
Abstract
Phase resetting by light is an important feature of circadian rhythms, and the current Drosophila model focuses on light-mediated degradation of the clock protein TIMELESS (TIM). PERIOD (PER) is the binding partner of TIM and a major repressor of the molecular clock, but direct evidence of PER in phase resetting is lacking. Because light sensitivity of the perS short period mutant strain is strongly enhanced compared with wild-type strains, we assayed the importance of PER degradation for light-induced phase shifting. The perS protein (PERS) is markedly less stable than wild-type PER, in tissue culture and in flies, and PERS as well as PER is stabilized by TIM in both systems. Consistent with this finding, light-induced TIM degradation appears to trigger PER degradation. Moreover, TIM degradation is similar in the clock neurons of both strains, suggesting that it is not strongly affected by PERS and does not dictate the difference in the light response. In contrast, there is a dramatic quantitative difference between PER and PERS degradation in these neurons, indicating that PER degradation dictates the enhanced amplitude of the light-induced phase response. The data indicate that TIM inhibits PER degradation and that PER degradation follows light-mediated TIM degradation within circadian neurons; PER degradation then dictates qualitative as well as quantitative features of light-mediated phase-resetting.
Circadian clocks control the physiology and behavior of most eukaryotes and even some prokaryotes. This endogenous clock keeps ticking with a period of about 24 h even without temporal input from the environment. The circadian mechanism of Drosophila includes a transcriptional feedback loop, which controls the cycling expression of multiple clock genes including period (per) and timeless (tim). The transcription of per and tim is activated by a heterodimer of 2 basic-helix-loop-helix (bHLH) transcription factors, CLOCK (CLK) and CYCLE (CYC). The mRNA levels of per and tim peak in the early night, and their proteins (PER and TIM), accumulate to peak levels in the middle to late night (Hardin et al., 1990; Marrus et al., 1996; Sehgal et al., 1995) in fly heads. PER and TIM then enter the nucleus, where they inhibit the transcriptional activity of CLK:CYC and as a consequence their own transcription (Darlington et al., 1998; Nawathean and Rosbash, 2004; Saez and Young, 1996). A similar feedback loop operates in mammals.
In Drosophila, the perS mutant strain is 1 of 3 original per alleles isolated by Konopka and Benzer in 1971. Its free running locomotor activity rhythm is striking, about 5 h shorter than wild-type (wt) strains. The perS mutation is a serine to arginine missense mutation at amino acid 589 (Bargiello et al., 1987; Yu et al., 1987). The region surrounding position 589 is intimately related to the DOUBLETIME (DBT) kinase. This is an important PER modifying enzyme and a casein kinase I family member, which affects circadian period (Kloss et al., 1998; Preuss et al., 2004; Price et al., 1998). The recent study by Edery and colleagues indicates that the normal phosphorylation of the 589 serine by DBT inhibits the rate of phosphorylation at other key sites and therefore delays the daily PER degradation program. The S589N mutation in the perS protein (PERS) prevents this phosphorylation and therefore causes premature phosphorylation at the other key sites (Chiu et al., 2011). All features of perS cycling, including those of PERS abundance and phosphorylation, are advanced in these flies (Edery et al., 1994). Also significantly advanced is perS mRNA cycling, which suggests premature PERS transcription (Hardin et al., 1990; Marrus et al., 1996). It is unknown whether all these advances are due to a primary effect of the mutation on PER half-life.
Another complication is that PER and TIM form a heterodimer, in artificial systems as well as in flies (Gekakis et al., 1995; Zeng et al., 1996). Although the function of the PER-TIM complex is uncertain, considerable data indicate that TIM and the PER-TIM complex are necessary for the nuclear entry of PER (Hunter-Ensor et al., 1996; Saez and Young, 1996; Vosshall et al., 1994). This is despite the fact that other experiments indicate that PER and TIM enter the nucleus independently (Meyer et al., 2006; Nawathean and Rosbash, 2004).
Given the observation that PER is more unstable without TIM in flies (Price et al., 1995), it is believed that PER is stabilized within the PER-TIM complex. Perhaps it protects PER from phosphorylation by DBT (Cyran et al., 2005; Kloss et al., 2001), or perhaps the complex is more accessible to stabilizing phosphates like protein phosphatase 1 (PP1) (Fang et al., 2007; Sathyanarayanan et al., 2004). It is also unknown whether these possibilities might help explain the difference between PERS and PER.
Possibly relevant to the PER-TIM complex is the sensitivity of Drosophila circadian rhythms to light. A short light pulse in the early night causes a phase delay of fly locomotor activity rhythms, whereas a late night light pulse causes a phase advance (Saunders et al., 1994). These are general features of the phase-response curve (PRC) of many circadian systems including those of mammals. The current model in flies is based on light-mediated degradation of TIM: Light triggers a conformational change of the Drosophila blue light photoreceptor CRYPTOCHROME (CRY) within circadian neurons. This causes formation of a CRY-TIM complex and then rapid TIM degradation (Busza et al., 2004; Ceriani et al., 1999; Emery et al., 1998; Hunter-Ensor et al., 1996; Stanewsky et al., 1998). In the early night, a light pulse therefore causes premature TIM degradation, which delays the TIM accumulation profile and leads to a phase delay. A late night light pulse accelerates the TIM degradation profile and therefore causes a phase advance. However, this model does not explain why there is little or no phase response to a light pulse during the middle of the night between the delay and the advance zone regions of the PRC. This is the so-called “crossover region” of a PRC. Relevant to this issue is the PRC of perS flies, which was found long ago to have relatively little light-insensitivity, that is, little or no crossover region (Saunders et al., 1994). Another manifestation of this enhanced light sensitivity of perS is that these flies lose rhythmicity at an intensity of constant light too low to affect wt flies (Konopka, 1979; Saunders et al., 1994).
In this study, we addressed the effects of PERS and used it as a tool to further understand the mechanism of light-mediated phase shifting and oscillator synchronization. The degradation rate of PERS is faster than that of PER, and PERS as well as PER is stabilized by TIM. This is true not only in flies, as previously shown, but also in tissue culture. A similar distinction takes place after a light pulse in clock neurons; that is, PERS disappeared faster than PER. This explains the previously described, dramatically different PRCs of wt and perS flies. The data indicate that PER degradation dictates qualitative as well as quantitative features of light-mediated phase-resetting and oscillator entrainment and should be added to the classical cell-autonomous phase-resetting model.
Materials and Methods
Drosophila Stocks and Plasmids
Drosophila melanogaster were reared on standard cornmeal/agar medium supplemented with yeast. The wild-type (Canton-S) and perS flies were described by Konopka and Benzer in 1971. The flies were entrained in 12:12 light-dark (LD) cycles at 25 °C. The per01, elav-GAL4, and UAS-per16 and UAS-per24 line were described previously (Blanchardon et al., 2001; Yang and Sehgal, 2001). The cry-GAL4(13) (Emery et al., 2000) and tim-GAL4 (Kaneko and Hall, 2000) were crossed into the per01 background. To generate UAS-perS transgenic flies, the coding region of per with a site mutation (G1766A) and a V5 tag sequence at the 3¢ end was inserted into pUAST between EcoRI and XbaI site. The pUAST-perS-V5 plasmid was injected into y w embryos (BestGene, Inc). Two transgenes, UAS-perS7M and 9M, were used in the experiments. For the heat-shock experiment in cell culture, the coding region of per and perS with a V5 tag sequence, and the coding region of tim with a HBH tag sequence, were inserted into pCaSpeR-HS between EcoRI and XbaI sites.
RNA and Protein Analysis
Total protein was extracted from fly heads in RBS buffer (Yu et al., 2006). The protein extracts were resolved on 3% to 8% Tris-Acetate gel (Invitrogen) and transferred on nitrocellulose membrane using the iBlot Dry Blotting (Invitrogen, Carlsbad, CA). Antibodies against PER and TIM were used as described (Dembinska et al., 1997; Tang et al., 2010).
S2 Cell Transfection and In Vitro Degradation Assay
Drosophila Schneider 2 (S2) cells were maintained in insect tissue culture medium (HyClone, South Logan, UT) with 10% fetal bovine serum and 1% anti-anti (Antibiotics-Antimycotics, GIBCO, Carlsbad, CA) at 25 °C. Transfection was performed when the cell confluence reached 50% to 70% with Cellfectin II (Invitrogen) by standard protocol (Nawathean et al., 2005). Then 300 ng of pCaSpeR-HS-per, perS, or tim plasmid was transfected into S2 cells in 6-well plates about 24 h before heat shock. To activate the expression of per and tim, the plates were put in a 37 °C water bath for 30 min. Then the plates were put back into a 25 °C incubator for 2 h. Next, the cells were treated with 100 µg/mL cycloheximide. The cells were lysed by RBS buffer at different time points. For immunoprecipitation, the cell lysates were incubated with anti-V5 agarose overnight. The agarose was then washed for 3 times and denatured by SDS buffer at 100 °C.
Locomotor Activity Analysis
Locomotor activity of individual male flies (aged 2-5 days) was measured with Trikinetics Activity Monitors (Waltham, MA) for at least 4 days under 12:12 LD conditions followed by 6 days of constant darkness at 25 °C. For light pulse experiments, a single pulse (~15 mW/cm2; 10 min) was delivered to the flies during the last night of an LD cycle, and the flies were then maintained in constant darkness. The resultant phase shift was measured as described previously (Tang et al., 2010). Briefly, the group activity was generated and analyzed with MATLAB (MathWorks, Natick, MA) (Levine et al., 2002). The phase difference between experimental and control groups after several cycles in constant dark condition was then measured and averaged.
Fly Brain Immunocytochemistry
Immunostaining was performed as described (Yoshii et al., 2008). Briefly, fly heads were removed and fixed in PBS (pH 7.2) with 4% paraformaldehyde and 0.008% Triton X-100 for 1 h at 4 °C. Fixed heads were washed in PBS with 0.5% Triton X-100 and dissected in PBS. The brains were blocked in 5% goat serum (Jackson Immunoresearch, West Grove, PA) and subsequently incubated with primary antibodies at 4°C overnight. For TIM staining, a polyclonal rat anti-TIM was diluted 1:200 in blocking solution. For PER staining, a polyclonal rabbit anti-PER was precleaned by per01 fly head extract and used at a 1:50 dilution. For PDF staining, a mouse anti-PDF antibody (Developmental Studies Hybridoma Bank, University of Iowa, Iowa city, IA) was used at a 1:20 dilution. After washing with 0.5% PBST 3 times, the brains were incubated with Alexa Fluor 622 conjugated anti-rabbit (PER) or anti-rat (TIM) and Alexa Fluor 488 conjugated anti-mouse (Molecular Probes, Carlsbad, CA) at 1:200 dilution. The brains were washed 3 more times before being mounted in Vectashield Mounting Medium (Vector Laboratories, Burlingame, CA) and viewed in 1.1-µm sections sequentially at 20x on a Leica SP2 confocal microscope. To compare the PER or TIM signals from different genotypes and time points, the laser intensity of the confocal was set at the same level during each experiment. The PER or TIM signals within PDF cells were quantified with ImageJ.
Results
PERS Degradation Is More Rapid Than PER and Is Inhibited by TIM
Previous work has shown that the mRNA levels of the cycling genes per and tim accumulate earlier in perS than in wt flies (Hardin et al., 1990; Marrus et al., 1996). We observed essentially identical early accumulation of tim pre-mRNA (data not shown), consistent with the expectation that the phase of clock gene mRNA cycling is predominantly transcriptional (Rodriguez et al., 2013). Because PER is a critical repressor of CLK-mediated transcription (Allada et al., 2001; Takahashi, 2004), it is possible that this earlier transcription rise is due to a more rapid turnover of PERS relative to PER in the late night (Marrus et al., 1996) (Fig. 1A). However, expression and turnover occur concurrently, making it difficult to measure definitively the relative turnover rates in vivo. To address this issue, we expressed both PER and PERS in S2 cells under the control of a heat-shock promoter. Following transcriptional activation by heat shock, the translation inhibitor cycloheximide (CHX) was added, and PER and PERS levels were then determined over a time course of 6 h with anti-PER Western blots.
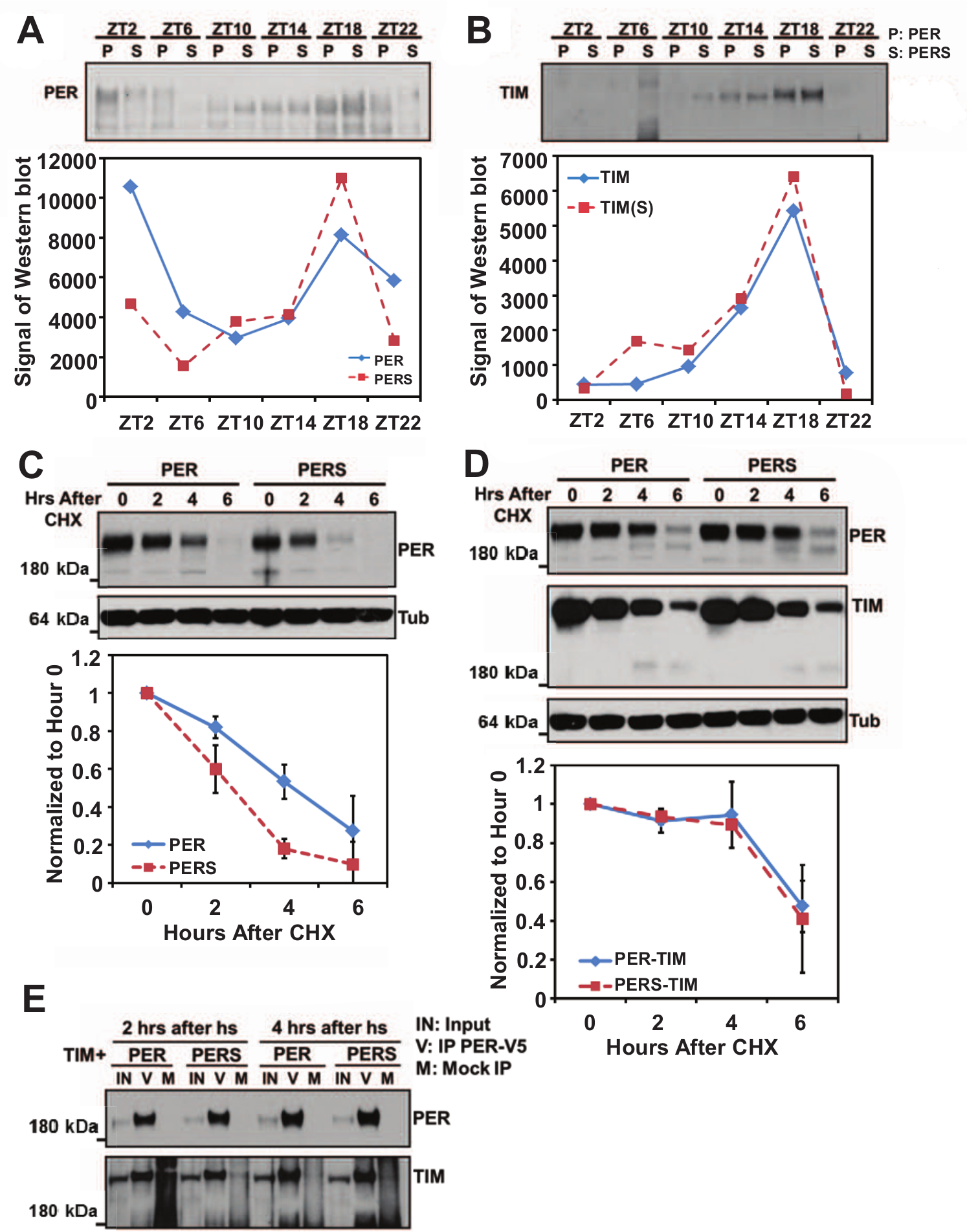
The faster degradation of PERS is inhibited by TIM. Heads of wt and perS flies were collected at different time points throughout the day. Extracted proteins were resolved on SDS-PAGE and detected by anti-PER (A) or anti-TIM (B) Western blots. Band intensities were quantified by ImageJ. PER degradation rate was assayed in vitro, in S2 cells. Per-V5 and perS-V5 (C) under the control of a heat shock promoter were induced using a 37
PERS decreased dramatically within the first 2 h after addition of CHX. The half-life of PERS is about 4 h shorter than that of PER (Fig. 1C), indicating that the stability of PERS is substantially lower than that of PER in this system. In flies, TIM forms a very stable heterodimeric complex with PER (Zeng et al., 1996) and also appears to stabilize PER (Ko et al., 2002; Nawathean and Rosbash, 2004; Price et al., 1995). Consistent with these observations, the difference in apparent degradation rate between PER and PERS is only detectable after about ZT18, a time in the night when TIM starts to disappear during a normal circadian cycle (Meyer et al., 2006) (Fig. 1, A and B). Although there may be some difference in the degradation rate of TIM between the wt and perS flies, TIM levels decrease quite fast after ZT18 in both genotypes.
To determine whether TIM can affect the degradation of PERS in the S2 cell system, we co-expressed a heat shock-inducible TIM with PER or PERS, and the degradation rates were assayed by Western blot as described above (Fig. 1D). The presence of TIM substantially extended the half-lives of PERS as well as PER. TIM appeared to affect the 2 proteins similarly, as neither one decreased significantly until 6 h after the addition of CHX.
We also verified that TIM interacts directly with PER/PERS in S2 cells. TIM and V5 tagged PER or PERS were expressed, PER/PERS was immunoprecipitated (IP) by anti-V5, and TIM was detected by Western blotting with an anti-TIM antibody (Fig. 1E). The association of PER/PERS and TIM occurred rapidly, within 2 h of expression, and there was no noticeable difference between PER-TIM and PERS-TIM complex formation (Fig. 1E).
Neuronal PERS Overexpression Causes a Short Period in per01 Flies
Previous work has shown that the PER can cycle independently of rhythmic transcription (Cheng and Hardin, 1998; Frisch et al., 1994). Moreover, overexpressing PER in clock neurons via the GAL4/UAS system in per01 null flies results in rhythmic flies with an approximately 24-h or even longer period (Busza et al., 2007; Grima et al., 2004; Kaneko et al., 2000) (Table 1). To assay whether overexpression of PERS via the same system in per01 flies give rise to a fast clock, we overexpressed PERS in both cry-expressing cells and the larger set of tim-expressing cells (which include all cry-expressing cells and many other cells). Both drivers rescue the arrhythmic per01 pheno-type and with a short period (Table 1).
Activity rhythm phenotypes produced by overexpression of per and perS in per01 background.
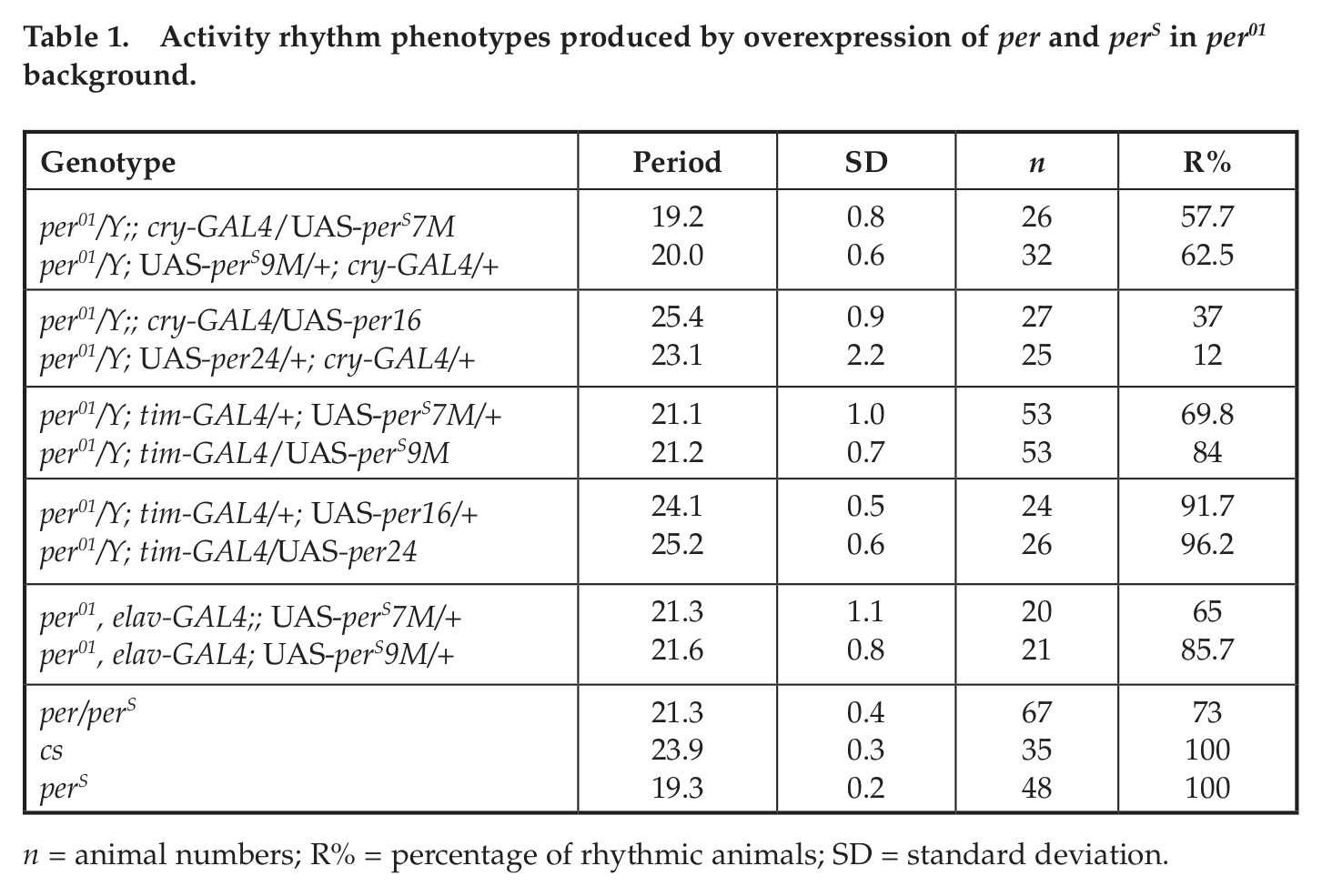
n = animal numbers; R% = percentage of rhythmic animals; SD = standard deviation.
Because these drivers have circadian promoters, PER overexpression may influence their activity. We circumvented this issue by also using the pan-neural driver elav-GAL4 (Robinow and White, 1988). It has no reported circadian transcriptional activity and in combination with UAS-per has been reported to give rise to approximately 24-h periods (Yang and Sehgal, 2001) (Table 1). We found that combining elav-GAL4 with either of 2 UAS-perS lines rescues per01 and gives rise to a short period (Table 1). This indicates the PERS alone dictates the short period.
Rapid Turnover of PERS in Clock Neurons Following a Light Pulse at ZT18
Light-induced phase shifting is linked to clock protein degradation. The current paradigm begins with a light-induced conformational change of CRY. This triggers CRY-mediated degradation of TIM, and the rhythm phase is then either delayed or advanced. The phase changes after light pulses at different circadian times are represented by a phase-response curve (PRC). It is characterized in wt flies by a Type 1 PRC, which has modest delays and advances as well as a substantial crossover region (little or no phase shift) in the middle of the night between the delays and advance (Fig. 2).
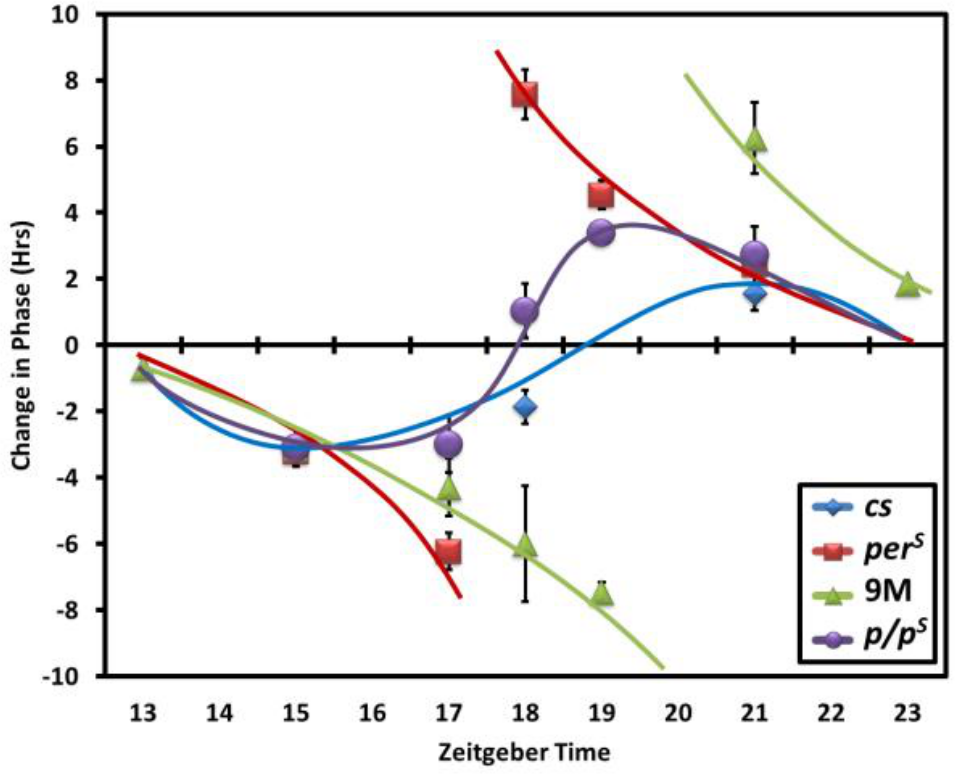
PERS causes a Type 0 light pulse PRC. A 10-min light pulse was delivered to the entrained flies at the indicated times during the night. The phase shift caused by the light pulse is shown as spots. The blue trend line indicates a Type 1 curve for wt flies, which has a gradual shift between the delay and advance zones. The perS flies have a Type 0 curve (red), which suddenly switches from the delay zone to the advance zone. The per01, elav::perS9M flies also have a Type 0 curve (green). The per/perS heterozygous female flies have a Type 1 curve (purple). The PRCs of wt and perS flies are based on our experiment results and published data (Bao et al., 2001). The tested time points and numbers of each genotypes: cs: ZT15 (n = 37), ZT18 (n = 32), ZT21 (n = 45); perS: ZT15 (n = 33), ZT17 (n = 24), ZT18 (n = 45), ZT19 (n = 63), ZT21 (n = 48); per/perS: ZT15 (n = 31), ZT17 (n = 42), ZT18 (n = 34), ZT19 (n = 39), ZT21 (n = 38); per01, elav::perS9M: ZT13 (n = 37), ZT15 (n = 47), ZT17 (n = 37), ZT18 (n = 48), ZT19 (n = 39), ZT21 (n = 46), ZT23 (n = 51). The phase delay of each genotype at ZT15: cs = −3.18 h; perS = −3.23 h; per/perS = −3.09 h; elav::perS9M = −3.09 h. The error bars indicate the standard deviation between multiple experiments.
Some strains like perS have a discontinuity PRC (also called Type 0 PRC), with more dramatic delays and advances as well as the absence of this distinct crossover region in the middle of the night; these PRCs have an abrupt shift from prominent delays to prominent advances (Konopka, 1979; Saunders et al., 1994) (Fig. 2). A Type 0 PRC is also characteristic of the short period doubletimeS strain, which like perS undergoes more rapid PER degradation (Bao et al., 2001). Interestingly, per01 flies rescued by elav-driven UAS-perS (per01, elav::perS9M) also have a Type 0 PRC (Fig. 2), indicating that it is likely due to the shorter half-life of PERS relative to PER rather than any transcriptional features of the gene. In per01, elav::perS9M flies, the delay-advance crossover point is also about 2 h delayed compared with perS flies, which presumably reflects the approximately 2 h longer period of per01, elav::perS9M flies compared with perS flies. Although different periods in perS and wt flies might affect the time of crossover in their PRCs, this experiment was performed to determine what type of PRC (0 v. 1) they displayed. The data suggest that the PRC is influenced by PER degradation as well as CRY-mediated degradation of TIM.
The difference between the wt PRC and the perS PRC is most dramatic at ZT18 in the middle of the night (Fig. 2). As mentioned above, it is notable that ZT18 is the starting point of spontaneous TIM degradation as assayed biochemically in both wt and perS fly head extracts (Fig. 1B). Moreover, PERS is more unstable than PER without TIM (Fig. 1C), and a 10-min light pulse is known to cause rapid TIM degradation in circadian neurons (Hunter-Ensor et al., 1996), especially in the late night (Tang et al., 2010). This predicts that there might be a difference in PERS v. PER degradation within circadian neurons subsequent to a light pulse.
To address this possibility, we assayed the effect of a 10-min ZT18 light pulse on PER and PERS levels as well as TIM levels by immunohistochemistry within PDF-expressing neurons (Figs. 3 and 4). One hour later at ZT19, PER levels in wt flies were not noticeably reduced compared with the no-light pulse control; this was also the case even 4 h after the light pulse, at ZT22 (Fig. 3, A and B). In contrast, PERS levels were dramatically lower 1 h after the light pulse, even less than those detected at ZT1 under standard LD conditions (Fig. 3B). TIM levels were reduced similarly in both genotypes and to a lesser extent than PERS levels (Fig. 4A and 4B). The most obvious TIM degradation occurred in nuclei, where PER and/or TIM contributes to transcriptional repression (Darlington et al., 1998; Nawathean and Rosbash, 2004; Saez and Young, 1996). The dramatic difference in PER v. PERS disappearance after a ZT18 light pulse is almost certainly relevant to the huge phase advance of perS flies at this time relative to virtually no phase shift of wt flies.
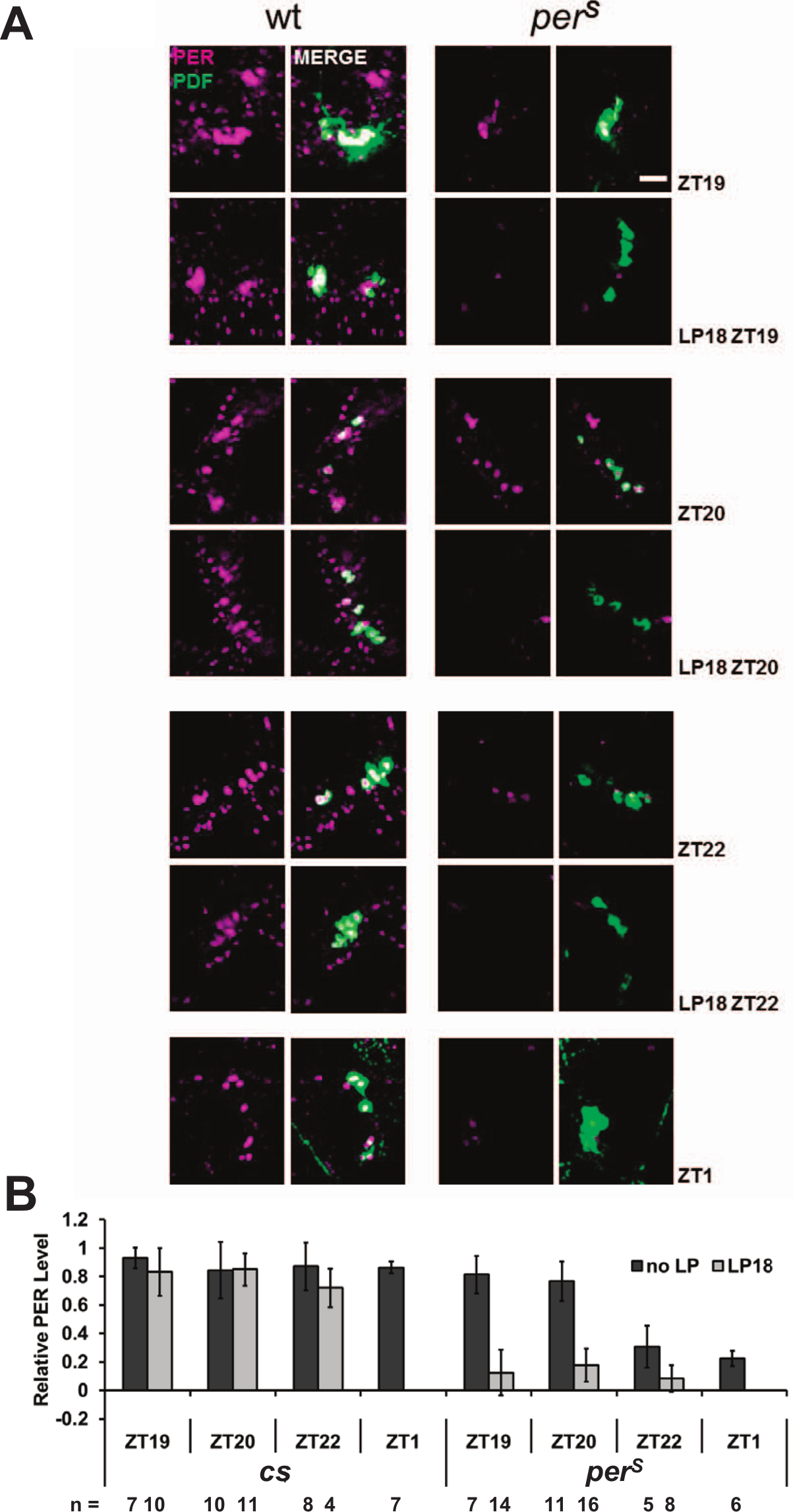
PERS degrades faster than PER in PDF cells after the light pulse at ZT18. A 10-min light pulse was given to the flies at ZT18. Fly brains were dissected and stained at ZT19, ZT20, and ZT22 to compare with the no-light pulse flies from same time points plus ZT1. (A) The PER staining pattern (magenta) in the PDF cells (green) of wt and perS flies with or without a light pulse at ZT18. Scale bar = 20 µm. (B) Quantification of PER signal in PDF cells from both genotypes per condition. All the results were normalized to a cs brain, which has the highest signal intensity at ZT19.
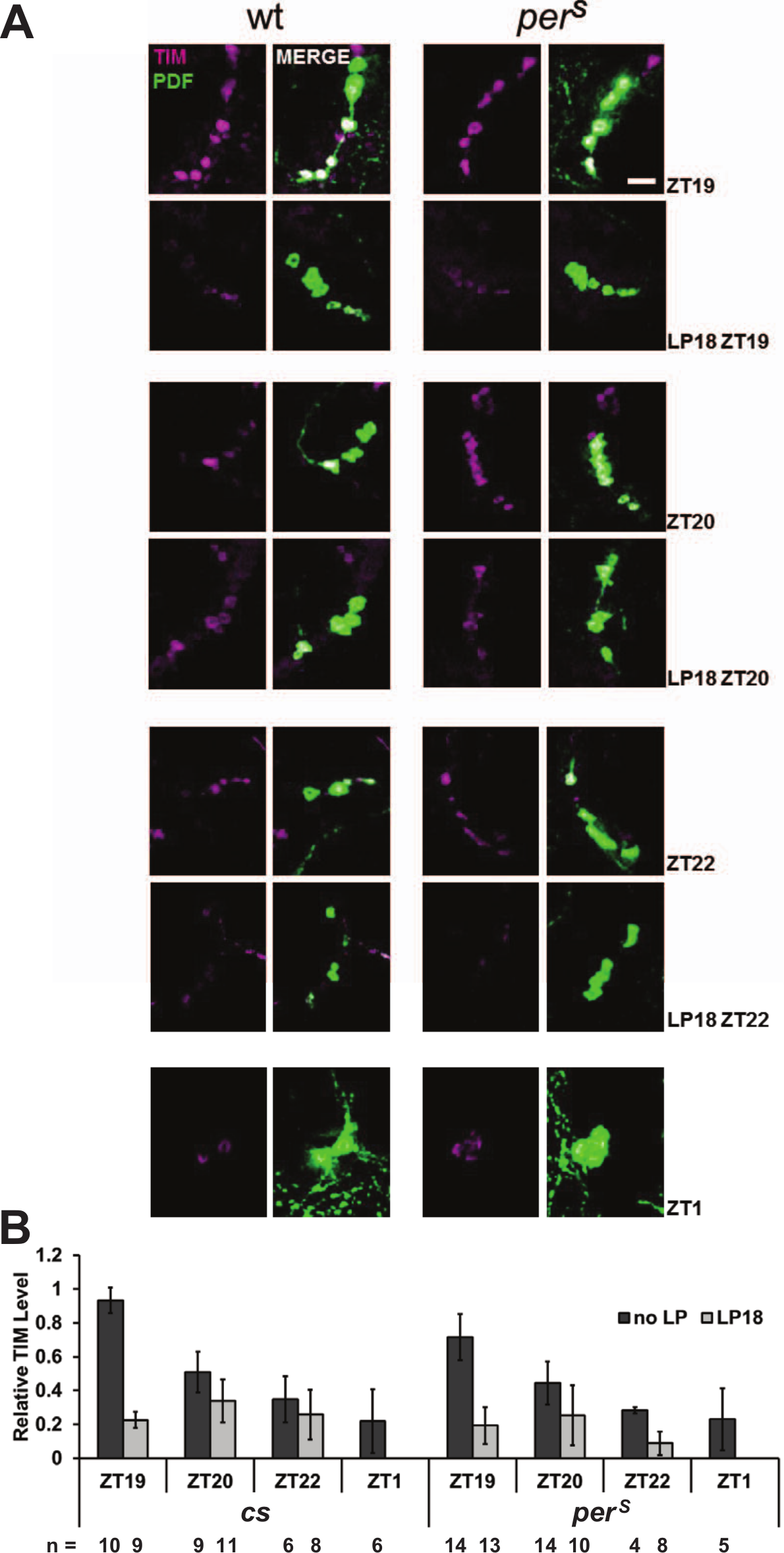
TIM degrades similarly in PDF cells of wt and perS flies after a light pulse at ZT18. Fly brains were collected in the same conditions described in Fig. 3. (A) The TIM staining pattern (magenta) in the PDF cells (green) of wt and perS flies with or without a light pulse at ZT18. Scale bar = 20 µm. (B) Quantification of TIM signal in PDF cells from both genotypes per condition. All the results were normalized to a cs brain, which has the highest signal intensity at ZT19.
In contrast to the middle of the night (ZT18; Fig. 2), the PRCs of wt and perS flies are quite similar to each other in the early night, at or before ZT15, as well as in the late night, at or after ZT21. A further staining experiment was therefore done after a 10-min light pulse at ZT21. In contrast to the ZT18 light pulse result, PER (Fig. 5A) as well as TIM (Fig. 5B) has similar levels in the 2 genotypes 1 h after this light pulse (Fig. 5, C and D). The result further supports the notion that PER degradation reflects or causes the magnitude of the phase shift and that residual PER levels after a light pulse dictate this magnitude.
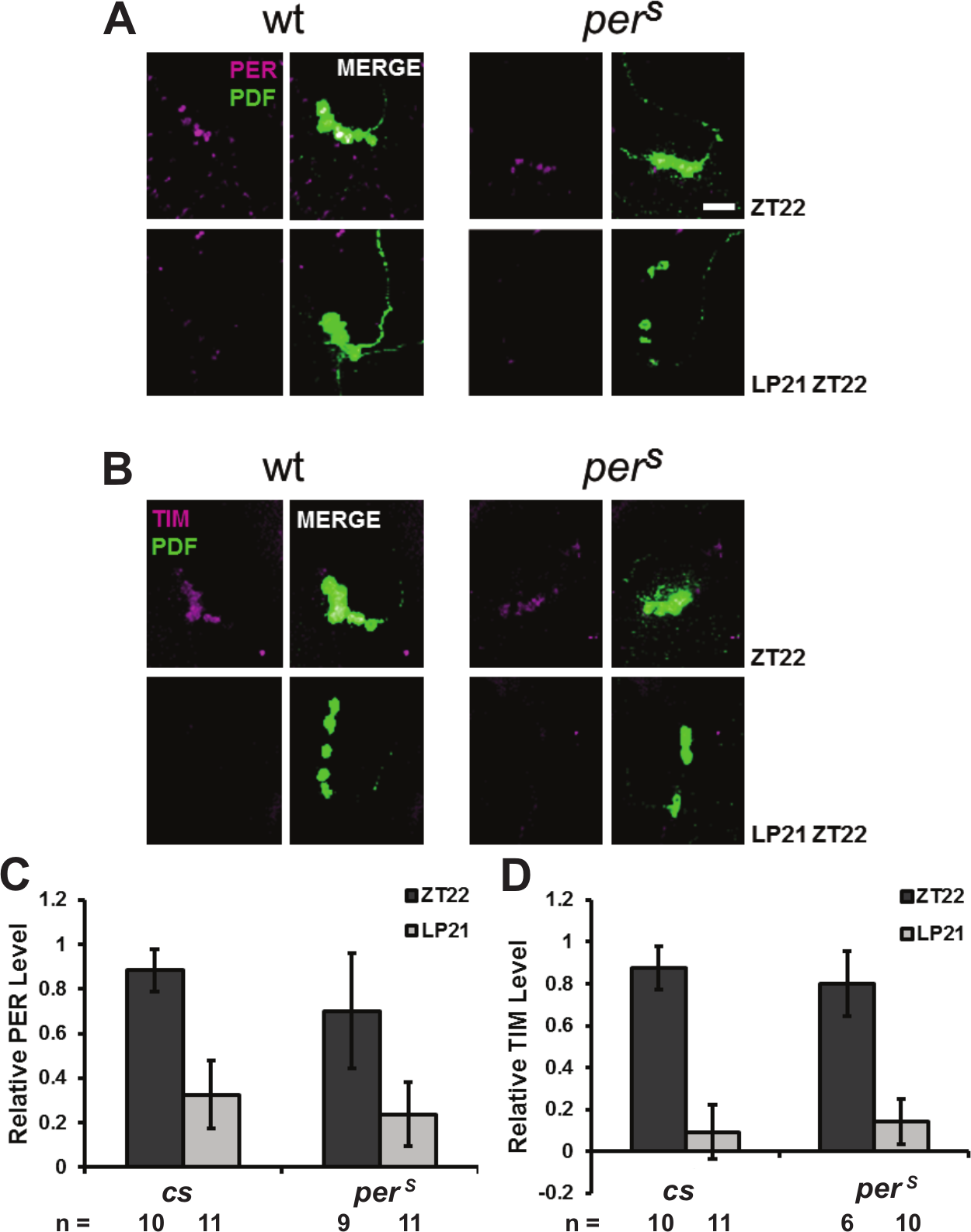
PER and PERS degradation is similar in PDF cells after a light pulse at ZT21. A 10-min light pulse was given to the flies at ZT21. Fly brains were dissected and stained at ZT22 to compare with the no-light pulse flies. (A) Anti-PER staining (magenta) in PDF cells (green). (B) Anti-TIM staining (magenta) of PDF cells. Scale bar = 20 µm. Quantification of PER (C) and TIM (D) signal in PDF cells of both genotypes per condition. All the results were normalized to a cs brain, which has the highest signal intensity at ZT22.
A prediction of this PER degradation view is that a light pulse will only trigger a decrease in PERS in per/perS heterozygous flies during the middle of the night period. This is because the substantial residual levels of wt PER after the light pulse should minimize the magnitude of the phase shift. The result shows that the per/perS heterozygous strain has a Type 1 PRC (Fig. 2) with almost no phase shift at ZT18. This indicates that the PRC effect of perS is recessive and suggests that residual PER levels after the light pulse indeed determine the magnitude of the phase shift. As the free-running period of this heterozygous strain is 21.3 h (Table 1), it suggests that the Type 0 PRC of perS flies is not attributable to the short period or the advanced phase of the molecular cycles. The data further suggest that PER multimers do not play an important role in light-induced PER turnover (see Discussion).
Discussion
We report here that PER is stabilized by TIM in tissue culture cells as well as in flies and that PER degradation occurs subsequent to TIM degradation. Importantly, the degradation rate of PERS is more pronounced than that of wild-type PER, both in tissue culture cells and following a light pulse in clock neurons. The results explain the previously described, dramatically different phase response curves of wt and perS flies. PER degradation rates therefore dictate qualitative as well as quantitative features of light-mediated phase-resetting and should be added to the standard cell-autonomous model of Drosophila phase shifting (Fig. 6).
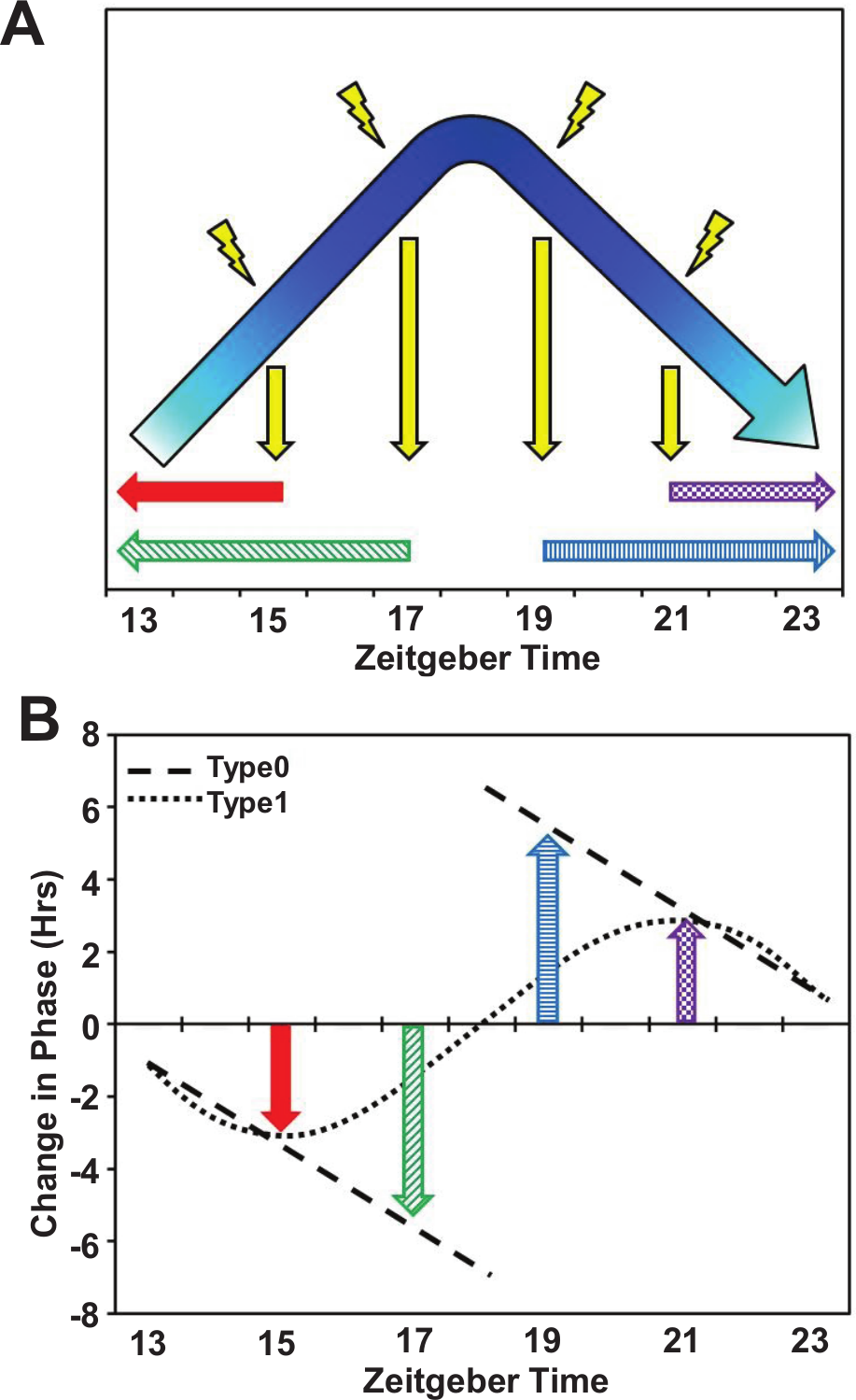
Current model to explain light-induced PRC. The current model to explain light pulse phase responses focuses on the degradation of TIM. The cycling of TIM occurs predominantly at night and includes 2 steps: climbing the slope (accumulation) and descending the slope on the other side (degradation). A light pulse causes the process to rapidly fall off the slope to trough levels. In the early night the process will restart from the beginning, which causes the delay, whereas in the late night the process will continue to the next cycle, thereby advancing the phase (A). Given this model, a plot of the change in phase, the PRC curve, will be Type 0 (B). To date, this type of PRC is only found in mutants that cause unstable PER (perS, doubletimeS [Bao et al., 2001] and per01, elav::perS), so we speculate that the process affected by the light pulse is PER degradation. Between ZT17 and ZT18, TIM is degraded by the light pulse, releasing PER for subsequent degradation. Since the degradation rate of PERS is faster than that of PER, at ZT17 PERS levels decrease to normal ZT12 levels and the clock is delayed. At ZT18, PERS is also degraded faster, which causes the accumulated PERS pool to decrease more quickly such that it resembles wt PER levels several hours later, which allows the clock to advance. For wt flies, PER cannot achieve the level of ZT12 or ZT24, so the flies only achieve a mild shift.
The earlier start of mRNA cycling and earlier disappearance of PERS protein had suggested that the half-life of PERS is shorter than that of wild-type PER and responsible for earlier transcription (Fig. 1A) (Marrus et al., 1996). Because transcription had not been directly measured, and because PER synthesis and degradation do not occur at completely distinct phases of the circadian cycle, we turned to tissue culture cells in which the start point of protein degradation can be precisely determined. PERS indeed turns over more rapidly in this system (Fig. 1C). This more rapid degradation then inspired an assay in flies using constitutive overexpression of PERS. It had been shown that constitutively expressed PER can endow per null flies with an approximately 24-h rhythm, indicating that cycling per transcription is not necessary for rhythmicity (Cheng and Hardin, 1998; Yang and Sehgal, 2001). Consistent with this notion, UAS-perS driven by the circadian promoters tim and cry linked to GAL4 has a short period phenotype in a per null background (Table 1). The driver elav-GAL4 is pan-neuronal and believed to promote constitutive gene expression with no relationship to the molecular clock (Robinow and White, 1988), and overexpression of UAS-perS with this driver also gives rise to a short period phenotype. We suspect that TIM levels and therefore the cycling of TIM are limiting for PER levels. This is because PERS becomes unstable as TIM levels decrease. So PERS cannot accumulate even when overexpressed. This makes more rapid PERS degradation an even more likely explanation for the short period phenotype of perS flies. Presumably other transcriptional targets of circadian feedback are sufficient for near normal rhythmicity.
The S2 cell experiments also showed that the well-described PER-TIM interaction (Zeng et al., 1996) can stabilize PERS as well as PER in this system (Fig. 1D). This result explains why the different degradation rates of PER and PERS are apparent in fly head extracts only after TIM begins to disappears at ZT18 (Fig. 1, A and B). As this is a time of maximal difference between the PRCs of wt and perS flies, we suspected that the very rapid TIM degradation subsequent to a light pulse might unmask the difference between PER and PERS degradation. Consistent with an important contribution of PER degradation to light-mediated phase shifting is the Type 0 PRC of the UAS-perS-rescued per01 flies, which is very similar to the perS PRC and completely different from a wt Type 1 PRC.
Although a light pulse causes a similar decrease in TIM staining in wt and perS central clock neurons at ZT18 (Fig. 4), PER staining was dramatically different in the 2 genotypes: It was stable in wt flies but rapidly decreased in perS flies after a light pulse (Fig. 3). We therefore conclude that the magnitude of PER degradation can determine quantitative as well as qualitative features of light-mediated phase shifting, including the Type 1 and Type 0 PRCs of wt and perS genotypes, respectively. It is notable that PERS is also more sensitive to low levels of constant light (Konopka et al., 1989), suggesting that the lengthened periods of perS flies under these conditions are due to decreased PERS levels.
A subsequent immunostaining experiment was done at ZT21, when a light pulse causes a similar modest phase advance in both wt and perS flies (Fig. 2). At this time, both PER and PERS staining decreases to a similar extent 1 h after the ZT21 light pulse (Fig. 5). Perhaps the accumulating PER modifications during the night endow the wt protein with a faster degradation rate subsequent to light-mediated TIM degradation at ZT21 than at ZT18. Alternatively, the major difference between PERS and PER degradation may occur earlier in the night. By ZT21, light-mediated degradation of residual PERS may be similar to light-mediated degradation of PER.
The cell-autonomous model of light-mediated phase shifting and its sole reliance on TIM degradation has changed only modestly since it was introduced in 1996 (Ceriani et al., 1999; Hunter-Ensor et al., 1996; Koh et al., 2006; Myers et al., 1996; Peschel et al., 2009). The early night is the accumulation phase of the PER-TIM cycle, so light-mediated TIM degradation at these times sets back this accumulation and therefore leads to phase delays. For example, the approximate 3-h phase delay caused by a light pulse at ZT15 sets back the circadian program to roughly ZT12. The late night is the decreasing or degradation phase of the PER-TIM cycle, so a light pulse at this time causes premature or more pronounced degradation and advances the cycle. The magnitude of the phase shifts caused by late night light pulses suggests that they advance the circadian program to the expected lights-on time of ZT0 or that they mimic premature dawn. Therefore, advances as well as delays appear to shift the clock to the 2 light transitions, lights-on (dawn) and lights-off (dusk), respectively. The fact that the early night phase delays and the late night phase advances of perS are quasi-normal despite the enhanced PERS degradation further suggests that ZT12 and ZT0 are important stopping points for the clock (Fig. 2 and Fig. 5).
It is notable that these 2 times are still relevant when perS flies experience light pulses in the middle of the night. Indeed, our results also offer a partial explanation for the crossover region of a normal Type 1 PRC, the time in the middle of the night when a light pulse causes little or no phase shift of wt flies. This region of the PRC is much more responsive in perS, indicating that the enhanced degradation rate of PERS can transition the PER-TIM program more dramatically, up to 6 h from the crossover point back to ZT12 and forward to ZT0. However, the crossover region of a wt Type 1 PRC, the transition point between delays and advances, still awaits a cogent explanation. It is likely that features of the fly brain circadian network are relevant (Stoleru et al., 2007); that is, the cell-autonomous model may be insufficient to explain this feature of Drosophila light-mediated phase shifting (Tang et al., 2010). For this reason, most of our analyses were restricted to the phase advance region. It may be more cell-autonomous than the phase delay region (Tang et al., 2010), making the immunohistochemistry focus on PDF cells more relevant to the resulting phase shifts.
A recent study on the crystal structure of PER showed that PER can form PER-PER homodimers via their PAS domains (Yildiz et al., 2005). Interrupting this dimerization with mutations within this domain dramatically inhibits these molecular interactions as well as behavioral rhythmicity (Landskron et al., 2009). This indicates that PER dimerization may contribute to the molecular clockworks. Unfortunately, the study did not address the region surrounding the perS serine 589. Although this mutation may therefore affect PER dimerization, we show here that per/perS heterozygous flies have a Type 1 PRC despite a free-running period of 21.3 h. This continuous PRC suggests that substantial PER levels must remain after the light pulse despite relevant PER-PER dimers in the heterozygous fly, that is, PER-PER, PERS-PERS, and PER-PERS.
It is still unclear what accounts for the enhanced degradation rates of PERS compared with PER, especially after a light pulse at ZT18 within PDF cells. Possibly relevant is the advance of the time-dependent phosphorylation events that occur on PERS relative to PER (Chiu et al., 2011). In this view, light simply unmasks PER and PERS by causing TIM degradation.
Footnotes
Acknowledgements
The authors thank Sean Bradley, Kate Abruzzi, Jerome Menet, Weifei Luo, and Fang Guo for thoughtful discussion. This work was supported by P01 NS44232 (M.R.).
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.