
Abstract
Whether the circatidal rhythm is generated by a machinery common to the circadian clock is one of the important and interesting questions in chronobiology. The mangrove cricket Apteronemobius asahinai shows a circatidal rhythm generating active and inactive phases and a circadian rhythm modifying the circatidal rhythm by inhibiting activity during the subjective day simultaneously. In the previous study, RNA interference of the circadian clock gene period disrupted the circadian rhythm but not the circatidal rhythm, suggesting a difference in molecular mechanisms between the circatidal and circadian rhythms. In the present study, to compare the neural mechanisms of these 2 rhythms, we observed locomotor activity in the mangrove cricket after surgical removal of the optic lobe, which has been shown to be the locus of the circadian clock in other crickets. We also noted the pigment-dispersing factor immunoreactive neurons (PDF-IRNs) in the optic lobe, because PDF is a key output molecule in the circadian clock system in some insects. The results showed that the circadian modulation was disrupted after the removal of the optic lobes but that the circatidal rhythm was maintained with no remarkable changes in its free-running period. Even in crickets in which some PDF-immunoreactive somata remained after removal of the optic lobe, the circadian rhythm was completely disrupted. The remnants of PDF-IRNs were not correlated to the occurrence and free-running period of the circatidal rhythm. These results indicate that the principal circatidal clock is located in a region(s) different from the optic lobe, whereas the circadian clock is located in the optic lobe, as in other crickets, and PDF-IRNs are not important for circatidal rhythm. Therefore, it is suggested that the circatidal rhythm of A. asahinai is driven by a neural basis different from that driving the circadian rhythm.
Keywords
Organisms show endogenous rhythms corresponding to cyclic changes in the external environment (Dunlap et al., 2004). Because the habitats of nearly all organisms are influenced by the day-night cycles, the circadian rhythm with a period of approximately 24 h is a common feature in a wide variety of organisms. In addition to the day-night cycles, intertidal zones are also influenced by tidal flooding and ebbing with a period of approximately 12.4 h, and this circatidal rhythm has been reported mainly in intertidal species (Palmer, 1995; Naylor, 2010). Physiological mechanisms generating the circadian rhythm have been studied in a wide variety of organisms; however, those generating the circatidal rhythm have been much less studied (Wilcockson and Zhang, 2008; Naylor, 2010).
Physiological mechanisms generating the circatidal rhythms have been investigated mainly in crustaceans (Palmer, 1995; Naylor, 2010). For instance, in the shore crab Carcinus maenas, a neuroendocrine system in the eyestalk is considered to be a mediator between the pacemaker and the circatidal locomotor activity rhythm (Naylor and Williams, 1968). In the estuarine crab Sesarma haematocheir, which shows circatidal hatching rhythms, surgical ablations have suggested that the pacemaker neurons or neurons under the control of the pacemaker are located in the medulla terminalis in optic peduncles of the eyestalk (Saigusa, 2002). The eyestalk is also considered to be one of the loci of the circadian clocks distributed in central nervous systems, and its neuroendocrine system is involved in circadian regulation in crustaceans (Aréchiga et al., 1993; Strauss and Dircksen, 2010). Together with the reports that the circatidal and circadian neurosecretory rhythms were sustained in the isolated eyestalks of C. maenas and of the crayfish Procambarus clarkii, respectively (Williams et al., 1979; Rodríguez-Sosa et al., 1994), it has been determined that the eyestalk is important for both the circatidal and circadian rhythms in crustacea and that the circatidal and circadian clock systems might share some neuronal components in the eyestalk.
In the mangrove cricket Apteronemobius asahinai, which, to date, is the only insect known to show a clear and persistent circatidal locomotor activity rhythm under constant conditions, the molecular mechanism generating this rhythm is suggested to be different from that of the circadian rhythm. In A. asahinai, the circatidal and circadian rhythms are shown simultaneously in its locomotor activity under constant conditions. The circatidal rhythm generates active and inactive phases, and the circadian rhythm modifies the circatidal rhythm by inhibiting activity during the subjective day (Satoh et al., 2008). This circatidal rhythm is considered to be generated by a circatidal clock independent from the circadian clock because the circatidal rhythm entrains to periodic water inundations, but not to light-dark cycles, and shows a phase response curve with a period of approximately 12.4 h (Satoh et al., 2008, 2009). The circadian modulation, however, entrains to light-dark cycles. RNA interference (RNAi) of the circadian clock gene period disrupted only the circadian modification and caused no effects on the circatidal rhythm (Takekata et al., 2012, in press), suggesting an independent circatidal clock from the circadian clock. Although the results of period knocking down suggest the circatidal rhythm is considered to be generated by a molecular mechanism different from that of the circadian rhythm, the neural components of the 2 rhythms are still unknown (Takekata et al., 2012).
In the present study, to compare neural tissues important for the circatidal and circadian rhythms in A. asahinai, we observed the effects of surgical removal of the optic lobe on the locomotor rhythms. In insects, the principal circadian clock has been localized in the brain, and the optic lobe of the brain is the circadian clock locus for crickets, cockroaches, and beetles (see Tomioka and Matsumoto, 2010, for review). In crickets, the role of the optic lobe was demonstrated by the culture experiment of the optic lobe in Gryllus bimaculatus (Tomioka and Chiba, 1992), and the removal of the optic lobes or severance of the optic stalk causes arrhythmicity in Teleogryllus commodus, Gryllodes sigillatus, and Dianemobius nigrofasciatus (Loher, 1972; Sokolove and Loher, 1975; Abe et al., 1997; Shiga et al., 1999). We have paid particular attention to pigment-dispersing factor immunoreactive neurons (PDF-IRNs) in the optic lobe. PDF is a key output molecule in circadian clock systems in insects and is involved in regulating the circadian locomotor activity rhythm (Renn et al., 1999; Lee et al., 2009; Hassaneen et al., 2011). In hemimetabolous insects, 3 distinct clusters of PDF-IRNs are located in the optic lobe (Homberg et al., 1991; Sehadová et al., 2003).
The results showed that circadian modulation was disrupted after removal of the optic lobes, but the circatidal rhythm was maintained with no remarkable changes in its free-running period, even in crickets with the 3 clusters of PDF-IRNs completely removed. Thus, the principal circatidal clock is located in a region different from the optic lobe; therefore, the circatidal rhythm of A. asahinai would consist of a neural basis different from that of the circadian rhythm.
Materials and Methods
Insects
Adult males of A. asahinai (body length 5.5-6.2 mm, head width including eyes 1.7-1.9 mm; Yamasaki, 1979) were collected from mangrove swamps in Ginoza (26°30′N, 127°59′E), Okinawa Prefecture, Japan, on the days around the spring tide in July and October, 2010-2011. Activity recordings from the day of collection were made using some of the collected males, which were also subsequently used for immunocytochemistry. The remaining insects were used in hematoxylin-eosin staining of brain sections.
Surgical Operation
The optic lobes were removed by the surgical operation. Before surgery, crickets were anesthetized using CO2 for approximately 5 sec, immediately mounted in a sealing compound with the frontal surface of the head exposed, and placed on ice (approximately 0 °C) for 5 min to harden the sealing compound. Under a stereoscopic microscope, the compound eyes were cut off under 0.9% NaCl using a razor blade. Connections of the optic lobe and central brain were severed using small scissors, and the optic lobes were taken off. The openings were then covered with their own compound eyes using dental paraffin wax (melting point, 59.3 °C; GC Corporation, Tokyo, Japan). For the sham operation, the frontal margin of the compound eye was incised with a blade, and dental paraffin wax was placed over it. The operations were performed either unilaterally or bilaterally.
Histological Observations
The brains from field-collected males without activity recordings were examined by hematoxylin-eosin staining. After collection, crickets were maintained at 25.0 ± 1.0 °C under light 16 h:dark 8 h (LD 16:8) conditions with dried insect pellets (Oriental Yeast, Tokyo, Japan) and slices of carrot. A piece of moist cotton was provided as a water source, and the relative humidity was maintained at >50%. Operations were performed within 10 days of collection. Fifteen days after the operations, the survivors were anesthetized using CO2 for approximately 5 sec, and their heads were removed. After fixation in aqueous Bouin, the heads were processed for paraffin sections (8-µm thick). The sections were stained with Mayer’s hematoxylin and eosin.
Activity Recording and Analysis
Activity was recorded as described previously (Takekata et al., 2012). Briefly, crickets were individually housed in a plastic chamber with a sufficient supply of food and water. The interruptions of an infrared beam (EE SPW-321, Omron, Kyoto, Japan) set across the chamber were recorded at 6-min intervals on a personal computer. Lighting schedules were controlled by the same computer as was used for recording activity. The recording chambers were set in an incubator maintained at 25 °C ± 1.0 °C and equipped with a 15-W fluorescent lamp (FL15 W, Panasonic, Kadoma, Japan) with an irradiance of 1.0 to 3.5 W/m2. Operations were performed on crickets after recording activity for 4 to 6 days under LD 12:12, and their activities were recorded again under LD 12:12 for 4 to 5 days and under constant darkness (DD) for a subsequent 10 days.
Under DD, circatidal rhythmicity was determined using a chi-square periodogram (Sokolove and Bushell, 1978). The activity with a clear single peak above the 0.05 confidence level between 10 and 15 h in the periodogram was judged to be circatidal. In only the crickets judged to be circatidal was the presence of circadian rhythmicity examined by comparison of activity levels between the neighboring circatidal cycles. The number of infrared beam interruptions was summed for every circatidal period. A period starting 12 h after the final light off, which is the time predicted to be the start of the subjective day, was designated as the first circatidal cycle. Thus, the first and the second circatidal cycles approximately coincided with the subjective day and night, respectively, because the circatidal period is approximately half of the circadian period. If a circadian modulation exists in an activity, there should be consistent differences in the activity levels between the odd- and even-numbered circatidal cycles through the experimental period. When a circadian free-running period is almost twice the circatidal period, the means of activity levels are different between the odd- and even-numbered circatidal cycles because the circadian inactive and active phases coincide with odd- and even-numbered circatidal cycles, respectively. When the circadian free-running period is largely different from twice the circatidal period, the circadian inactive phase gradually deviates from the odd-numbered circatidal cycles and the circadian active phase from the even-numbered cycles day by day. Therefore, the activity levels in the odd-numbered cycles increase and those in the even-numbered cycles decrease during the experimental period, and thus the slopes of the regression lines are different between even- and odd-numbered cycles. In contrast, if a cricket shows only the circatidal rhythm, the trend of the activity levels should be the same between odd- and even-numbered circatidal cycles, and there is no difference both in the slope of regression lines and the means of activity levels. The difference in odd- and even-numbered cycles was examined by ANCOVA with the assumption that the activity levels regress linearly. The activity was judged to be influenced by a circadian rhythm when the slopes of regression lines or the means of the activities were significantly different between the odd- and even-numbered circatidal cycles (p ≤ 0.05; for details, see fig. S1 in Takekata et al., 2012). This method examines the presence of the circadian modulation only. We did not determine the circadian period. It is difficult to detect the circadian free-running period by chi-squire periodogram because multiples of the circatidal period are often fused with the circadian peaks.
Immunocytochemistry
At the end of the experiment, all crickets that had bilateral optic lobes removed and some of the intact and sham-operated crickets were sacrificed for PDF immunocytochemistry using the ABC method (Vector Laboratories, Burlingame, CA, USA). The rabbit anti–G. bimaculatus–PDF antiserum was provided by Dr. K. Tomioka (Okayama University, Okayama Prefecture, Japan). The crickets were anesthetized using CO2 immediately after recording their activity. The head was cut off and the posterior cuticle was removed to expose the brains to fixatives. The head was fixed in 4% paraformaldehyde for 4 h at 4 °C. After washing in phosphate-buffered saline (PBS; pH = 7.2) with 0.5% Triton X-100 (PBST) for 1 h at room temperature (RT), the brain was excised from the head in PBS. The brain was washed with PBST and incubated in 0.3% H2O2 for 1 h at RT to reduce endogenous peroxidase activity. The brain was then incubated in 2% normal goat serum in PBST with the primary anti-PDF antiserum at a dilution of 1:20,000 for 2 days at 4 °C. After washing with PBST, the primary antiserum-treated brain was incubated in the biotinylated goat anti-rabbit IgG secondary antibody for 1 day at 4 °C. After washing with PBST, the brain was incubated in an avidin-biotin complex solution at a dilution of 1:100 for 1 day at 4 °C. After washing in Tris-HCl (pH = 7.4), the brain was preincubated in 0.03% diaminobenzidine (Sigma-Aldrich Co, LLC, St. Louis, MO, USA) in Tris-HCl for 1 h at RT and incubated in a mixture of 0.01% H2O2 and 0.03% diaminobenzidine for approximately 30 sec at RT. After washing with Tris-HCl, the brain was dehydrated in an ethanol series and cleared in methyl salicylate for observation. The immunoreactive neurons were photographed using a digital microscope camera (DS-Ri1; Nikon, Tokyo, Japan), and the lengths of the major axis of their somata were determined using imaging software (NIS-Elements; Nikon, Tokyo, Japan).
Results
In intact crickets, 3 distinct neuropils of the lamina, medulla, and lobula were found in the optic lobe, which is closely connected to the central brain (Fig. 1A). In the crickets from which the optic lobes were bilaterally removed, the lamina and the medulla were completely absent, whereas a remnant of the lobula sometimes remained (Fig. 1B).
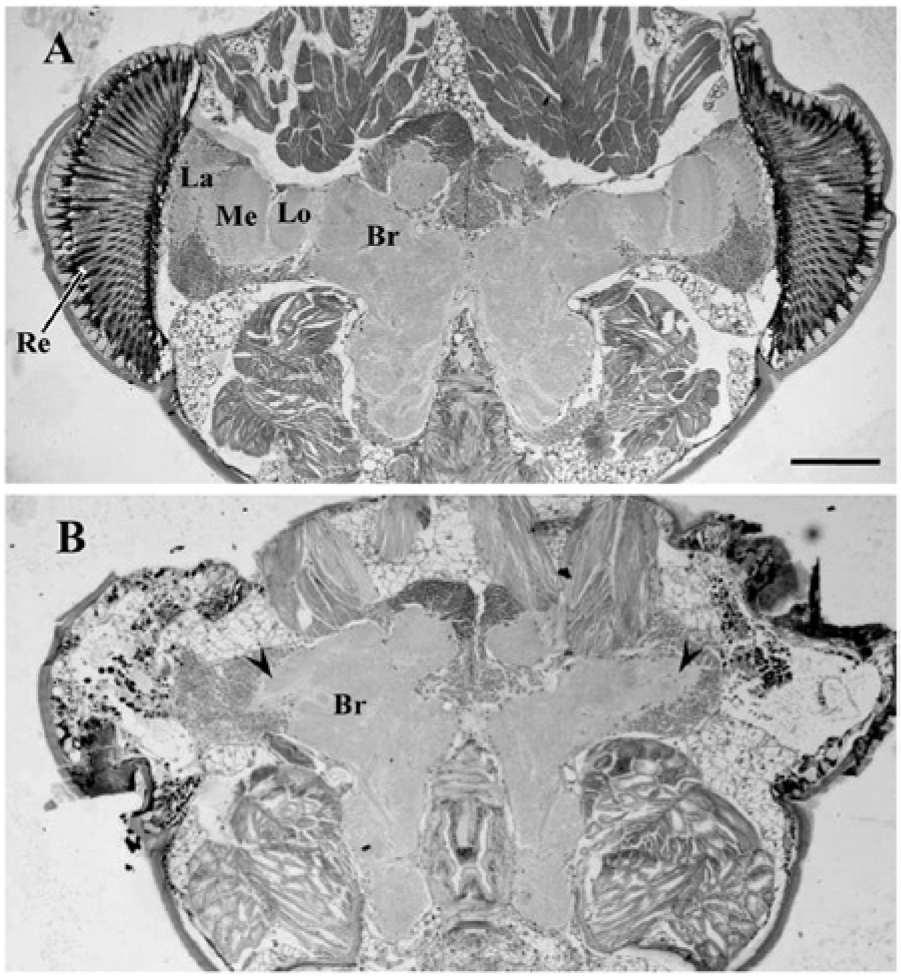
Representative sections of the head of Apteronemobius asahinai, including the optic lobes and central brain, stained with the hematoxylin-eosin, kept intact (A) or with optic lobes removed (B). In intact crickets, the optic lobe is closely connected to the central brain (Br). The lamina (La), medulla (Me), and lobula (Lo) underlie the retina (Re). In the head with the optic lobes bilaterally removed, the lamina and medulla were completely removed. The remnant of the lobula (arrowheads) bilaterally remained. Scale bar, 50 µm.
In the activity recordings, 17 of 18 intact, 13 of 21 unilaterally sham-operated, 15 of 20 bilaterally sham-operated, 16 of 31 unilaterally removed, and 24 of 41 bilaterally removed crickets survived for the entire experimental period. Figure 2 shows representative locomotor activities. An intact cricket showed a circatidal activity rhythm with a period of 12.9 h under DD (Fig. 2A). Circadian modulation of the activity was also observed under DD in the different activity levels between odd- and even-numbered circatidal cycles, and the means of the activity counts in the even-numbered cycles, of which components had continued from the scotophase of preceding LD cycles, were significantly higher than those in the odd numbered cycles, of which components had continued from the photophase (Fig. 2A). A sham-operated cricket also showed a clear circatidal rhythm with a period of 12.9 h and a clear circadian modulation (Fig. 2B), but circadian modulation was not detected in another sham-operated cricket (Fig. 2C). Figures 2D and E show two representative crickets with bilateral optic lobes removed. Both also showed clear circatidal activity rhythms with free-running periods of 13.0 or 13.2 h. However, there was no difference in activity levels between the adjacent circatidal cycles; hence, circadian modulation was lost.
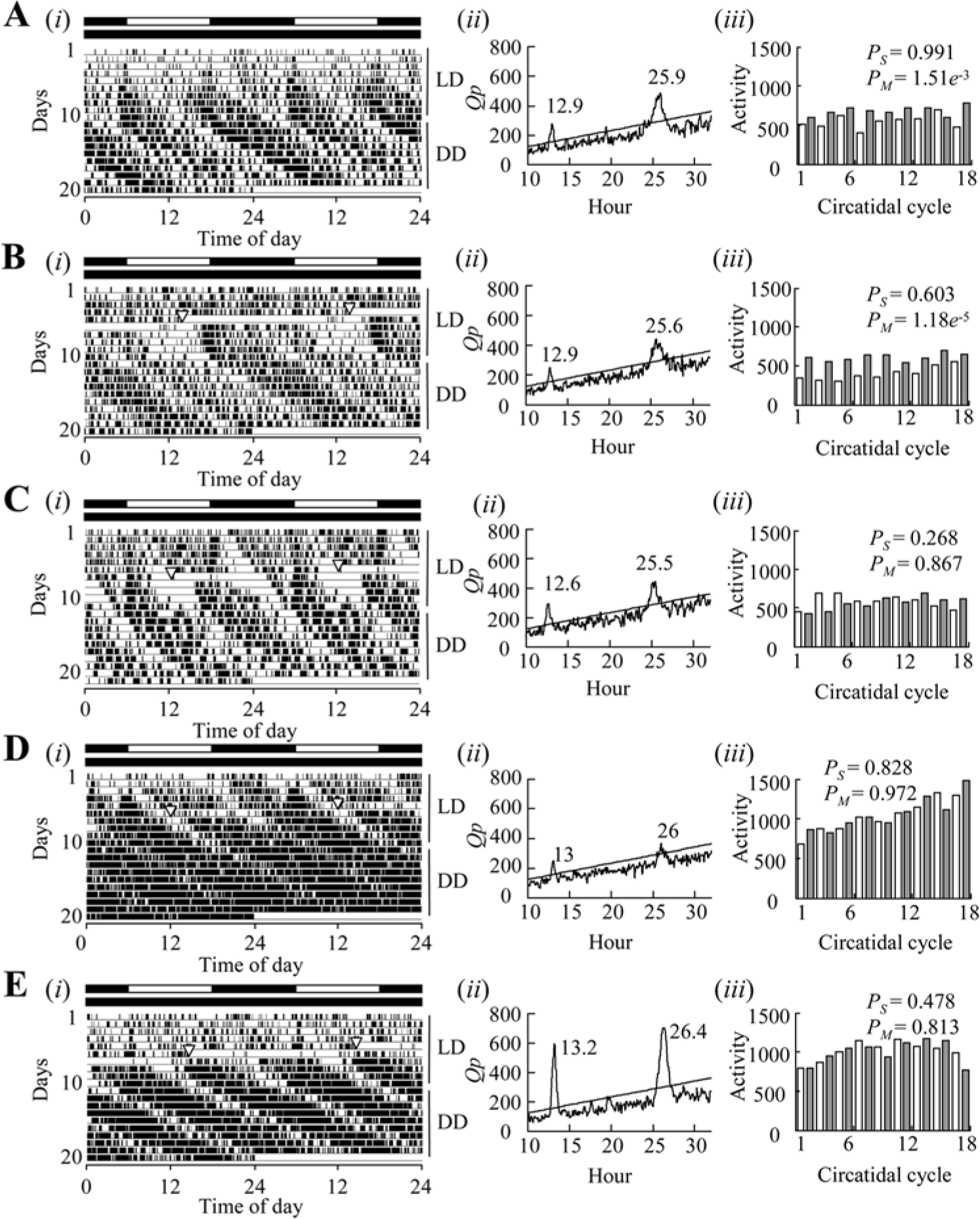
The representative locomotor activity rhythms of adult males of Apteronemobius asahinai. Intact (A), bilateral sham operation (B, C), and bilateral removal of the optic lobes (D, E). Locomotor activities were recorded under 12 h light and 12 h darkness cycles (LD) for 10 or 11 days and under constant darkness (DD) for a subsequent 10 days. After recording activity, crickets were sacrificed for PDF immunocytochemistry to check whether somata of PDF-IRNs had been completely removed (Fig. 4–6). (i) The activities presented by the double-plotted actograms. Black and white bars above the actograms indicate photo- and scotophases, respectively. Inverted triangles on the actogram show the time of the operation. (ii) The chi-square periodograms of activities during DD. The oblique line in the periodogram indicates the significance level of α = 0.05, and a peak value above the line is designated as significant. (iii) The histograms of activity levels counted for each circatidal period in DD. White and gray histograms represent the odd- and even-numbered circatidal cycles, respectively. PS and PM indicate the p values of ANCOVA for the slope of the regression line and the means, respectively.
Under LD cycles, the activity levels in the photophase were significantly lower than those in the scotophase in the 4 groups of intact, unilateral sham-operated, bilateral sham-operated, and unilateral optic lobe-removed crickets, but the activity amounts were not significantly different between scotophase and photophase in bilateral optic lobe-removed crickets (Wilcoxon signed rank test, p > 0.05; Suppl. Fig. S1). Under DD, approximately 80% of the crickets showed circatidal rhythms regardless of whether the optic lobes were removed. There was no significant difference in the proportion of crickets having circatidal rhythms among the 5 groups of intact, sham-operated, and optic lobe–removed crickets (Fig. 3A). The free-running periods were not significantly different among these 5 groups. Most of the intact crickets with a circatidal rhythm also showed a circadian rhythm (Fig. 3B). In the sham-operated groups, approximately one-half of the crickets showed a circadian rhythm. In the optic lobe–removed groups, only 3 crickets showed a circadian rhythm after unilateral removal of the optic lobe, and none showed a circadian rhythm after its bilateral removal. There was no significant difference in the proportion of crickets with a circadian rhythm between intact and sham-operated groups and between the bilateral sham and optic lobe–removed groups. But the proportions in the optic lobe–removed groups were significantly lower than in the intact group (Fig. 3B). This result suggests the surgery itself caused damage on circadian rhythmicity to some extent. However, bilateral removal of the optic lobes completely disrupted circadian rhythms, and the occurrence of circadian rhythms was much less frequent than that in the bilaterally sham-operated group.
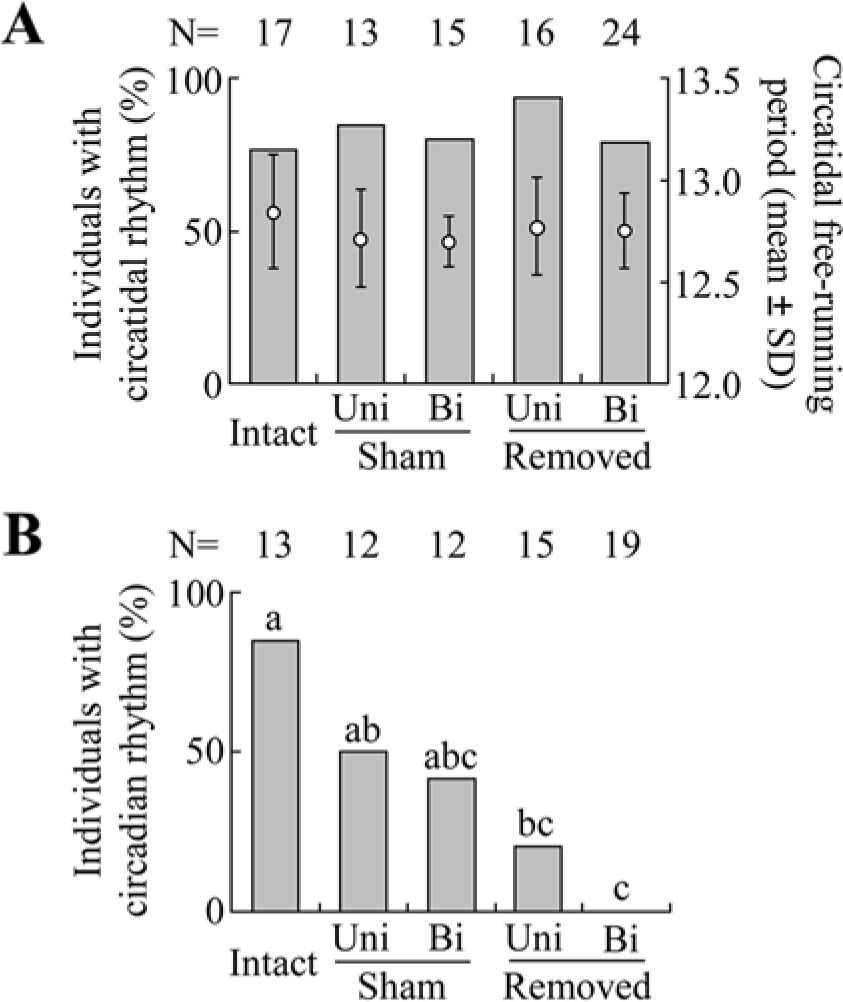
The effects of removal of the optic lobes on circatidal and circadian rhythms in male adults of Apteronemobius asahinai under constant darkness. Crickets were kept intact (Intact), had the margin of the compound eyes incised for sham operation (Sham), or had the optic lobes removed (Removed). The operations were performed unilaterally (Uni) or bilaterally (Bi). (A) The proportion of crickets with a circatidal rhythm and its free-running period determined by a chi-square periodogram. There was no significant difference among the 5 groups in proportions (p > 0.05, chi-square test) and circatidal periods (p > 0.05, one-way ANOVA). (B) The proportion of crickets with a circadian rhythm among those with a circatidal rhythm. The circadian rhythmicity was determined by ANCOVA for the activity levels of even and odd circatidal cycles. The proportions with the same letters are not significantly different (Tukey-type multiple comparisons for proportions, p < 0.05; Zar, 2010).
After recording activity, we observed PDF-IRNs of the brain. In intact crickets, 3 groups of somata were found to be PDF-immunoreactive in the optic lobes (Figs. 4A, 5A). A cluster of somata was found at the posterior dorsal (PDFLad) and posterior ventral (PDFLav) edge of the lamina. A third group of cells (PDFMe) was found at the anterior boundary between the medulla and lobula. This group contained large (≥5.0 µm) and small (<5.0 µm) somata (Table 1). From PDFLad and PDFLav, fibers appeared to project into the proximal region of the lamina. From PDFMe, fibers centrally projected along the posterior surface of the lobula to the central brain and peripherally extended to the anterior surface of the medulla. The peripheral fibers from PDFMe projected close to the PDFLad and PDFLav. In sham-operated crickets, no remarkable difference in the PDF-IRNs was observed from intact crickets (Fig. 5B).
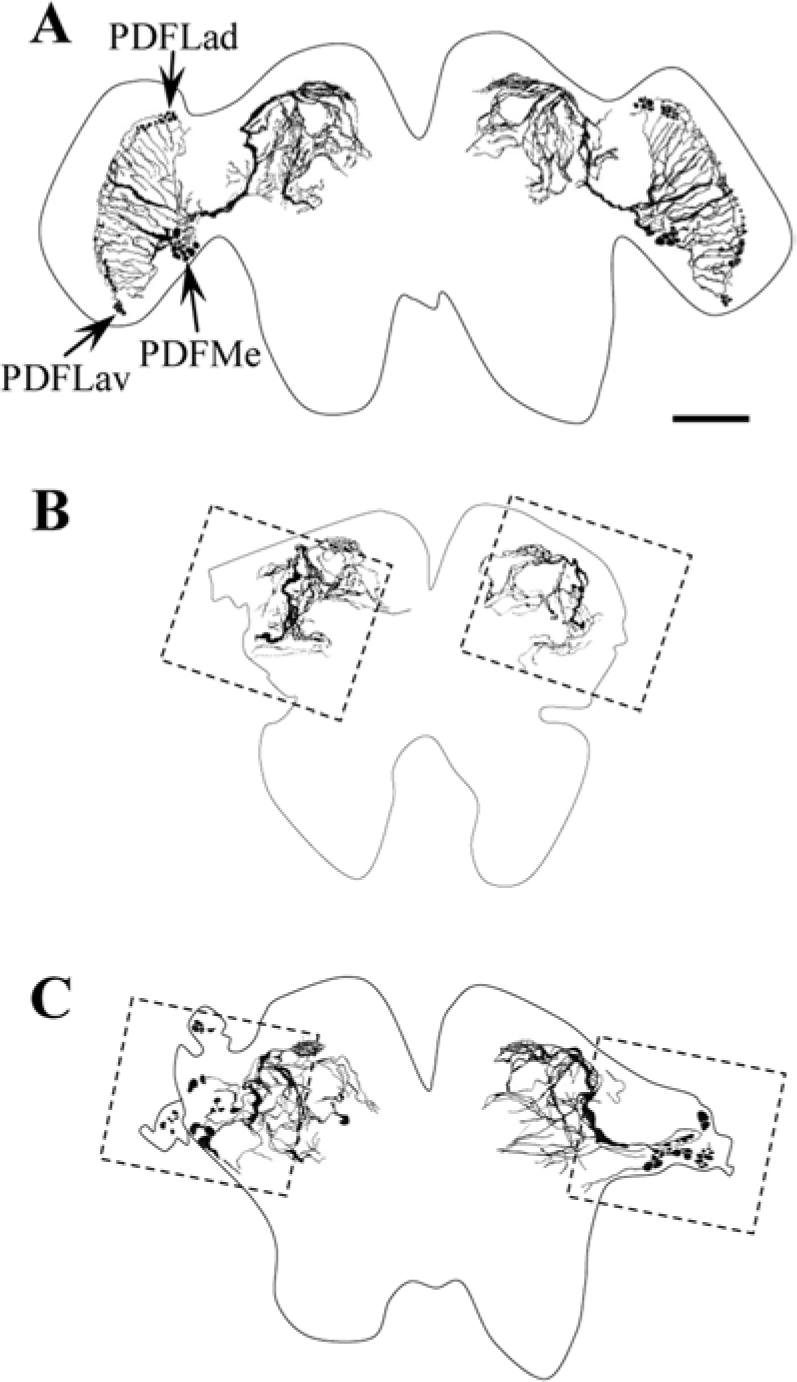
The tracings of pigment-dispersing factor immunoreactive neurons (PDF-IRNs) in the brains of Apteronemobius asahinai kept intact (A) or with bilateral optic lobes removed (B, C). The somata of the PDF-IRNs were completely removed in B but not in C. The tracings of B and C were obtained from the same individual in Figure 2D and 2E, respectively. The dotted squires in B and C indicate the region corresponding to Figure 5C and 5D, respectively. Scale bar is 50 µm.
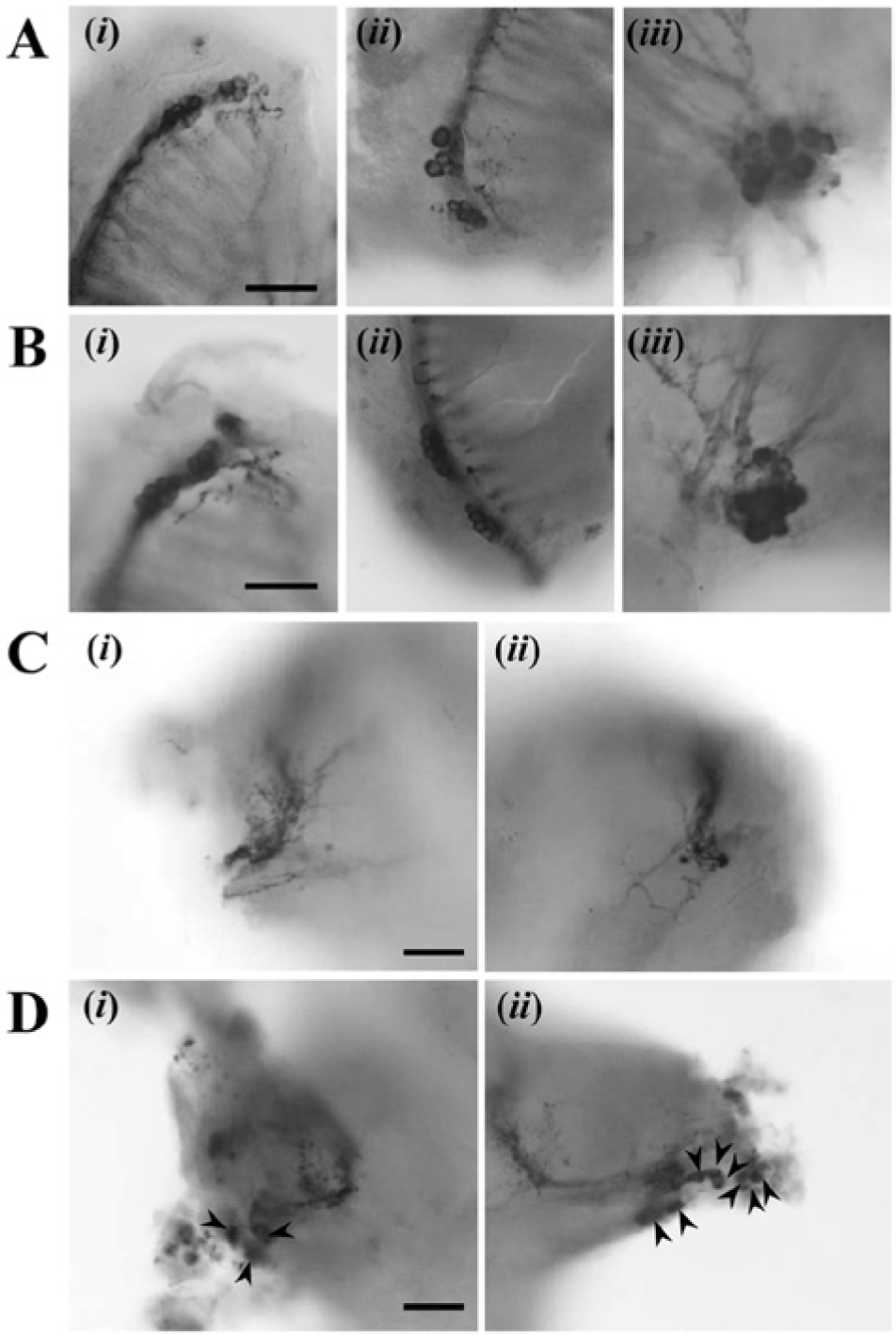
The photomicrographs of pigment-dispersing factor immunoreactive neurons in the brains of Apteronemobius asahinai kept intact (A), with the margin of the compound eyes incised for sham operation (B), or with the optic lobes removed (C, D). The somata in i, ii, and iii of A and B correspond to PDFLad, PDFLav, and PDFMe, respectively. In C and D, i and ii are enlarged images of the amputated parts in the left and right site of the brain (dotted squares in Fig. 4B, C), respectively. The arrowheads indicate the remnants of PDF-immunoreactive somata. Scale bars are 20 µm.
Diameter of pigment-dispersing factor (PDF) immunoreactive somata in the optic lobe of Apteronemobius asahinai.
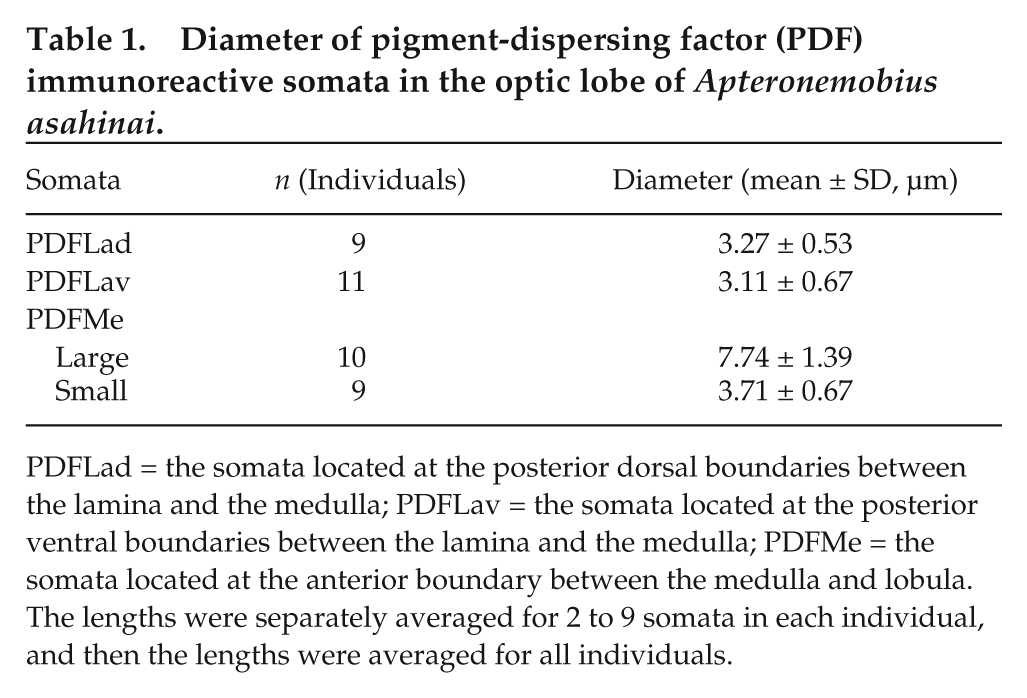
PDFLad = the somata located at the posterior dorsal boundaries between the lamina and the medulla; PDFLav = the somata located at the posterior ventral boundaries between the lamina and the medulla; PDFMe = the somata located at the anterior boundary between the medulla and lobula. The lengths were separately averaged for 2 to 9 somata in each individual, and then the lengths were averaged for all individuals.
In 23 of 24 crickets with the optic lobe bilaterally removed, the PDF-IRNs were stained successfully. PDFLad and PDFLav were completely removed in all crickets; however, PDFMe were completely removed in only 6 crickets (Fig. 4B, 5C). In the other 17 crickets, while the clusters were destroyed in all optic lobes, some somata were observed unilaterally or bilaterally (Fig. 4C, 5D). Because usually somata regions were injured after surgery, it was difficult to distinguish large and small types of the remnant PDF-IRNs somata. The degree of damage on the somata was variable. Some somata were found in an irregular place especially close to the amputated region, and some of them irregularly produced short fibers toward the central brain. The fiber projections in the central brain were similar to those in intact brains in all 23 crickets, and therefore fiber projections of PDF-IRNs seemed not to be affected by the presence of somata. The presence of somata of PDF-IRNs was not correlated with the occurrence of circatidal rhythms or with their free-running periods (Fig. 6). Regardless of the PDF-IRNs remnant, approximately 80% showed circatidal rhythms, and the free-running periods were 12.75 ± 0.20 h in the crickets without the remnant (n = 5) and 12.76 ± 0.15 h in those with the remnant (n = 14; Fig. 6).
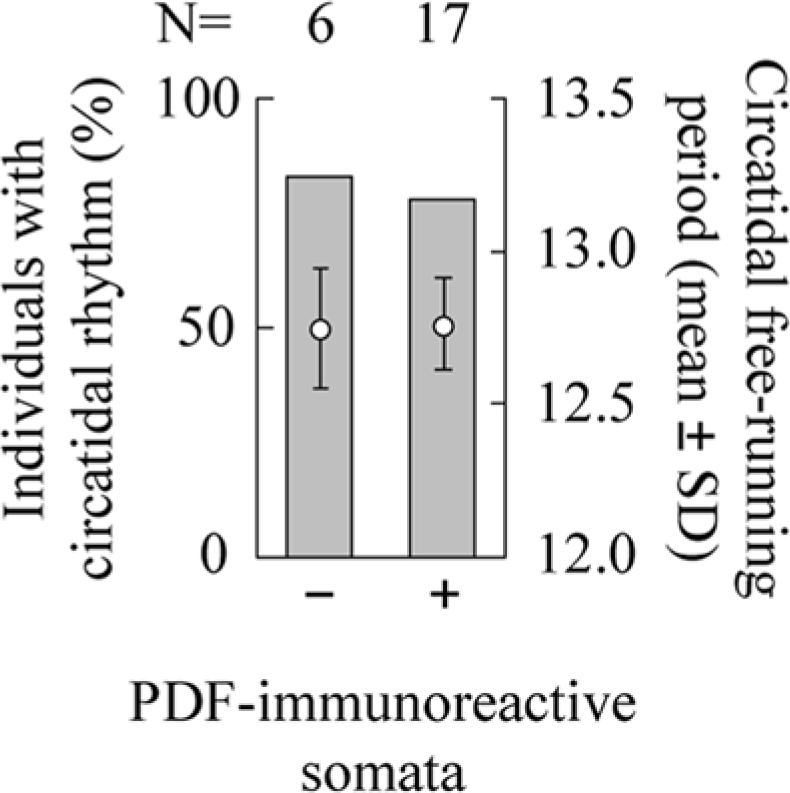
The relationship between the circatidal rhythm and PDF-immunoreactive somata. Occurrence of a circatidal rhythm and its free-running period under constant darkness in male adults of Apteronemobius asahinai receiving bilateral removal of the optic lobes without PDF-immunoreactive somata (−) and with PDF-immunoreactive somata (+) are shown. After recording activity, whether PDF immunoreactive somata had been completely removed was checked. Both of the proportions and the free-running periods were not significantly different (p > 0.05, Fisher exact test and t test, respectively).
Discussion
Although the circadian modulation was disrupted, the circatidal rhythm was maintained with no remarkable changes in its free-running period after removal of the optic lobes. These results clearly show that the principal circatidal clock is located in regions different from the optic lobe; therefore, the neural basis of the circatidal rhythm would be different from that of the circadian rhythm. When also considering the results of period RNAi (Takekata et al., 2012), it is noted that the physiological mechanism generating the circatidal rhythm is greatly different from that generating the circadian rhythm in A. asahinai.
Whether the circatidal rhythm is generated by the machinery common to the circadian clock is one of the important and interesting questions in chronobiology (Palmer, 1995; Wilcockson and Zhang, 2008; Naylor, 2010; Strauss and Dircksen, 2010). Concerning the clock mechanism generating the circatidal rhythm, several hypotheses have been established from the behavioral analysis of intertidal organisms, and the applicable hypothesis appears to be different among species (Wilcockson and Zhang, 2008; Naylor, 2010). The circalunidian clock hypothesis is that the circatidal rhythm is generated by 2 circalunidian clocks, each with a period close to a lunar day (24.8 h), coupled in antiphase (Palmer, 1995). This hypothesis is applicable to species such as the mud crabs Macrophthalmus hirtipes and Helice crassa, in which 2 circatidal activity peaks expressed each day show free-running periods different from each other (Palmer and Williams, 1986). In this hypothesis, it is postulated that the circalunidian clock is fundamentally the same as the circadian clock, because the free-running period of the circalunidian clock is very similar to that of the circadian clock (Palmer, 2000). This postulation is supported by observations of the plaice Pleuronectes platessa and the cumacean Dimorphostylis asiatica in which the circatidal rhythm is easily changed to the circadian rhythm under laboratory conditions (Gibson, 1973; Akiyama, 1997). In decapods, the importance of eyestalk for both the circatidal and circadian rhythms may imply the common machinery (Williams et al., 1979; Rodríguez-Sosa et al., 1994). In A. asahinai, however, the predominant clock generating the circatidal rhythm is located in a neural region different from that of the principal circadian clock, and a circadian clock gene period is not important (Takekata et al., 2012). Therefore, the clock system generating the circatidal rhythm in A. asahinai is not simply the same as the clock system generating the circadian rhythm as postulated by Palmer (2000).
The circatidal clock hypothesis would be most applicable for the circatidal rhythm of A. asahinai. The circatidal clock hypothesis is that the circatidal rhythm is generated by a circatidal clock independent of the circadian clock (Naylor, 1958, 2010) and can be applied to species such as the shore crab C. maenas, which shows simultaneous circatidal and circadian rhythms (Naylor, 1958). In A. asahinai, the phase response curve of the circatidal rhythm strongly suggests the existence of a circatidal clock with a period of 12.4 h (Satoh et al., 2009). Considering the results of period RNAi (Takekata et al., 2012) and optic-lobe removal in the present study, the most probable explanation for A. asahinai is that the circatidal rhythm is generated by the circatidal clock, which consists of a molecular and neural basis different from that of the circadian clock.
The bilateral removal of the optic lobes disrupted circadian modulation of activities in A. asahinai, suggesting that the circadian clock is located in the optic lobe. This interpretation is consistent with the results from studies on other crickets (Loher, 1972; Sokolove and Loher, 1975; Tomioka and Chiba, 1992; Abe et al., 1997; Shiga et al., 1999), although some different features were found. Unilateral removal of the optic lobe also disrupted the behavioral circadian rhythmicity of A. asahinai, in contrast to other crickets in which removal showed no significant effect (Loher, 1972; Sokolove and Loher, 1975; Ushirogawa et al., 1997; Shiga et al., 1999; Okamoto et al., 2001). In addition, sham operations also reduced the occurrence of circadian rhythms, although it is not significantly different from intact crickets. These facts suggest that circadian modulation in locomotor activity is fragile and disappears more easily in A. asahinai than in other crickets showing only circadian rhythms in behavioral activities. The surgical operation itself might affect the circadian modulation to some extent, causing no significant differences observed between bilateral sham-operated and bilateral optic lobe–removed groups. Although we cannot completely exclude the possibility that disruption of the circadian rhythm is partly caused by damage of the surgical manipulation, it may be probable to interpret that the complete absence of circadian rhythms in crickets without the optic lobe shows the importance of the optic lobe in circadian rhythmicity. With respect to the culture experiment showing that the optic lobe contains the circadian pacemaker in G. bimaculatus (Tomioka and Chiba, 1992), it is reasonable to conclude that the optic lobe in A. asahinai also contains the circadian pacemaker.
Among PDFLad, PDFLav, and PDFMe, a few PDFMe remained after surgery in some crickets, but the presence or absence of PDFMe somata was unrelated to the occurrence of the circadian and circatidal rhythms in A. asahinai, suggesting that neither large nor small PDF-IRNs are functionally indispensable in driving the circatidal and circadian rhythm. In surgical examinations of the Madeira cockroach Leucophaea maderae, PDFMe is suggested to be the principal circadian clock neuron (Stengl and Homberg, 1994; Reischig and Stengl, 2003). In contrast, PDF-IRNs do not appear to be the circadian clock neurons in the cricket G. bimaculatus; therefore, the principal circadian clock could be located in the distal region of the medulla, as indicated by partial removal of the optic lobes (Okamoto et al., 2001). The present result—the complete absence of the circadian modulation regardless of the remnants of PDFMe cells—appears to agree with the results in G. bimaculatus. Concerning the circatidal rhythm, the persistence of the rhythm after complete elimination of the somata of PDF-IRNs shows the presence of other principal circatidal clock neuron(s). Further, considering that the absence of the PDF-IRN somata is not related to the circatidal free-running period, PDF-IRNs would not be the circatidal clock neurons, at least not predominant ones. While the participation of PDF immunoreactive fibers in the midbrain could not be still denied, PDF-IRNs do not appear to be important in the circatidal clock system.
The present study shows that the principal circatidal clock is located in a region different from the optic lobe, and the next step should be to reveal its locus. In the house cricket Acheta domestica, the pars intercerebralis was reported to be the locus of the circadian clock (Cymborowski and Dutkowski, 1969, 1970; Cymborowski, 1981). Removal of the pars intercerebralis might reveal clues regarding the locus of the circatidal clock in A. asahinai. In addition, recording electrical activities from a target brain region could be a means by which to localize the circatidal clock, as with the circadian clock (Colwell and Page, 1990; Tomioka and Chiba, 1992). After the components of the circatidal clock system are revealed, we will be able to compare the clock machineries to discuss the homology between the circadian and circatidal rhythms.
Footnotes
Acknowledgements
The authors thank Takashi Haramura for his help with collecting insects, as well as Kenji Tomioka for providing the antiserum.
Conflict of Interest Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.