
Abstract
Circadian rhythms in mammals are regulated by the hypothalamic suprachiasmatic nucleus (SCN). The generation of circadian oscillations is a cell-autonomous property, and coupling among cells is essential for the SCN to function as a pacemaker. The development of SCN anatomy and cytology has been extensively studied, but the point in development when the SCN first has the capacity to generate circadian oscillations has not been established. We therefore examined the development of circadian oscillations using per2::luc mice in which bioluminescence tracks the expression of the circadian clock protein, PER2. In vitro, hypothalamic explants first expressed consistent oscillations when isolated between 15 and 16 days postfertilization (e15). Oscillations were more robust at later ages. Explants from other brain areas did not express oscillations, indicating that the development of oscillations is not a general property of embryonic tissue. SCN explants obtained on e14 did not initially express oscillations but developed them in vitro over 4 to 6 d. Although coupling among cells is required for the long-term expression of tissue-level oscillations, explants from mice lacking the coupling peptide vasoactive intestinal peptide still developed oscillations. In the mouse, the capacity to generate molecular oscillations on e15 coincides with the completion of neurogenesis and SCN-specific transcription factor expression. Thus, within a day of its genesis at an age approximately equivalent to the end of the first trimester in humans, the SCN develops the capacity to express circadian oscillations and autonomously develops mechanisms sufficient to couple and synchronize its cells.
Keywords
Circadian cycles are regulatory components of nearly all cells, from bacteria to differentiated mammalian cells. They arise from cell-autonomous molecular and biochemical processes, including interactions among several transcriptional activators and repressors (circadian regulatory genes [CRGs]) whose expression is essential for the generation and persistence of the cycles (Ko and Takahashi, 2006). The rhythmic activity of CRGs permeates regulatory networks throughout the cell, ultimately driving rhythmicity in a wide range of cellular processes from metabolic pathways to signal transduction (Takahashi et al., 2008). In multicellular organisms, the cells within a tissue are sometimes synchronized directly by entrainment to environmental cues, particularly light (Whitmore et al., 2000). In mammals, however, cells and tissues are entrained to rhythms in physiology such as neural or endocrine signals or rhythms in temperature and feeding. These in turn are regulated by a central nervous system pacemaker, the hypothalamic suprachiasmatic nucleus (SCN; Dibner et al., 2010). The SCN and the rhythms it regulates are entrained by the environmental light/dark cycle via direct retinal projections. The SCN, its input and output mechanisms, and the peripheral tissue rhythms it regulates constitute a circadian system presumed to generate, under normal conditions, an adaptive, functionally significant temporal structure, from the timing of sleep relative to environmental cycles to the management of energy intake, storage, release, and use (Dijk and Czeisler, 1994; Rudic et al., 2004; Lamia et al., 2008; Martino et al., 2008; Takahashi et al., 2008; Vollmers et al., 2009; Dibner et al., 2010).
At the cellular level, circadian cycles transcriptionally and posttranscriptionally regulate the expression of thousands of genes (Koike et al., 2012). While the scale of circadian regulation is similar for all cells, the complement of genes regulated by circadian cycles differs depending on the differentiated cell type (Panda et al., 2002; Storch et al., 2002; McCarthy et al., 2007). This suggests that although the core CRGs are the same in all cells, the targets of circadian regulation are determined as part of cellular differentiation. In embryonic stem cells, circadian cycles appear to be repressed by factors that promote and maintain pluripotency, indicating that circadian organization, including the initiation of core regulatory cycles and their control of downstream targets, also occurs as part of cellular differentiation (Kowalska et al., 2010; Yagita et al., 2010). Although when and how circadian regulatory cycles develop in mammalian tissues are not known, it is reasonable to expect that this occurs in peripheral tissues after differentiation of the SCN and the development of the output mechanisms required to coordinate the circadian system. The expression of circadian cycles in individual cells of peripheral tissues might not be functionally useful without synchronization and regulation of the circadian system as a whole by the SCN (Yoo et al., 2004; Reddy et al., 2007; Loh et al., 2011).
Current evidence indicates that circadian cycles develop in the SCN around the time of birth in laboratory rodents. This has been shown by detecting core CRG expression in the SCN using in situ hybridization (Shimomura et al., 2001; Ohta et al., 2003; Sladek et al., 2004; Li and Davis, 2005; Kovacikova et al., 2006) and in older studies by the expression of other genes (Reppert and Uhl, 1987), by levels of metabolic and neural activity (Reppert and Schwartz, 1983; Shibata et al., 1983), and by perinatal entrainment studies (Davis and Mannion, 1988). In addition, rhythms in perinatal SCN explants have been measured in vitro using a bioluminescent reporter of CRG expression (Ohta et al., 2008). The tracking of bioluminescence from the same tissue over time is likely to be a more sensitive assay of rhythmicity than is sampling tissues at different times of day, but this method has not yet been used to determine the age at which the SCN can first express circadian rhythms. The current study was undertaken to determine when SCN explants from mice develop the capacity to express circadian rhythms using a bioluminescent reporter of Per2 expression (per2::luc mice). We found that SCN explants taken from mice between 16 and 17 days postfertilization (dpf), 2 to 3 d before birth, consistently expressed robust rhythms in vitro. Weaker rhythms were consistently expressed a day earlier (e15) but not 2 or 3 d earlier (e13 and e14). If tissues obtained on e14 were, however, left in culture long enough, rhythms spontaneously developed. The capacity to generate rhythms at this age was not seen in other brain areas, suggesting that the SCN is uniquely differentiated as a circadian pacemaker as soon as it is formed (Kabrita and Davis, 2008). Because the bioluminescence assay requires synchrony among cells within the explants, our results indicate that soon after the SCN is formed, SCN cells have the capacity to generate circadian rhythms and to synchronize through cell-cell interactions. The development of the SCN in vitro may provide a system with which to study the cellular interactions that are critical for the synchrony of cellular oscillations.
Materials and Methods
Animals
Female C57BL/6J mice were maintained on a 14:10 LD cycle and paired for 2 h beginning at lights-on with males homozygous for an mPeriod2::luciferase (PER2::LUC) fusion protein (Yoo et al., 2004; congenic on C57BL/6J). To investigate the role of vasoactive intestinal peptide (VIP), homozygous females (congenic on C57BL/6J) with a targeted null mutation of the VIP gene (Colwell et al., 2003) were mated with males homozygous for both transgenes (mper2::luc and Vip-/-). Matings were confirmed by the presence of vaginal plugs. The afternoon after a positive mating was considered to be 0.5 dpf, while the entire first 24 h was designated e0. All experiments were performed according to protocols approved by Northeastern University’s Institutional Animal Care and Use Committee and the U.S. National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Explant Harvest and Bioluminescence Recording
Pregnant mice were killed by cervical dislocation and their embryos dissected into 4 °C HBSS. In the first study (VIP-positive mice), 12 to 32 hypothalamic explants were taken from 3 to 6 litters at each of 6 ages (e13-e18). In a second study (VIP-deficient mice), 8 to 16 hypothalamic explants from 1 to 3 litters were taken at each of 5 ages (e14-e18). Explants (ventral hypothalamus containing SCN or other tissues as noted) were removed under a dissecting microscope using iridectomy scissors. Hypothalamic explants were separated at the midline, and unilateral explants were individually placed onto Millicell-CM support membranes (Millipore, Billerica, MA) floating on 1.2 mL of Lumicycle culture media (Pantazopoulos et al., 2010) in 35-mm culture dishes. In most cases (70%), only 1 unilateral explant was used from a fetus, but occasionally, when the number of fetuses was less, both explants from a single fetus were used. Dishes were coverslipped, sealed with silicone grease, and placed in a Lumicycle (Actimetrics, Wilmette, IL) maintained at 36 °C, in which bioluminescence was recorded at 10-min intervals/dish for at least 8 d. All explants were left undisturbed until the termination of recording.
Data Analysis
Baseline-corrected (24-h running average, unsmoothed) bioluminescence records and their X2 periodograms were exported from Lumicycle Data Analysis (Actimetrics) to Microsoft Excel. Initial analysis windows comprised days 2 to 7 of recording. Where applicable, subsequent windows were analyzed that also contained 5 d of recording. For measurements of baseline-corrected amplitude and peak-to-peak intervals, parabolas were fit to segments of the bioluminescence records where values remained above or below baseline long enough to represent candidate peaks and troughs (see Suppl. Fig. S1). Differences among group means were analyzed by t test or analysis of variance.
Results
First Expression of Circadian Oscillations in the Hypothalamus In Vitro
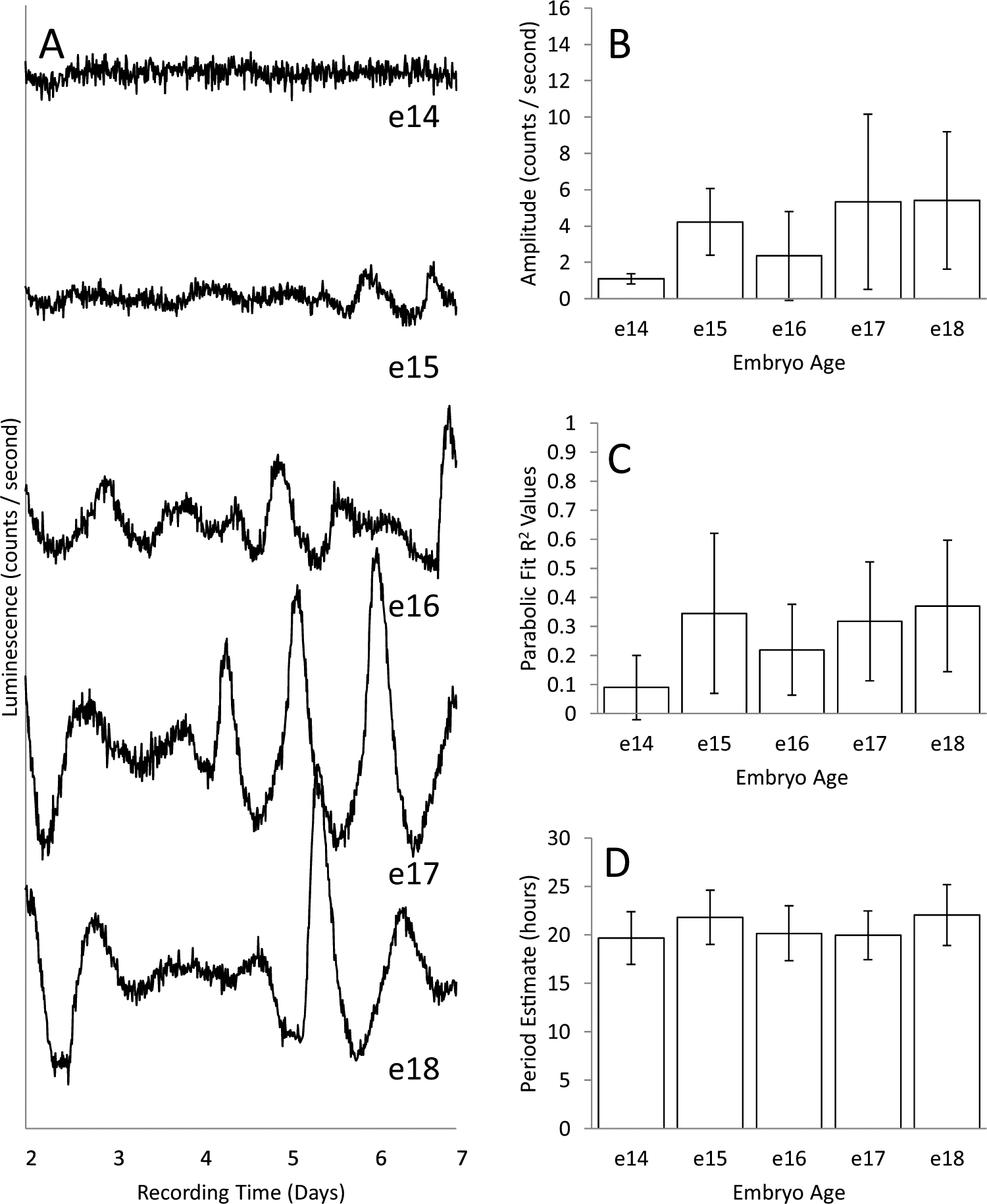
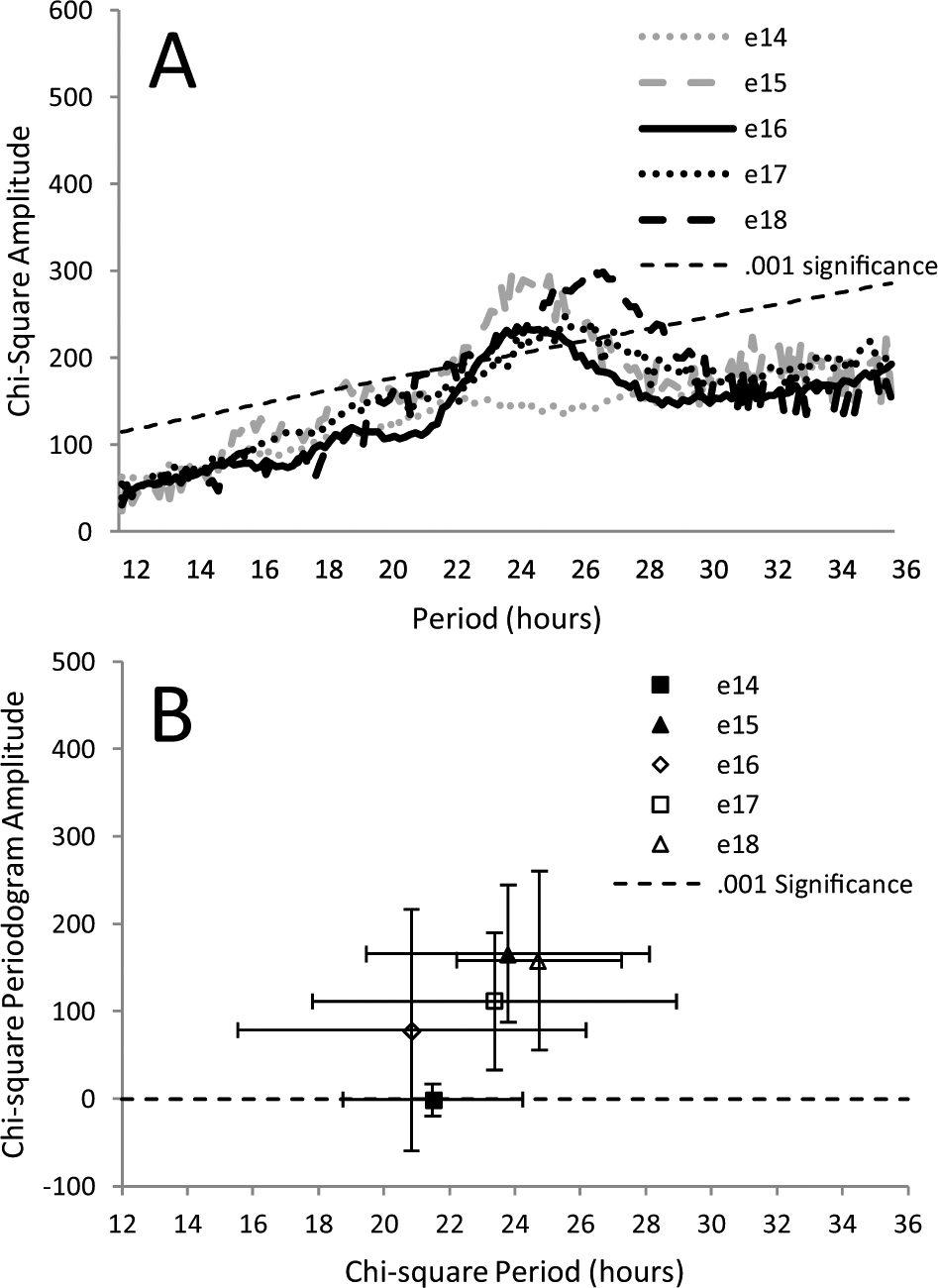
Figure 1A shows representative records of bioluminescence measured continuously for 5 d from embryonic suprachiasmatic hypothalamic (SCH) explants taken on different days of embryonic development (13-18 dpf, where 0 is the estimated time of fertilization and birth normally occurs 18.5-19.5 dpf). Explants obtained between 13 and 14 dpf are denoted as e13. Explants taken on e15 show a clear rhythm in bioluminescence, and the rhythms from older explants are more robust. The records for all explants are shown in Supplemental Figure S2. Quantification of rhythm parameters in Figure 1B-D and Figure 2 shows that the expressed rhythms become more robust between e15 and e18. In Figure 1B and C, this is seen in the mean amplitude and parabola goodness-of-fit (R2) of deflections from baseline (peaks and troughs), both of which increased with age (see Suppl. Fig. S1 for methods of analysis). In Figure 2A, averaged periodograms show an increase in power with age, indicating stronger expression of circadian periodicity at older ages. An increase in power and convergence on a circadian period after e15 is also supported by the means of periodogram peak values (Fig. 2B). The results for SCH explants are compared with periodograms from explants of cortex taken from the same embryos (Fig. 2C and D). There was little evidence of circadian rhythmicity in cortical explants at any age.
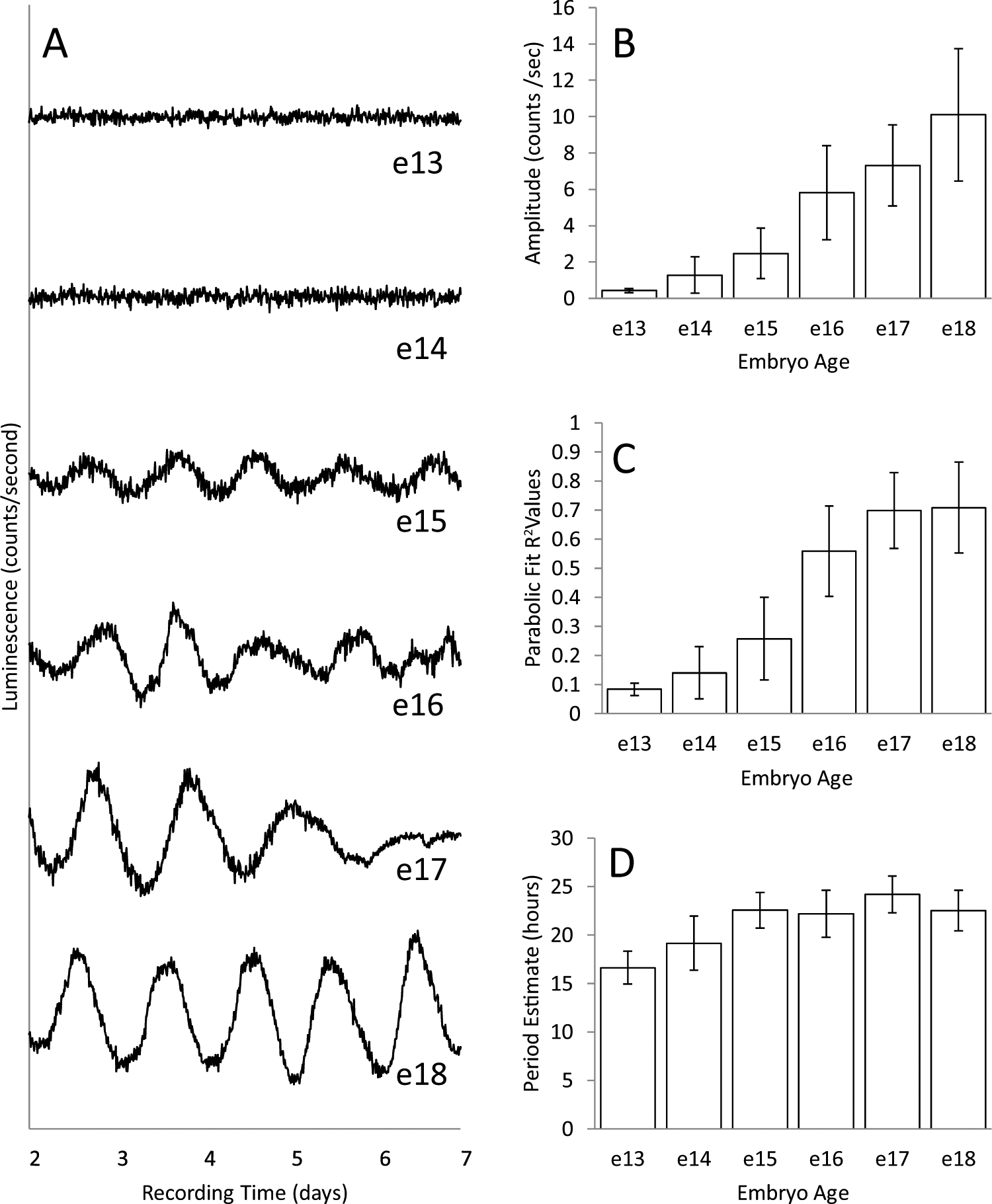
Baseline-subtracted records of PER2::LUC expression from representative explants collected on 6 successive days of prenatal development, embryonic ages e13 through e18 (A), and histograms summarizing the characteristics of records at each age (B-D). The total numbers of explants recorded were 14, 32, 12, 16, 16, and 20 for e13 to e18, respectively. Records for all explants are shown in Supplemental Figure S2. Histograms summarize values obtained by fitting parabolas to individual peaks and troughs as described in Supplemental Figure S1. Values within a record were averaged, and those means were then averaged for an embryonic age. Amplitude (B) is the deviation of peaks and troughs from baseline. Parabolic fit (C) is a measure of how distinct the peaks and troughs are in a record, and period estimate (D) is the average interval between successive peaks and troughs. Although some deviations from baseline were detected at e13 and e14, their amplitudes were very low and, in contrast to peaks and troughs at later ages, were poorly defined. Error bars are standard deviations, and differences among ages were highly significant for amplitude and parabolic fit (p < 0.001, analysis of variance [ANOVA]) but only marginally so for period estimates (ages e15-e18, p = 0.045, ANOVA).
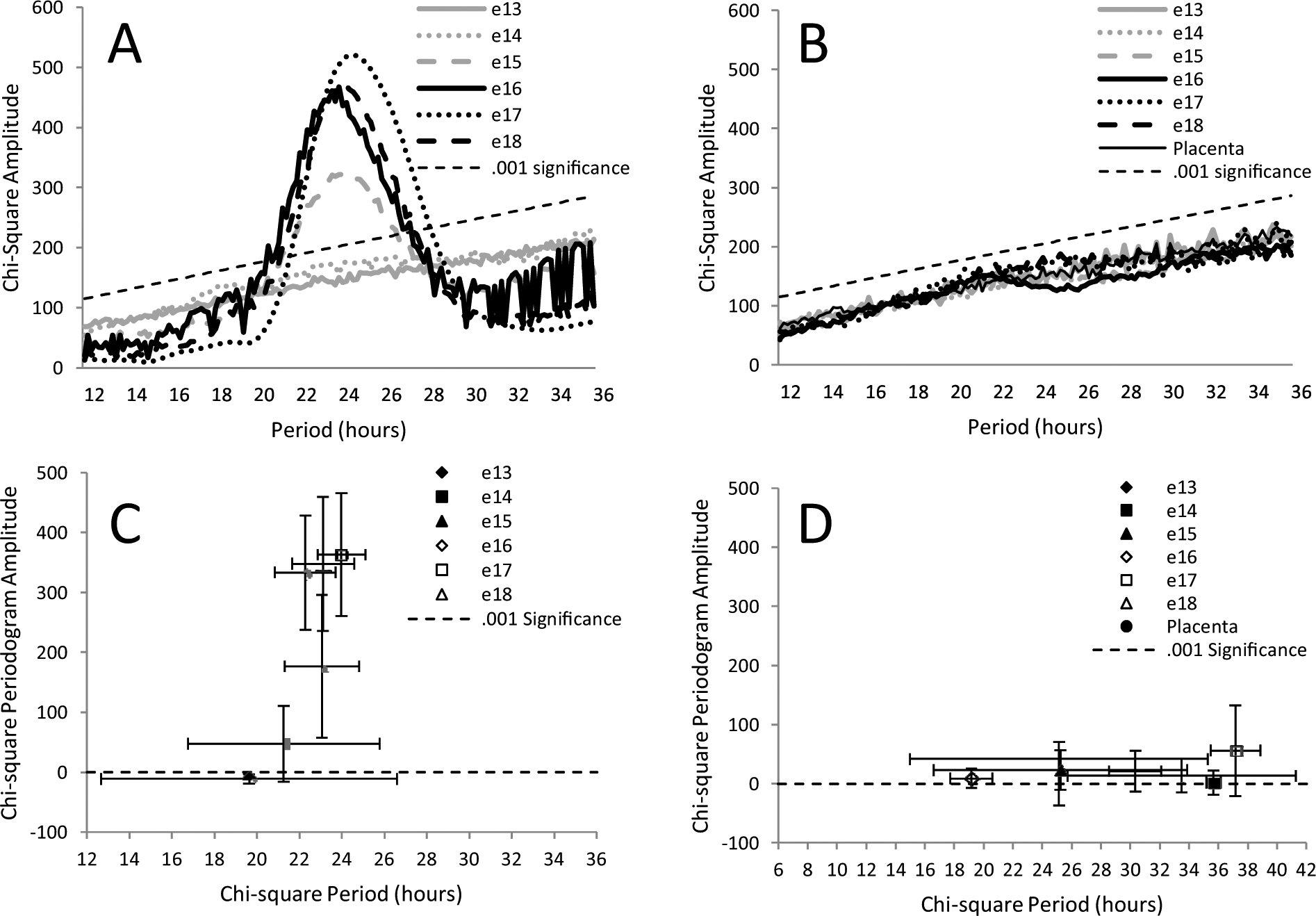
Average chi-square periodograms for records at each age, from either suprachiasmatic (SCH) (A) or cortical (B) explants. Included with cortex data is an average periodogram for 10 placental explants combined from all ages (no placenta at any age showed rhythmicity). The average peak amplitude of periodograms is plotted against the average period at which the peak amplitude occurred (C and D). Error bars are standard deviations. For e14 SCH explants (C) and some cortical explants (D), peak periodogram values rose above the 0.001 level of significance (dotted line). This is not reflected in the averaged periodograms because differences in period meant that the peaks of individual periodograms did not necessarily align when averaged. For SCH explants (C), both amplitude and period (e15-e18) are significantly different among ages (p < 0.001 and p = 0.01, analysis of variance).
Development of Circadian Oscillations Is a Distinct Feature of SCH Explants
As the data from the cortex in Figure 2 suggest, the ability to express circadian oscillations is not a property of all embryonic brain tissues. This was further examined by taking explants from areas around the SCH after it was removed for recording on e15. Explants anterior, lateral, and posterior to the SCH (as well as cortex) were taken from the same embryo and recorded. None of the other tissues expressed rhythms as distinct or persistent as those expressed by explants containing the SCN. This is illustrated by the averaged periodograms shown in Figure 3A. The lack of persistent rhythms in other embryonic brain tissues indicates that the expression of rhythms by the SCH reflects the development of unique functional properties.
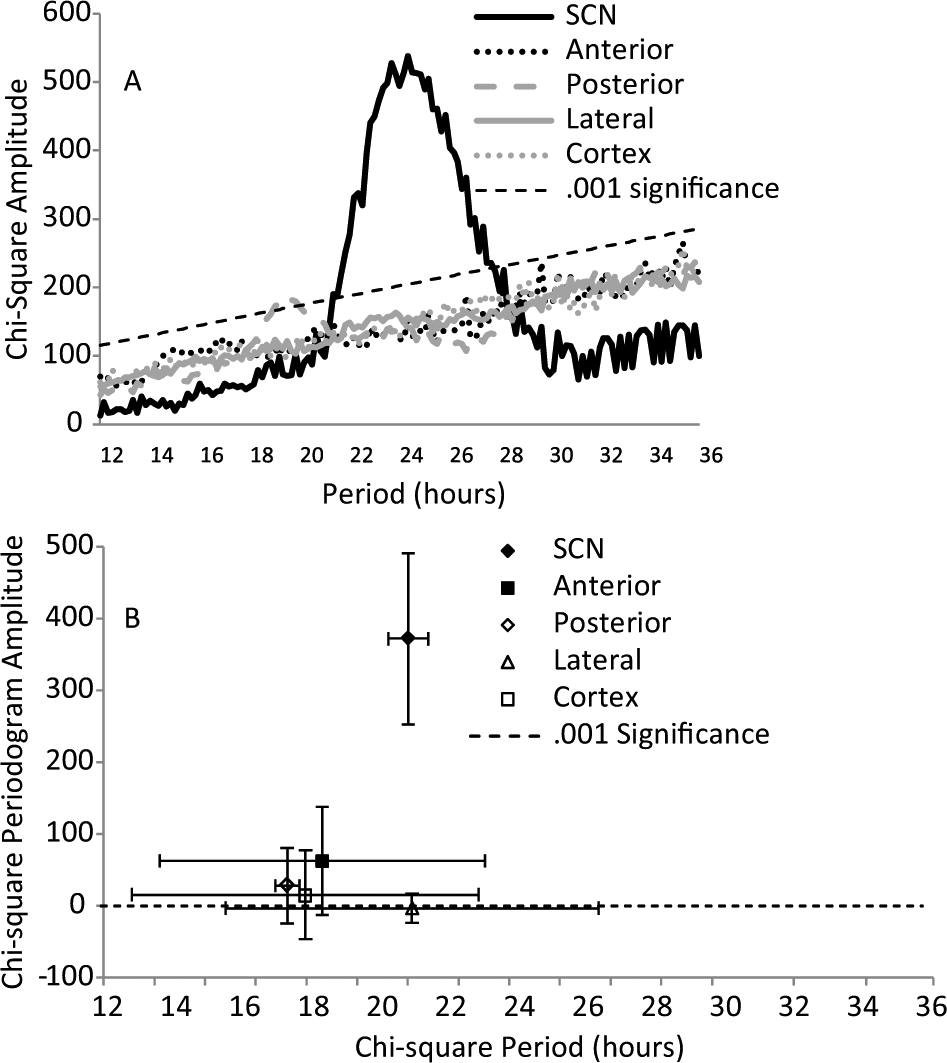
Average periodograms for PER2::LUC expression from explants of different brain areas obtained on e15 from 4 different fetuses. (A) After SCH explants were taken, similar-sized explants were taken from areas of the same fetal brain, anterior, posterior, and lateral to the SCN. Cortical explants were also taken. The average period of the periodogram peak is plotted against the average amplitude of the peak as in Figure 2 (B). Error bars are standard deviations.
Circadian Oscillations Develop In Vitro
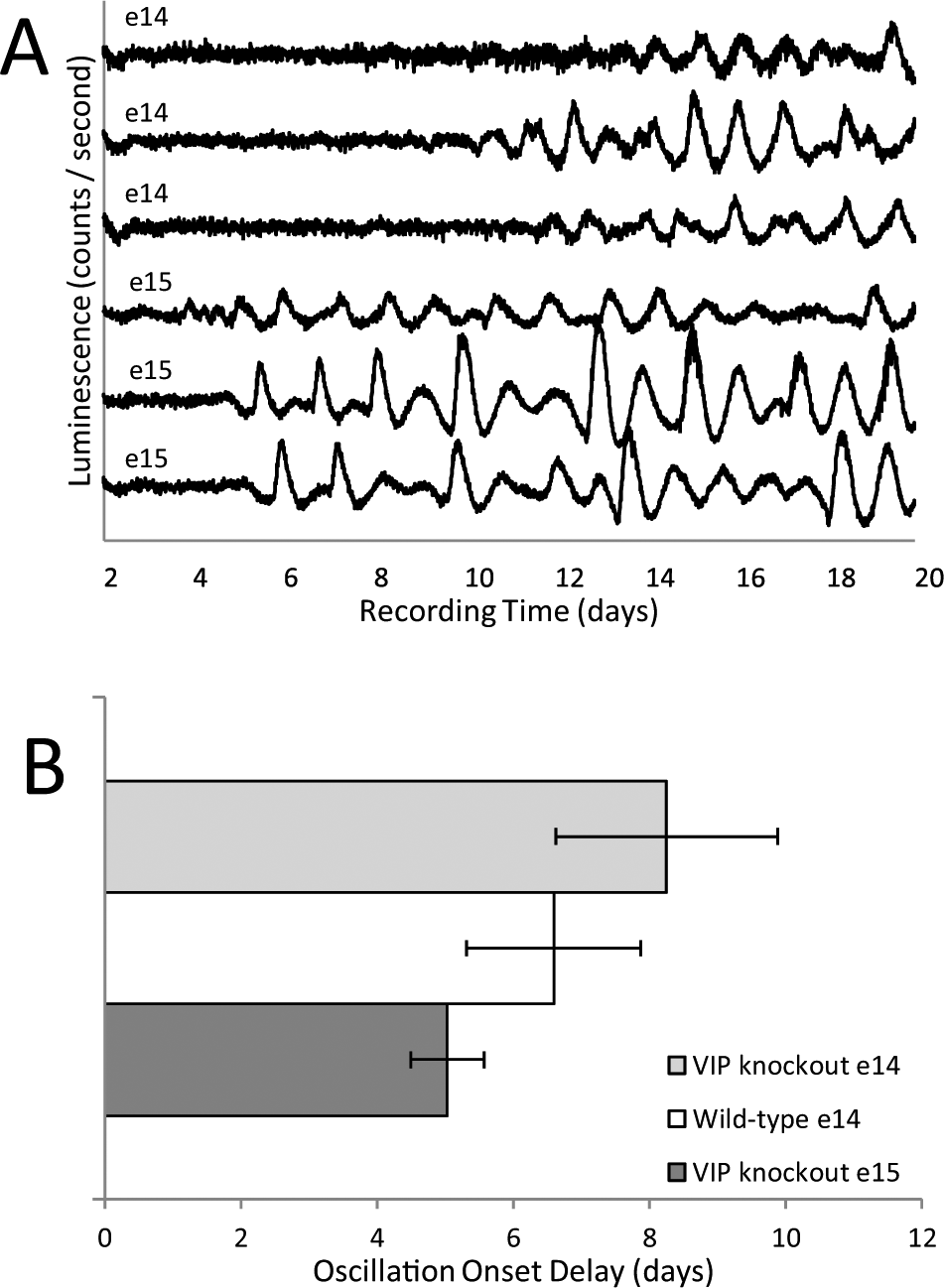
Circadian oscillations were rarely seen in explants from e14 in the first study. In that study, explants were recorded for 7 d. To determine whether oscillations would eventually develop, explants taken on e14 were cultured and recorded for 20 d. Figure 4 shows recordings from several explants that did not show rhythms for the first 5 to 7 d in culture but then gradually, or in some cases abruptly, began to express robust rhythms. The eventual expression of rhythms was seen in all 21 explants examined. The development of rhythms in explants in vitro was delayed relative to development of the SCH in vivo since 4 d in culture for an explant taken on e14 is the same age (in dpf) as an in vivo explant on e18, yet the e14 (+4) explants did not express rhythms while the e18 explants did (Fig. 1). The rhythms from e14 explants that appear in vitro continue to mature. The amplitude and goodness-of-fit of parabolas fit to identifiable peaks and troughs increased during 15 d in culture, approaching values characteristic of explants taken on e18. Even after 15 d, however, they were not equivalent to e18 explants (Suppl. Fig. S3).
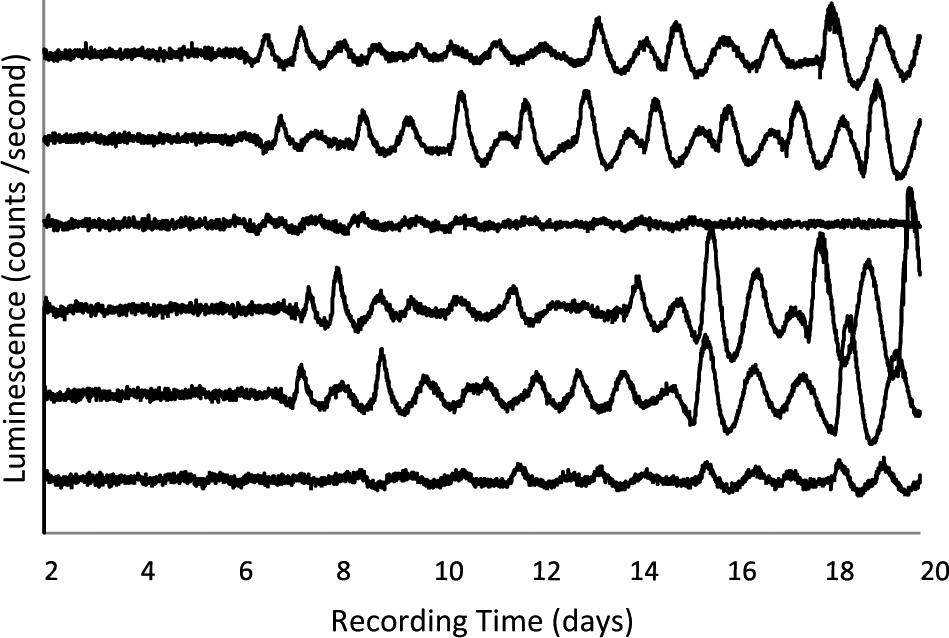
Baseline-subtracted records of PER2::LUC expression from representative explants collected on e14 and left undisturbed in culture for at least 20 d. No media changes or other disturbances occurred throughout the days of recording.
Circadian Oscillations Develop in the Absence of VIP
VIP is produced by a subset of SCN neurons with processes throughout the nucleus and into surrounding adjacent regions. VIP appears to be a secreted signal for coupling and synchrony among SCN neurons. The genetic deletion of VIP or one of its receptors (VPAC2) causes disruption of activity/rest rhythms, and within the SCN, these deficiencies cause a loss of synchrony among the rhythms of individual cells (Maywood et al., 2006). Application of VIP to SCN cells in vitro is sufficient to restore synchrony (Aton et al., 2005). Per2::luc mice have been crossed with mice deficient in VIP (VIP-/-), making it possible to assess PER2 expression in SCN that lack VIP. The rhythms from SCN explants of adult VIP -/- mice have lower amplitudes than do rhythms expressed by VIP-positive SCN (Loh et al., 2011; Maywood et al., 2011). The amplitudes of the damped rhythms can be restored by co-culture with a VIP-positive SCN, probably as a result of VIP secreted from the VIP-positive SCN (Maywood et al., 2011). Because VIP is a strong candidate for a physiologically important coupling signal within the SCN, it is possible that the development of coupling within the SCN is compromised by the loss of VIP. To test this, we measured the rhythms of SCN explants taken at different embryonic ages (e14-e18) from Per2::luc, VIP -/- double transgenic mice. We also measured the development of rhythms in e14 and e15 explants kept in vitro for 20 d. Figure 5A shows records from explants of each age and shows that explants from VIP-/- mice can express tissue-level rhythms in PER2 expression. Quantification of these rhythms in Figure 5B suggests that they are not as robust as are the rhythms from VIP-positive mice and do not show the same developmental progression toward more robust rhythms at older ages. Similarly, the periodogram analysis in Figure 6 shows that circadian periodicity in the rhythms of VIP-/- explants is never as strong as those from VIP-positive mice. The records of all VIP-/- SCH explants are shown in Supplemental Figure S4.
Baseline-subtracted records of PER2::LUC expression from representative suprachiasmatic hypothalamic explants collected from VIP-/- mice at 5 ages, e14 to e18 (A), and histograms summarizing the characteristics of records at each age (B-D). The numbers of explants recorded were 15, 8, 16, 16, and 8 for e14 to e18, respectively. Records for all explants are shown in Supplemental Figure S3. Histograms summarize values as describe in Figure 1. Error bars are standard deviations, and differences among ages were significant for amplitude and parabolic fit (p < 0.01, analysis of variance [ANOVA]) but were not significant for period estimates (ages e15-e18, p > 0.05, ANOVA).
Average chi-square periodograms for records from VIP-/- suprachiasmatic hypothalamic explants taken at 5 ages, e14 to e18 (A). As in Figure 2, the average peak amplitude of periodograms is plotted against the period at which the peak amplitude occurred (B). Error bars are standard deviations. Differences among ages were significant for amplitude (p < 0.001, analysis of variance [ANOVA]) but not for period (p > 0.05, ANOVA).
Long-term recordings of e14 (n = 14) and e15 (n = 8) explants from VIP-/- embryos confirm the ability of the SCH to develop rhythms in the absence of VIP. Figure 7A shows sample records from e14 and e15 explants taken from VIP-/- embryos. Rhythms are obvious, but they also show considerable variability both between and within explants. The development of rhythms in SCH explants may be delayed by the lack of VIP since explants from e14 VIP -/- embryos took longer to express circadian oscillations than did explants from VIP-positive embryos, and even e15 explants from VIP -/- embryos often showed a delay in the onset of rhythms (Fig. 7B). Similar to explants from VIP-positive explants, oscillations from VIP-/- explants all eventually developed some rhythmicity and became more robust with time in vitro (Suppl. Fig. S3).
Baseline-subtracted records of PER2::LUC expression from representative VIP-/- explants collected on e14 or e15 and left undisturbed in culture for at least 20 d (A). The numbers of days in culture before rhythms initiated are compared for wild-type explants obtained on e14 and VIP-/- explants obtained on e14 or e15 (B). The criteria for the first significant deflection (peak or trough) were stricter than criteria used in other analyses (Suppl. Fig. S1). Initial deflections consisted of values above baseline for at least 6 continuous hours and were followed by another deflection within 12 h. In some records in A, a “shoulder” is associated with a peak. In these cases, the shoulder and peak would have been fit by a single parabola, reducing the goodness-of-fit value.
Discussion
Recent reviews of circadian rhythm development in mammals focus on a timeline of structural and functional changes in the SCN (Davis and Reppert, 2001; Seron-Ferre et al., 2012; Sumova et al., 2012; Brooks and Canal, 2013). The current study adds to these the earliest age when the mouse SCN has a capacity to express circadian oscillations, at about 15 d after fertilization and 3 to 4 d before birth. The oscillations measured were in a reporter of protein levels from a gene that participates in the core molecular feedback loop, per2. While it is likely that the oscillations reflect the activities of other components of the core molecular clock as well, it has been shown that stochastic oscillations in PER2::LUC can emerge in explants from mice lacking a molecular circadian clock (bmal1 -/-; Ko et al., 2010). Noise in PER2::LUC expression combined with cellular coupling within the SCN appears to generate variable, short period oscillations (mean = 17.8 h). While we cannot exclude the absence of cellular oscillations in e15 explants, the oscillations we observed at that age were already similar to those on e18 with respect to mean period (22.55 and 22.52 h) and to within-record peak-to-peak variation (p > 0.05 for F-test comparison of variances). Although less robust than oscillations from adult explants, oscillations from embryonic explants on e15 and older resembled more the oscillations expressed by explants from mice with functional cellular clocks.
The mouse SCN is formed from postmitotic cells produced by a progenitor population in the germinal epithelium of the third ventricle between 12 and 15 dpf (Kabrita and Davis, 2008). Therefore, by 15.5 dpf, when we consistently observed oscillations in vitro, SCN cells are postmitotic. Also at this time, SCN cells appear to become specified by temporal and spatial patterns of transcription factor expression in the ventral hypothalamus. At the age when the SCN is first able to express oscillations in vitro (15.5 dpf), the SCN can be identified by the expressions of Six3, Lhx1, and Rorα (VanDunk et al., 2011). Lhx1 and Rorα are most specific to the SCN, and their expression is compromised by a targeted mutation in Six3, which is expressed earlier and over a broader area. Both the Six3 mutation and a mutation in Six6, another homeobox transcription factor expressed within the developing ventral hypothalamus, result in the failure of SCN neurons to develop characteristic peptide expression or to form an anatomically distinct nucleus (Clark et al., 2013). The early definition of the SCN by Lhx1 and Rorα expression precedes the expression of mRNAs for peptides that characterize the SCN (VIP, AVP), suggesting that those transcription factors are involved in the postmitotic differentiation of SCN cells (VanDunk et al., 2011). In support of this, compromised expression of Lhx1 in the anterior ventral hypothalamus (Six3-Cre;Lhx1lox/lox mice) causes reduced expression of SCN peptides and disrupted behavioral rhythms. The nucleus itself is formed and expresses rhythms in core CRGs (e.g., Per2). It was suggested that SCN-specific genes expressed before e16.5 are not dependent on Lhx1 expression (Bedont et al., 2014). Based on our results, this could include the initial expression of Per2 and possibly other genes required to drive its oscillations. Although a mutation of Rorα did not affect postnatal peptide expression or SCN anatomy (VanDunk et al., 2011), early differentiation could have been delayed and/or transcription factor redundancies could have compensated. While specification of the SCN during development is likely to result from a cascade of transcription factor expression before and during SCN progenitor proliferation, e15 and the end of neurogenesis appears to be a critical time for SCN differentiation. This time might also be the point at which the core feedback loop consisting of transcriptional activation of Per2 by CLOCK/BMAL1 and negative feedback from PER2 (as well as PER1, CRY1, and CRY2) initiate. RORα is a transcriptional activator of bmal1 and could play a role in the initiation of the first oscillations (Sato et al., 2004; Akashi and Takumi, 2005). Our demonstration that regions surrounding the suprachiasmatic region did not have the capacity to express circadian oscillations on e15 supports the conclusion that the SCN’s ability to generate circadian oscillations at this age is a property specific to SCN development and differentiation.
Before e15, the mouse SCN does not consistently express circadian oscillations detectable by the bioluminescent reporter used here. It is possible that circadian oscillations are expressed earlier in individual cells but that the cells are few in number and/or are not synchronized. Alternatively, the expression of oscillations we observed in vitro could represent an abnormally early onset of oscillations, earlier, for example, than the onset that normally occurs in vivo. Thus far, the earliest age for SCN rhythms in vivo detected by in situ hybridization in mice is e17 for per1 and postnatal day 6 for per2 (Shimomura et al., 2001). Immunohistochemistry revealed rhythms in PER1 and PER2 (but not other CRGs) on e18 (Ansari et al., 2009). In rats, one study using in situ hybridization detected the expression of several CRGs in the e19 SCN but did not see rhythms until the next age examined, postnatal day 3 (Sladek et al., 2004). A subsequent study detected a weak per1 rhythm on e20, but rhythms in other CRGs were seen only after birth (Kovacikova et al., 2006). Another study reported rhythms in both per1 and per2 on E20, 1 to 2 d before birth (Ohta et al., 2003) and did not examine earlier ages or other genes. An in vitro rhythm using a bioluminescent reporter driven by the per1 promoter was also seen at this age in rats (Ohta et al., 2008). In the present study, placing tissue in culture may have artificially initiated and/or synchronized oscillations; fetal liver, heart, and kidney express rhythms in vitro that cannot be detected by polymerase chain reaction in vivo (Dolatshad et al., 2010). Even if the procedure stimulates the expression of rhythms, the effect is specific to the SCN and to a particular point in development, since the same procedure did not induce rhythms in other brain areas or in the SCN at earlier ages (e13 or e14). If the phase of in vitro oscillations were related to maternal rhythms or to other entraining signals present before isolation for culture, this would support the presence of entrainable oscillations at an even earlier age. Because our procedure had a strong phase-setting effect on the subsequent in vitro oscillations (Suppl. Fig. S5), we could not assess phase for evidence of prior entrainment.
When tissues obtained on e14 were left in culture long enough, they began to express circadian oscillations. It is possible that if tissues are placed in culture before they have the capacity to generate oscillations, oscillations develop according to a program resembling one that occurs in vivo. This would suggest that the development of oscillations in vivo is due to a tissue autonomous program that can occur in isolation without signals from elsewhere in the embryo or from the mother. This is supported by the fact that embryonic SCN transplanted into adult host brains eventually generate synchronized oscillations sufficient to drive the host’s behavior (Ralph et al., 1990; Kaufman and Menaker, 1993). The development of oscillations in mouse embryonic fibroblasts and submandibular glands in vitro also suggests autonomous development (Inada et al., 2014). Similar to the delayed in vitro development of rhythms in submandibular glands, e14 SCN expressed oscillations later than they would if isolated at an older age, equivalent to about 20 to 21 dpf. SCN isolated on e15 and later show robust rhythms immediately. Thus, it is likely that development does not proceed in vitro as it would in vivo.
In tissues other than the SCN or in cell lines, it is common for individual oscillating cells to drift out of synchrony over time in culture. The opposite occurred in embryonic SCN isolated on e14. The explants did not require additional synchronizing stimuli in order to develop oscillations, indicating that coupling mechanisms sufficient to synchronize cells develop autonomously in vitro. Interestingly, we observed the spontaneous development of oscillations even in tissues that do not express VIP. VIP is currently thought to be an important signal for coupling among SCN neurons, and some differences in robustness between embryonic wild-type and VIP-/- SCN were seen, similar to reports for in vitro SCN from older animals (Loh et al., 2011; Maywood et al., 2011; An et al., 2013). Our results show, however, that the development of circadian rhythms in vitro is not dependent on VIP. Rather than having greater dependence on known coupling signals, the fetal and neonatal SCN may be less dependent. The developing SCN appears hypersensitive to phase-setting signals (Viswanathan and Davis, 1997; Nishide et al., 2008) and may be less dependent on mechanisms required for coupling in adults; neonatal SCN appear to use an unknown diffusible coupling mechanism that is lost during development. Neonatal SCN from mice without functional cryptochrome genes (Cry1-/- and Cry2-/-) still express rhythms in vitro, but adult SCN do not, suggesting that the development of adult coupling mechanisms depends on functional cry (Ono et al., 2013).
The present results establish the earliest age for the onset of autonomous circadian oscillations in the developing SCN. Although the onset of oscillations as early as e15 has yet to be established in vivo, it is likely that the SCN begins to function as a circadian pacemaker within a few days of its genesis. In humans, hypothalamic neurogenesis, including midline structures such as the SCN, is probably complete by the end of the first trimester (13 wk), when few mitotic figures are observed in the neuroepithelium (Koutcherov et al., 2002). This is consistent with comparisons of development in other mammals (van Eerdenburg and Rakic, 1994; Otis and Brent, 1952). Early within the second trimester (17-24 wk), the SCN shows characteristics of differentiation such as the expression of melatonin receptors, distinct subnuclei, and innervation by retinal axons (Reppert et al., 1988; Koutcherov et al., 2002; Thomas et al., 2002). In nonhuman primates, the SCN shows functional responses to light at a prenatal age equivalent to 25 wk in humans, and rhythms in SCN metabolic activity and CRG expression have been seen late in gestation (the earliest age examined; Reppert and Schwartz, 1984; Hao and Rivkees, 1999; Torres-Farfan et al., 2006). The early formation of the human SCN and the capacity for circadian oscillations soon after formation, as demonstrated here, indicate that the human SCN could be functional at the start of the second trimester, 26 wk before birth. The observations in rodents that several drugs can induce gene expression in the SCN raise the possibility that for much of gestation, the human fetal SCN is susceptible to perturbations affecting gene expression and possibly its normal development (Weaver et al., 1992; Clegg et al., 1995; Bender et al., 1997; Shearman and Weaver, 2001).
Footnotes
Acknowledgements
The authors thank Mary Harrington for providing per2::luc knock in mice with permission from Joe Takahashi and Erik Herzog for providing double transgenic mice (vip-/-; per2::luc). This work was supported by National Institutes of Health grant MH068796 to F.C.D. and by a Northeastern University Tier I Provost Grant. The authors thank Mary Harrington, Penny Molyneux, Andrew Cary, and Harry Pantazopoulos for technical assistance.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.