 h. *p < 0.05, **p < 0.01.
Chronic Nighttime Light Exposure Abolished PER1 and PER2 Rhythms in the SCN PER1 Expression
Clear daily variations in PER1 expression in the SCN were observed in the L:D but not in the L:DLD condition (Fig. 2A). The number of PER1-ir nuclei was analyzed in the entire SCN and within the core and shell subregions (Fig. 2C). Statistical analysis revealed that the daily rhythms in the L:DLD group were abolished in the SCN and within each subregion (1-way ANOVA, effect of time, p > 0.05). A similar pattern was observed for PER2 expression (Fig. 2B) and confirmed by quantitative analysis (Fig. 2D). Detailed results of statistical analyses are shown in Table 1.
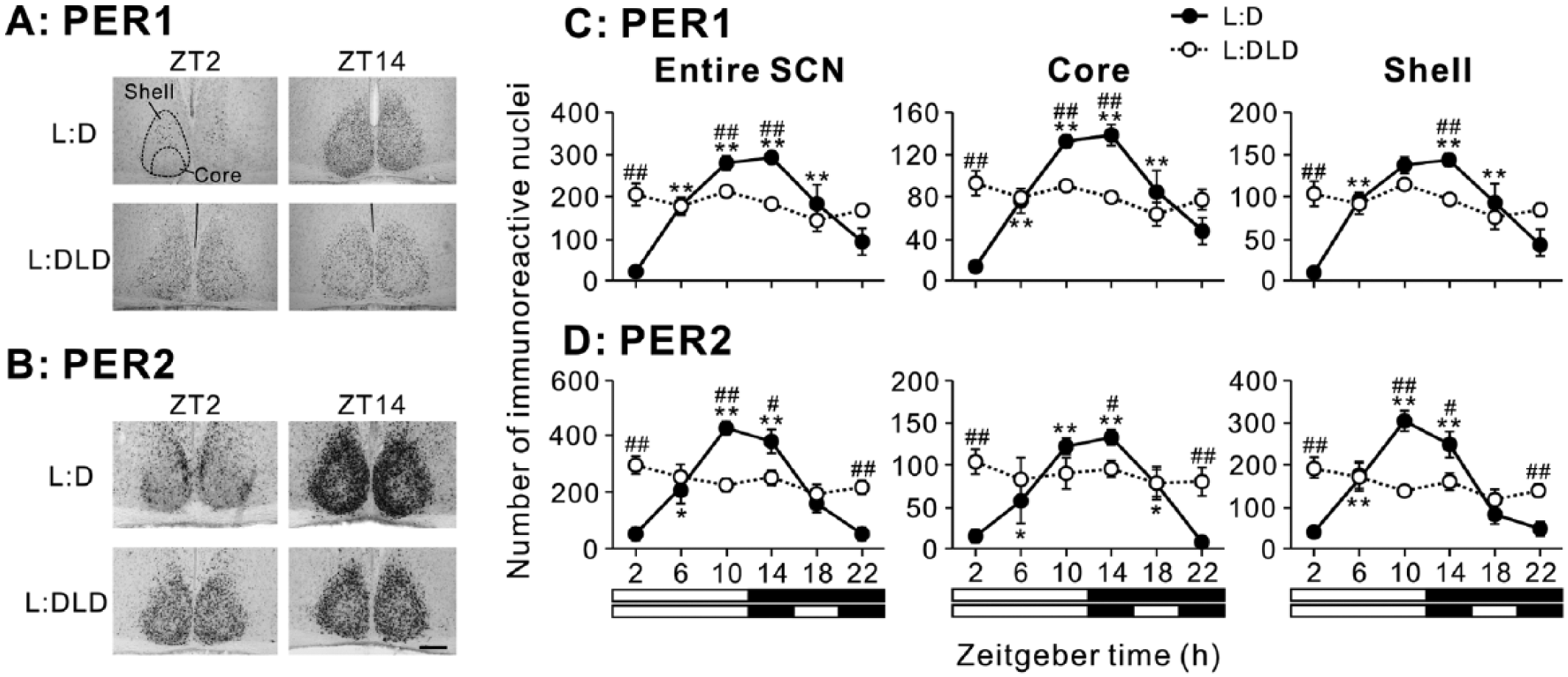
PER1 and PER2 expression in the SCN of L:D and L:DLD mice. Representative photographs depicting (A) PER1 and (B) PER2 immunoreactivity in the SCN. Scale bar: 100 µm. Quantitative analyses showing the number of (C) PER1-ir and (D) PER2-ir nuclei in the entire SCN (left), core (middle), and shell (right) regions. *p < 0.05, **p < 0.01 (vs. ZT2 in the same group); #p < 0.05, ##p < 0.01 (L:D vs. L:DLD at the same time point).
Statistical Results.
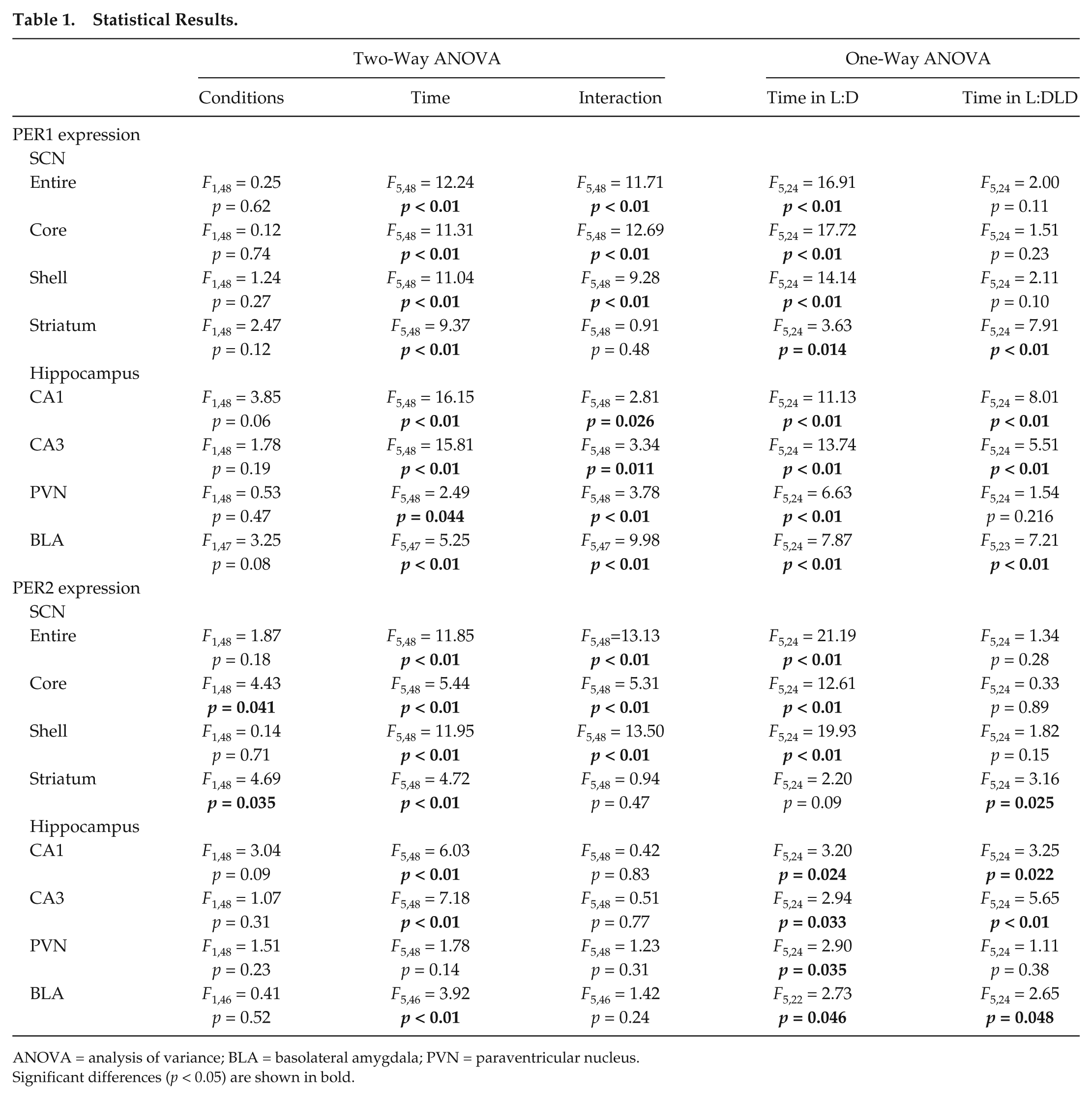
ANOVA = analysis of variance; BLA = basolateral amygdala; PVN = paraventricular nucleus.
Significant differences (p < 0.05) are shown in bold.
PER1 and PER2 Rhythms Remained Relatively Intact in the Striatum and Hippocampus
In the dorsal striatum (Fig. 3A), PER1 expression remained rhythmic in the L:DLD condition, without significant difference from the L:D group (2-way ANOVA: effect of time, p < 0.01; effect of conditions, p > 0.05; interaction of time and conditions, p > 0.05; Fig. 3B,D). The expression of PER2 also remained rhythmic in the striatum in the L:DLD group (Fig. 3C,E). In addition to a significant effect of time (p < 0.01), 2-way ANOVA revealed a significant effect of lighting conditions (p < 0.05), although there was no significant interaction between the 2 factors (p > 0.05). Follow-up 1-way ANOVA suggests that the daily rhythm in PER2 expression was slightly enhanced in the L:DLD condition (effect of time: p = 0.09 in L:D, p < 0.05 in L:DLD conditions). Detailed results of statistical analyses are shown in Table 1.
PER1 and PER2 expression in the dorsal striatum of L:D and L:DLD mice. (A) The dorsal striatum used for the analysis. Scale bar: 500 µm. Representative photographs depicting (B) PER1 and (C) PER2 immunoreactivity. Scale bar: 100 µm. Quantitative analyses showing the number of (D) PER1-ir and (E) PER2-ir nuclei. *p < 0.05, **p < 0.01 (vs. ZT14 in the same group).
In the hippocampus, PER1 and PER2 expression showed daily rhythms in CA1 and CA3 areas in both L:D and L:DLD groups (Fig. 4A-C). In both CA1 and CA3 areas, 2-way ANOVA revealed a significant effect of time (p < 0.01) but no significant effect of lighting conditions (p > 0.05) for both PER1 and PER2-ir (Fig. 4D,E). A significant interaction between the lighting conditions and time was also detected for PER1 (p < 0.05), likely due to the elevated expression in the L:DLD condition at ZT2 (t8 = 2.97, p < 0.05). Detailed results of statistical analyses are shown in Table 1.
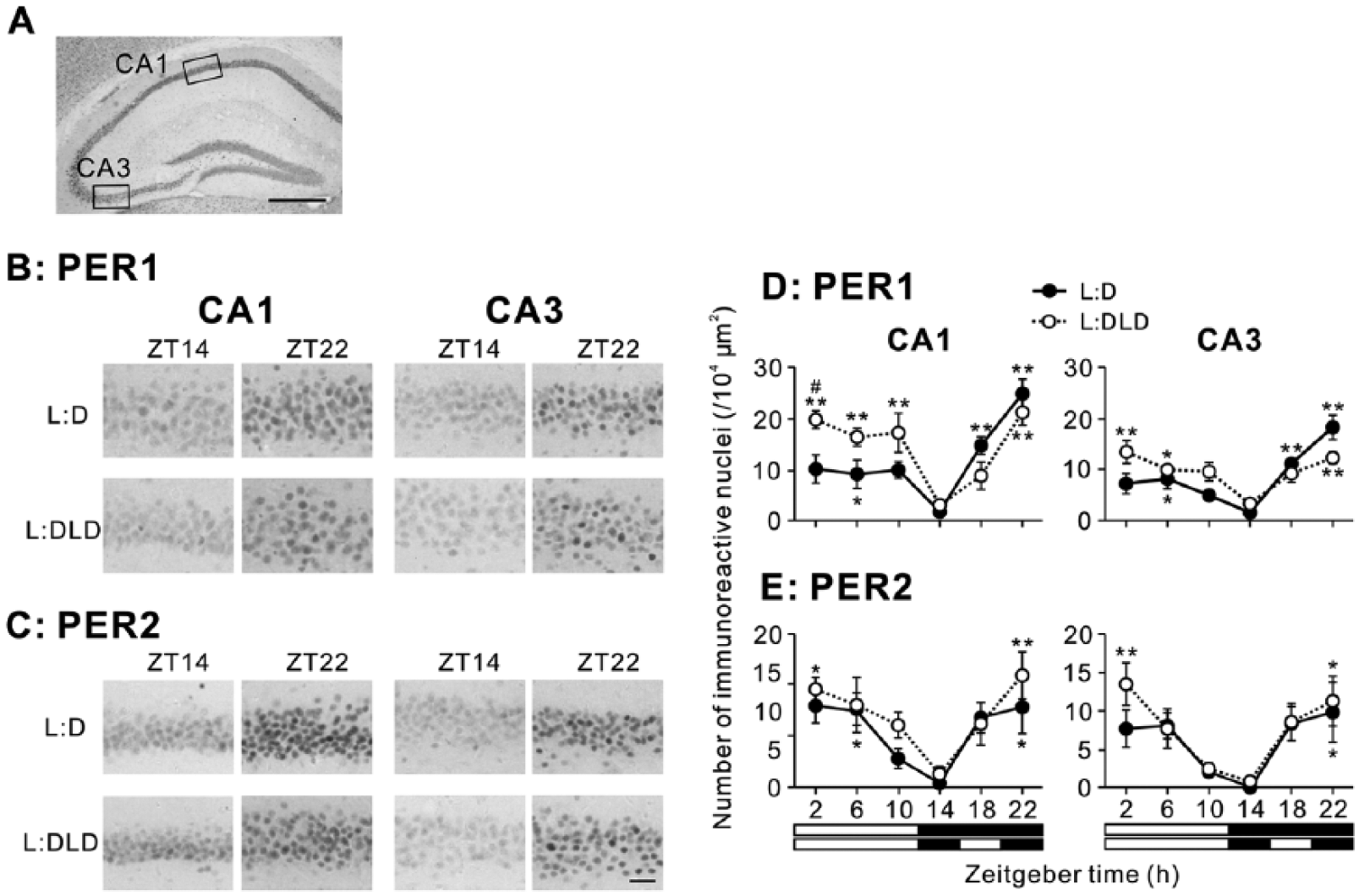
PER1 and PER2 expression in the hippocampus of L:D and L:DLD mice. (A) Hippocampal subfields used for the analysis. Scale bar: 500 µm. Representative photographs depicting (B) PER1 and (C) PER2 immunoreactivity in the CA1 and CA3 regions. Scale bar: 50 µm. Quantitative analyses showing the number of (D) PER1-ir and (E) PER2-ir nuclei in CA1 (left) and CA3 (right) regions. *p < 0.05, **p < 0.01 (vs. ZT14 in the same group); #p < 0.05 (L:D vs. L:DLD at the same time point).
PER1 and PER2 Rhythms Were Altered in the PVN and BLA
In the PVN, higher PER1 expression was observed at night in the L:D condition but during the day in the L:DLD condition (Fig. 5A,C). Two-way ANOVA revealed a significant interaction between lighting conditions and time (p < 0.01). At ZT2, PER1 expression in the L:DLD condition was significantly higher than that in the L:D condition (t8 = 3.35, p < 0.05) (Fig. 5C). The pattern of PER2 expression was similar to that of PER1 (Fig. 5B,D). However, 2-way ANOVA failed to reveal any significant effects of time and conditions or interaction between the factors (p > 0.05). One-way ANOVA for each lighting condition revealed a significant effect of time in the L:D (p < 0.05) but not in the L:DLD condition (p > 0.05). Detailed results of statistical analyses are shown in Table 1.
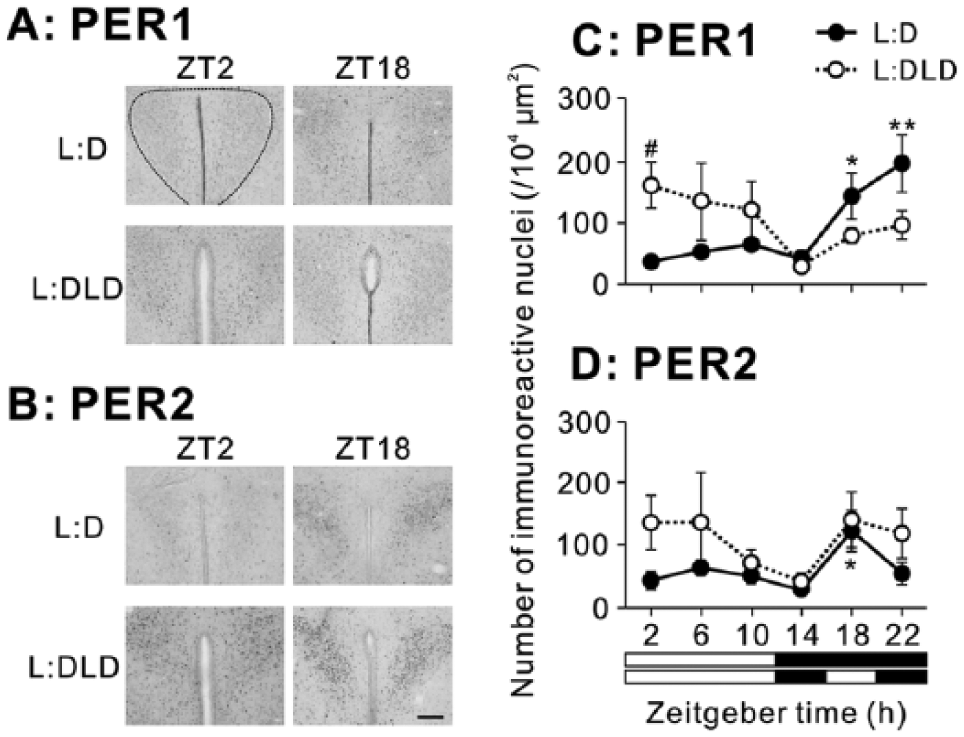
PER1 and PER2 expression in the PVN of L:D and L:DLD mice. Representative photographs depicting (A) PER1 and (B) PER2 immunoreactivity. Scale bar: 100 µm. Quantitative analyses showing the number of (C) PER1-ir and (D) PER2-ir nuclei. *p < 0.05, **p < 0.01 (vs. ZT14 in the same group); #p < 0.05 (L:D vs. L:DLD at the same time point).
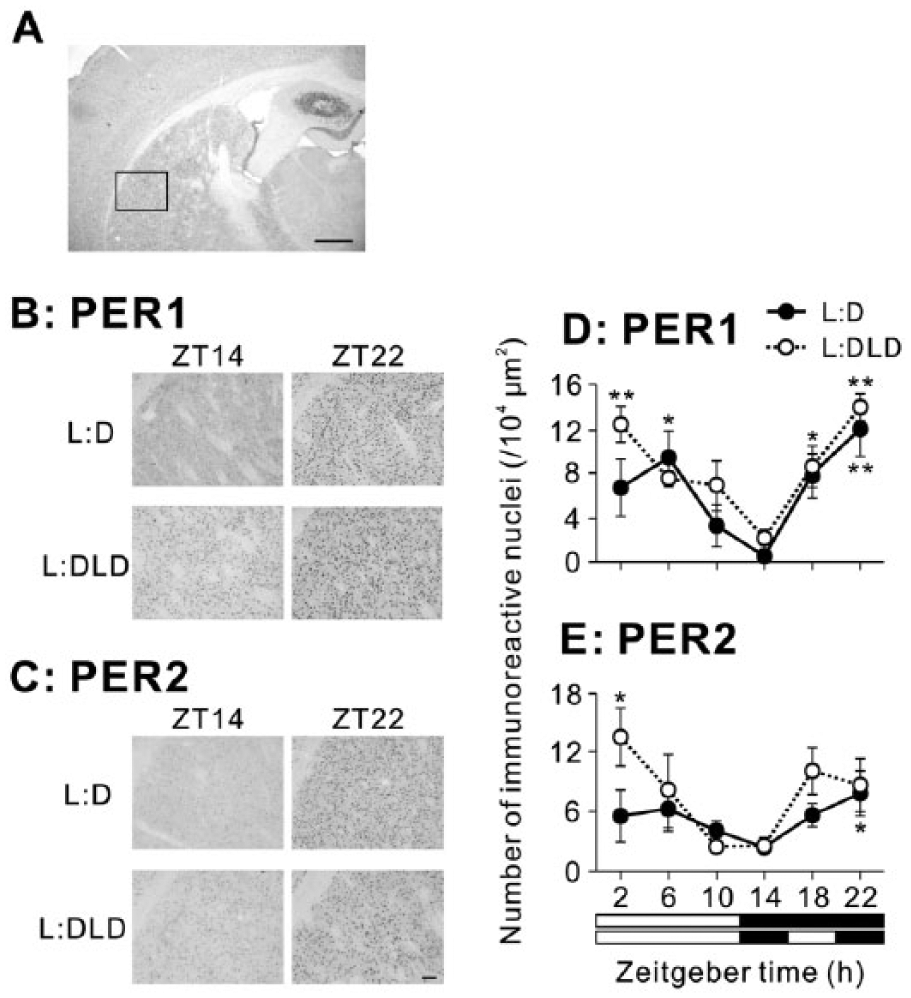
In the BLA, PER1 expression increased at night in the L:D condition but increased during the day in the L:DLD condition (Fig. 6A). Two-way ANOVA revealed a significant interaction between conditions and time (p < 0.01; Fig. 6C). PER1 expression was elevated in the L:DLD conditions compared to the L:D condition during the day (t8 = 4.04, p < 0.01 for ZT2; t8 = 11.21, p < 0.01 for ZT10), but it was decreased in the L:DLD condition compared to the L:D condition at ZT18 (t8 = 3.04, p < 0.05) (Fig. 6C). There were no differences in PER2 rhythms in the BLA between L:D and L:DLD conditions (2-way ANOVA: effect of time, p < 0.01; effect of conditions, p > 0.05; interaction between the factors, p > 0.05; Fig. 6B,D). However, PER2 expression in the L:DLD condition was significantly higher than that in the L:D condition at ZT2 (t7 = 2.73, p < 0.05; Fig. 6D). Detailed results of statistical analyses are shown in Table 1.
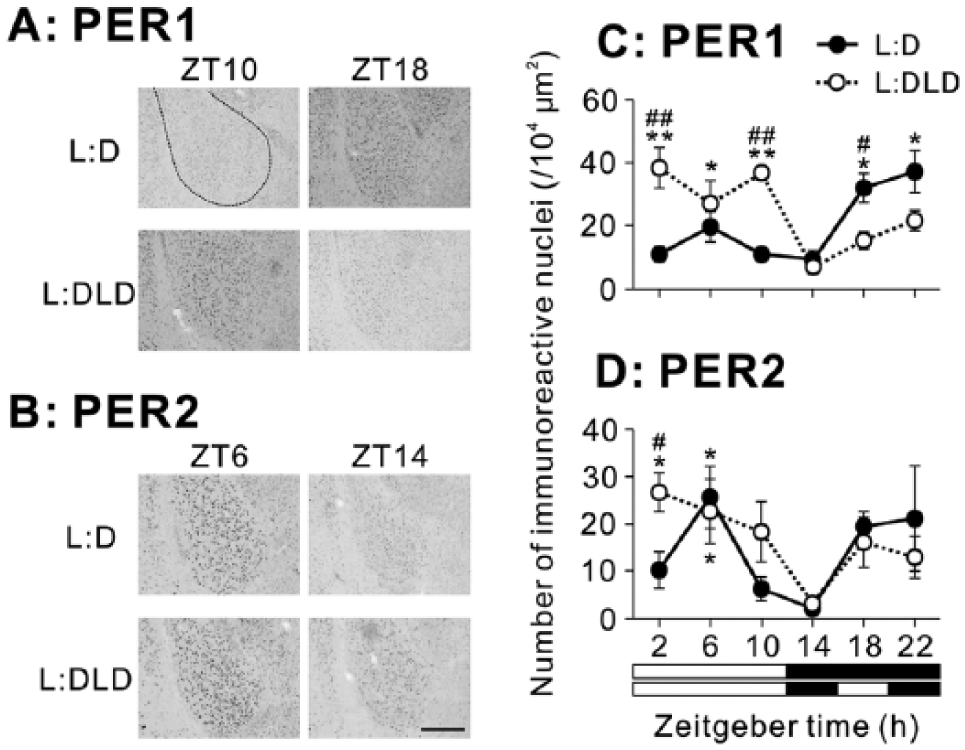
PER1 and PER2 expression in the BLA of L:D and L:DLD mice. Representative photographs depicting (A) PER1 and (B) PER2 immunoreactivity. Scale bar: 100 µm. Quantitative analyses showing the number of (C) PER1-ir and (D) PER2-ir nuclei. *p < 0.05, **p < 0.01 (vs. ZT14 in the same group); #p < 0.05, ##p < 0.05 (L:D vs. L:DLD at the same time point).
Chronic Nighttime Light Exposure Led to Increased Anxiety-Like Behaviors and Decreased Bdnf mRNA Expression
During the open field test, most of the animals moved away from the center right after being placed and started exploring the arena. The L:DLD group spent significantly less time in the center than the L:D group (t14 = 2.78, p = 0.02; Fig. 7A). In addition, the L:DLD group produced more fecal boli compared to the L:D group during the test (t14 = 2.44, p = 0.03; Fig. 7B).
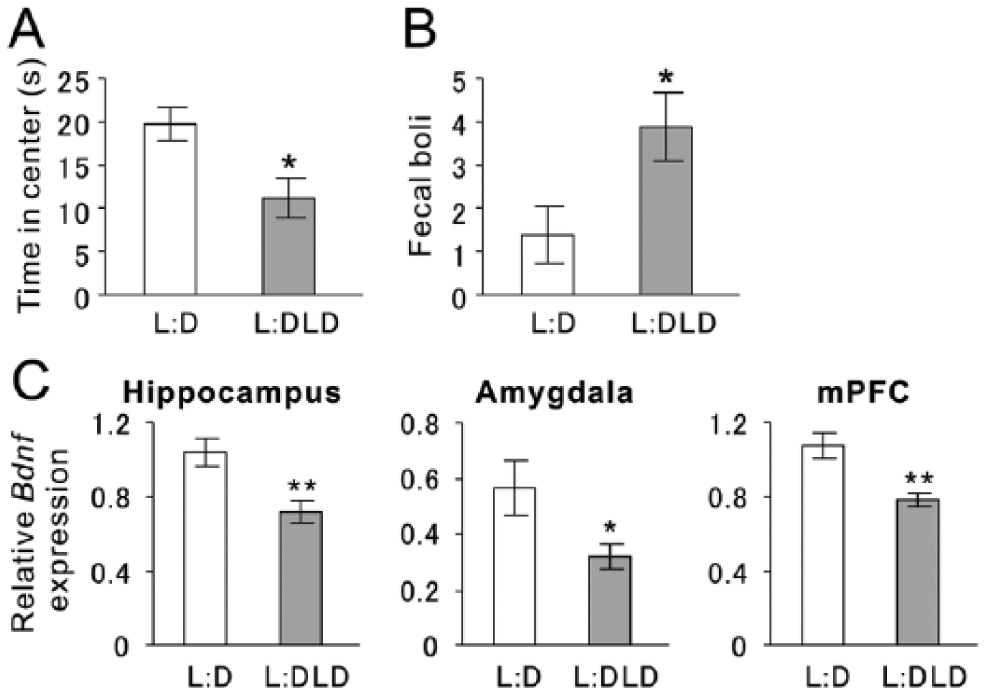
Anxiety-like behaviors and Bdnf messenger RNA (mRNA) expression of L:D and L:DLD mice. (A) Total time spent in center of the arena during the open field test. (B) Total numbers of fecal boli during the open field test. (C) Relative Bdnf mRNA expression was examined in the hippocampus, amygdala, and medial prefrontal cortex (mPFC). *p < 0.05, **p < 0.01.
Compared to the L:D group, the L:DLD group also showed lower levels of Bdnf mRNA expression in the hippocampus (t14 = 3.29, p < 0.01), amygdala (t14 = 2.34, p = 0.035), and mPFC (t13 = 3.42, p < 0.01) (Fig. 7C).
Discussion
In the present study, a 4-h light interval was introduced to the 12-h dark phase that resulted in a 12:4:4:4-h L:DLD. This paradigm revealed a unique disturbance of the circadian system with the rhythms of the SCN and extra-SCN oscillators responding differently in a region-specific fashion. The L:DLD paradigm resulted in altered daily patterns in locomotor activity and the absence of daily rhythms of PER1 and PER2 expression in the SCN. However, the daily rhythms of PER1 and PER2 expression remained intact in some of the extra-SCN regions (i.e., the hippocampus and striatum) but more disturbed in other regions, including the PVN and BLA. The results suggest that the L:DLD paradigm leads to desynchrony among brain oscillators and disturbs the circadian time-keeping system. Additionally, mice exposed to L:DLD showed increased anxiety-like behaviors as well as decreased Bdnf mRNA expression in 3 brain regions important for emotional regulations.
Compared to the control L:D condition, mice in the L:DLD condition showed fragmented activity patterns. In rodents (i.e., Syrian hamsters, Siberian hamsters, or mice), the 24-h LDLD cycle can induce “splitting” or “bifurcation” in activity rhythms, with 2 activity bouts entrained to each of the twice-daily scotophases (Gorman and Lee, 2001; Gorman and Elliott, 2003; Evans et al., 2005; Watanabe et al., 2007; Evans et al., 2011). However, the L:DLD paradigm used in the present study did not result in a clear bifurcation with 2 well-entrained components in the locomotor activity. The different behavioral outcome could be due to species differences, nonphotic cues (i.e., running wheels or cage changing), or the difference in the actual lighting schedules (i.e., 12:4:4:4 vs. 7:5:7:5 h). Nonetheless, presently the fragmentation in the daily activity bouts was evident in actograms and periodograms, suggesting a disturbance in the daily rhythms of the L:DLD animals (Fig. 1B). Although the animals showed a masking response to light (i.e., decreased activity when first introduced to the L:DLD schedule), they resumed their activity during the 4-h light phase after a few days (Fig. 1B). When the activity was averaged over 2 weeks, there was no indication of the suppressed activity level in the L:DLD animals during the 4-h light phase (Fig. 1C), suggesting that the fragmentation in their activity pattern was unlikely due to negative masking. The disturbed clock function is also indicated by the altered phase angle in L:DLD compared to control L:D mice. Control L:D mice showed increased activity about 30 min before lights-off and decreased activity 2 h before lights-on, anticipating the light/dark transitions (i.e., dusk and dawn). This phenomenon has been demonstrated in a variety of animals and requires a functional circadian clock (Wheeler et al., 1993; Majercak et al., 1999; Dudley et al., 2003; Mathew et al., 2013; Loos et al., 2014). In contrast, the activity of L:DLD mice reached peak levels about 1 h after the dark onset without any anticipatory activity prior to dusk. Thus, these behavioral data (i.e., fragmented nighttime activity and altered anticipation of light/dark transition) collectively suggest the clock function in the L:DLD group was disturbed.
In accordance with the findings from the behavioral study, we found that the daily rhythms of PER1 and PER2 were abolished in the SCN of L:DLD mice. The results need to be interpreted cautiously as it is possible that the SCN in individual L:DLD mice was still rhythmic but out of phase with each other, which could also result in a loss of rhythms at the group level. This is unlikely, though, because PER1 and PER2 expression in the SCN of all L:DLD mice showed moderate levels consistently across each time point and the entire group with variations among animals comparable to the L:D group. The intact rhythms in the hippocampus and striatum also suggest that the abolished rhythms observed in the SCN were not due to the different phase of free-running of each animal but rather a loss of synchrony among cellular oscillators within the SCN. Previous studies have shown that in mice or hamsters showing bifurcated behavioral rhythms under L:DLD conditions, the cellular oscillators within the SCN reorganized into 2 groups, with antiphased oscillation in the anatomically distinct ventral/core and dorsal/shell subregions (Yan et al., 2010). We thus further analyzed the daily rhythms of PER1 and PER2 in each subregion but did not find any time-dependent changes in the expression of either protein. Due to technical limitations, we are unable to analyze the daily oscillation within individual cells or in a tightly coupled group done in recent work using real-time imaging (An et al., 2013; Koinuma et al., 2013; Evans et al., 2013). However, given that there were rhythmic components in their activity, it is likely that at least a subset of SCN neurons remained synchronized with each other in the L:DLD condition and produced daily oscillation that responsible for the behavioral rhythms. We expect when the perturbation is removed (e.g., if the animals are released to constant darkness or the L:D condition), the SCN neurons will resynchronize with each other and start to produce overt rhythms.
The daily rhythms of PER1 and PER2 were also examined in extra-SCN brain regions. In the control L:D condition, the temporal patterns of the PER1 and PER2 expression in the striatum, hippocampus, PVN, and BLA were consistent with the previous studies (Lamont et al., 2005; Feillet et al., 2008; Girotti et al., 2009; Harbour et al., 2013). Although PER1 immunoreactivity was not previously found in the PVN and BLA of mice (Feillet et al., 2008), the present study revealed rhythmic PER1 expression in the PVN and BLA with a peak level at the end of night in the L:D condition, which is consistent with the previous demonstrations that Per1 mRNA was rhythmically expressed in these brain regions and increased at the early night (Takahashi et al., 2001; Savalli et al., 2015b). In the L:DLD condition, contrary to the severe disruption on the SCN, the temporal patterns of PER1 and PER2 in the striatum and hippocampus remained intact and showed relatively greater amplitude. On the other hand, the rhythms of PER1 and PER2 were abolished in the PVN and phase-shifted in the BLA. The results suggest that the extra-SCN brain regions responded differently to the L:DLD paradigm, leading to misalignment among multiple brain oscillators in addition to disruption of the SCN clock.
Evidence to date suggests that the molecular oscillations in extra-SCN regions are driven by rhythmic outputs from the SCN (Kalsbeek and Buijs, 2002; Holzberg and Albrecht, 2003). However, our findings that nighttime light exposure disturbed SCN and PVN oscillations but not other extra-SCN oscillations (i.e., the striatum, hippocampus, and BLA) suggest that these brain regions can maintain daily rhythms independent from the overt rhythms within the SCN. As discussed previously under the behavioral data, it is possible that a distinct subset of SCN cells that remained synchronized with each other were capable to entrain the rhythms in these extra-SCN sites in the L:DLD group. Alternatively, non-SCN factors (e.g., rhythmic feeding or locomotor activity) could have also contributed to the rhythmic patterns in some of the extra-SCN sites. The responses of extra-SCN oscillators to feeding schedules vary among the brain regions or between PER1 and PER2 (Ángeles-Castellanos et al., 2007; Verwey et al., 2007; Feillet et al., 2008), which could have resulted in different responses in PER1 and PER2 rhythms in different brain regions observed in the present study. It should be acknowledged that the present study only analyzed 2 clock proteins. Future studies examining the expression of other clock genes in addition to PER1 and PER2, especially combined with the cutting-edge imaging techniques that allow single-cell analysis, will provide a more complete picture on how an aberrant lighting condition affects the circadian clock function.
The circadian system has been implicated in emotional processes (Evans and Davidson, 2013; LeGates et al., 2014), which is supported by studies using animal models with deficiency in clock genes (Kondratova et al., 2010; De Bundel et al., 2013; Savalli et al., 2015a) or using aberrant lighting conditions (Fonken et al., 2009; Fonken et al., 2010; Bedrosian et al., 2011; Fonken et al., 2012; LeGates et al., 2012; Tapia-Osorio et al., 2013; Fonken and Nelson, 2013; Moriya et al., 2015). Especially, the importance of clock gene oscillations in the BLA has been demonstrated in mice showing depression-related behaviors (Savalli et al., 2015a). In the present study, we explored the effect of L:DLD housing on emotional regulations. In the open field test, L:DLD mice spent less time at the center of the open arena and also produced more fecal boli compared to the L:D controls, indicating an increase in anxiety-like responses in their behavior and physiology (Simon et al., 1994; Ennaceur et al., 2006). Future studies using a battery of other tests for anxiety-like behaviors (e.g., the light/dark box or elevated plus maze) will provide further support of the behavioral phenotype observed in the present study.
We also found reduced Bdnf mRNA expression in the hippocampus, amygdala, and mPFC. BDNF plays critical roles in neuronal plasticity, and deficits in BDNF signaling in various brain regions are implicated in the neuropathology of affective disorders (Yu and Chen, 2011). The hippocampus, amygdala, and mPFC have a reciprocal network for emotional controls (LaBar and Cabeza, 2006; Godsil et al., 2013). Decreased BDNF in these regions has been associated with anxiety in animal models (Pandey et al., 2006; Uysal et al., 2011; Sharma et al., 2012; Yu et al., 2015) and in human patients (Dwivedi et al., 2003; Dwivedi et al., 2005; Karege et al., 2005). The expression of Bdnf mRNA and BDNF protein undergoes diurnal variations (Bova et al., 1998; Liang et al., 1998; Berchtold et al., 1999; Schaaf et al., 2000; Katoh-Semba et al., 2008; Hamatake et al., 2011; Golini et al., 2012; Ikeno et al., 2013; Martin-Fairey and Nunez, 2014). It is possible that the L:DLD paradigm alters the circadian regulation of Bdnf expression, resulting in diminished Bdnf expression. Indeed, decreased Bdnf expression in the amygdala has been shown in animals following a phase shift (Katoh-Semba et al., 2008) and in clock gene mutants (Savalli et al., 2015a). Taken together, our findings of increased anxiety-like responses and decreased Bdnf expression in L:DLD mice suggest that chronic nighttime light exposure alters not only the circadian system but also the neural pathways underlying emotional regulation.
In summary, the present study demonstrates that light exposure at night induces circadian disturbances in a brain region–specific manner: some regions including the SCN were severely disturbed, while others including the hippocampus were less affected. In addition, the L:DLD paradigm results in emotional dysregulations. Exposure to light at nighttime occurs to many people due to night-shift, sleep disorder, or lifestyle choice. Further research of the underlying mechanisms through which nighttime light exposure differently modulates brain oscillators should contribute to a better understanding of the health issues associated with circadian rhythm disruption.
Footnotes
Acknowledgements
The authors thank Joel Soler, Sean Deats, and Widya Adidharma for technical assistance and Dr. Tony Nunez for helpful comments on the manuscript. This work is supported by a MSU IRGP grant to LY.