
Abstract
Most of what we know today about the molecular constituents of the insect circadian clock was discovered in the fruit fly Drosophila melanogaster. Various other holometabolous and some hemimetabolous insects have also been examined for the presence of circadian genes. In these insects, per, tim1, and cry2 are part of a core feedback loop system. The proteins inhibit their own expression, leading to circadian oscillations of mRNA and proteins. Although cockroaches are successfully employed circadian model organisms, their clock genes are mostly unknown. Thus, we cloned putative circadian genes in Rhyparobia maderae (synonym Leucophaea maderae), showing the presence of period (per), timeless 1 (tim1), and mammalian-type cryptochrome (cry2). The expression levels of per, tim1, and cry2 in R. maderae were examined in various tissues and photoperiods employing quantitative PCR. In brains and excised accessory medullae, expression levels of rmPer, rmTim1, and rmCry2 oscillated in a circadian manner with peaks in the first half of the night. Oscillations mostly continued in constant conditions. In Malpighian tubules, no significant oscillations were found. In animals raised in different photoperiods (LD 18:6, 12:12, 6:18), the peak levels of rmPer, rmTim1, and rmCry2 expression adjusted with respect to the beginning of the scotophase. The daily mean of expression levels was significantly lower in short-day versus long-day animals. We suggest that rmPer, rmTim1, and rmCry2 are part of the Madeira cockroach nuclear circadian clock, which can adjust to different photoperiods.
Cockroaches have been widely used in chronobiology due to their large size, adaptiveness, behavioral richness, and easy maintenance. The Madeira cockroach Rhyparobia maderae (synonym Leucophaea maderae) (for details on nomenclature, see Kevan [1980]), in particular, has been studied for circadian research since the 1960s, where its circadian pacemaker region controlling locomotor rhythms was located in the optic lobes of the brain (Nishiitsutsuji-Uwo and Pittendrigh, 1968a, 1968b). Even before the cellular localization of circadian pacemaker neurons, the general organization of the circadian system was elucidated in many elegant behavioral studies (Page et al., 1977; Block and Page, 1978; Page, 1982). Later, lesion and transplantation experiments located the circadian pacemaker center, which controls locomotor activity rhythms, to the accessory medulla (AMe), with the associated pigment-dispersing factor (PDF) containing neurons (Stengl and Homberg, 1994; Reischig and Stengl, 2003). During the past 20 years, the Madeira cockroach has become a model organism for neuropeptidergic and cellular circadian research (Petri and Stengl, 1997; Homberg et al., 2003; Reischig et al., 2004; Decker et al., 2007; Saifullah and Page, 2009; Wei et al., 2010; Söhler et al., 2011; Schulze et al., 2012). However, so far, no information on the molecular circadian clockwork of R. maderae is available.
The molecular circadian clockwork of insects has been most intensively studied in the fruit fly Drosophila melanogaster (Allada and Chung, 2010; Hardin, 2011; Peschel and Helfrich-Förster, 2011). Period (dPer) and timeless 1 (dTim1) are part of the “core feedback loop” of the circadian clockwork. Their expression is being activated by the transcription factors CLOCK and CYCLE (dCLK and dCYC), resulting in an accumulation of dPER and dTIM1 in the cytoplasm, where they are posttranslationally modified. With a characteristic delay, these clock proteins then translocate to the nucleus, where they inhibit their own transcription and close the feedback loop. Interlocked with the core feedback loop are the clock loop (Cyran et al., 2003; Glossop et al., 2003) and another feedback loop controlled by CLOCKWORK ORANGE (dCWO) (Kadener et al., 2007; Lim et al., 2007; Matsumoto et al., 2007). This nuclear circadian clock results in circadian oscillations of clock mRNA and protein levels. Furthermore, as a cell-autonomous light input pathway to the clockwork in the fruit fly, CRYPTOCHROME (dCRY) allows for light-dependent degradation of dTIM1. It controls light-dependent phase shifts of the circadian rhythm of clock gene expression (Stanewsky et al., 1998; Emery et al., 2000; Helfrich-Förster et al., 2001; Ivanchenko et al., 2001).
Various other holometabolous insects like Apis mellifera, Tribolium castaneum, and several lepidopteran (Antheraea pernyi, Bombyx mori, and Danaus plexippus) and dipteran species (Drosophila spp., Protophormia terraenovae, Anopheles gambiae, and others) were examined for circadian clock genes. Hemimetabolous insects were studied less intensively: Circadian genes of Pyrrhocoris apterus (Hodkova et al., 2003; Syrová et al., 2003), Riptortus pedestris (Ikeno et al., 2008), and Gryllus bimaculatus (Uryu and Tomioka, 2010) were characterized. Only in 2010, the first genome of a hemimetabolous insect, Acyrthosiphon pisum, was published, followed by an analysis of the circadian system (Cortés et al., 2010). In cockroaches, only per cDNA sequences of Blattella germanica (Lin et al., 2002) and Blattella bisignata and partial sequences of Periplaneta americana per (Reppert et al., 1994) and tim1 were published. However, no blattodean genome is available to the scientific community to date, in spite of their widespread employment as model organisms for basic research.
The basic principle of the core feedback loop described above for D. melanogaster is similar between species, and a generally high homology of the circadian genes involved in the molecular clockwork was found. However, genome sequencing projects revealed that in detail, there are striking differences between insect species (Rubin et al., 2006; Cortés et al., 2010). This is especially true for cry and tim. In insects, 2 different types of cry genes were found: Drosophila-type cry1 (cry1) and mammalian-type cry2 (cry2). The only cry of Drosophila is cry1, which encodes for a blue light photoreceptor. It is absent from the genomes of other insects such as A. mellifera and T. castaneum (Rubin et al., 2006). In contrast, cry2 is present in all nondrosophilid insect species examined so far. Some insects such as D. plexippus and A. pisum (with duplicated cry2 genes) express both cry1 and cry2 (Zhu et al., 2005; Cortés et al., 2010). It was shown in vitro that insect CRY2 works as a repressor of CLK-CYC–mediated transcriptional activation and appears to be light insensitive (Yuan et al., 2007). This is similar to mammals, where CRY has first been shown to be part of the negative feedback loop (Okamura et al., 1999; Van der Horst et al., 1999; Vitaterna et al., 1999). Not only the absence or presence of cry1 and cry2 differs between insect species, but also tim1 may be missing. A. mellifera does not have a tim1, and its function in the circadian core feedback loop seems to be “replaced” by CRY2. This is similar to the mammalian clock since the only mammalian “tim” is more similar to insect timeout (tim2) (Benna et al., 2000). Whereas TIM1 is part of the core feedback loop as described in Drosophila, TIM2 is important for DNA metabolism and chromosome integrity and only plays a marginal role in light entrainment (Benna et al., 2010).
Circadian Clocks and Photoperiodic Clocks
The effect of photoperiod on the circadian system as well as the role of the circadian system in the insect photoperiodic calendar is quite diverse in different species (Koštál, 2011; Saunders and Bertossa, 2011). It is not known whether the circadian pacemaker circuit constitutes also a photoperiodic clock (Shiga and Numata, 2007, 2009; Muguruma et al., 2010; Koštál, 2011). Photoperiodic clocks consist of a “photoperiodic timer” that allows for the distinction of short- and long-day photoperiods and a “photoperiodic counter” that initiates photoperiodic adaptations after about 2 weeks in the new photoperiod (Koštál, 2011). Furthermore, it is not known whether the circadian pacemaker of a circumtropical species such as the Madeira cockroach has the endogenous properties to adjust to different photoperiods.
Thus, here, we first cloned putative R. maderae core feedback loop homologs of per, cry2, and a short fragment of tim1. Using quantitative PCR, we examined whether these genes show circadian expression in different tissues and whether they are affected by different photoperiods.
Materials and Methods
Animal Cultures
R. maderae animal cultures were maintained at 27 °C in a 12:12 light:dark (LD) cycle. For short- and long-day experiments, animals were raised and maintained in 18:6 (long-day) and 6:18 (short-day) LD cycles. They were fed with dry dog food, potatoes, and apples; water was supplied ad libitum.
Molecular Cloning
Male R. maderae were dissected, and tissues from the supraesophageal ganglion (brain) and muscle plus Malpighian tubules were pooled, respectively. Total RNA was extracted using Trizol (Invitrogen, Carlsbad, CA). Poly A+ RNA was purified using Oligotex (Qiagen, Hilden, Germany). Adapter-ligated cDNA was produced using the Marathon RACE Kit (Clontech, Mountain View, CA). Degenerate primers for timeless 1 and cryptochrome 2 were designed by comparing known amino acid sequences of different insect species and choosing regions with high homology. For cloning of period, the nucleotide sequence of P. americana (accession no. U12772) was used to generate initial primers. PCR with degenerate primers was performed using Advantage Taq (Clontech) and the following parameters: 3 cycles at 94 °C (1 minute), 37 °C (1 minute), and 68 °C (3 minutes); 32 cycles at 94 °C (1 minute), 40 °C (1 minute), and 68 °C (3 minutes); and final elongation at 68 °C (3 minutes) followed by 4 °C. After an initial cDNA sequence fragment was obtained with degenerate primers, gene-specific primers (GSPs) were designed using Primer3 and Vector NTI (Invitrogen). The 5′ and 3′ RACE were then performed using the GSP and primer AP1 (Clontech), followed by nested PCR using a nested GSP and primer AP2 (Clontech), according to the manufacturer’s manual. For primer sequences, see Supplementary Tables S1 and S2. Bands of interest were excised, and DNA was extracted using the High Pure PCR Product Purification Kit (Roche, Basel, Switzerland). Fragments were TA ligated into pGEM-T or pGEM-T Easy vector systems and transformed into Escherichia coli K12 DH5α (Invitrogen). Positive clones were selected and cultured overnight in LB medium containing ampicillin. Plasmids were extracted using the PureYield Plasmid Miniprep System (Promega, Madison, WI). Sequencing from SP6, T7, or GSP was performed by Eurofins MWG Operon (Ebersberg, Germany) or JenaGen GmbH (Jena, Germany). The cDNA fragments were assembled using Vector NTI (Invitrogen).
Motif Prediction and Phylogenetic Analysis
We used SMART (Schultz et al., 1998; Letunic et al., 2012) and CDD (Marchler-Bauer et al., 2011) for motif prediction. NLStradamus (Nguyen Ba et al., 2009) was used to predict nuclear localization signals. Motifs with an amino acid identity higher than 40% were considered conserved. MEGA5 (Tamura et al., 2011) was used for phylogenetic analysis, and CLUSTALW as implemented in MEGA5 was used for alignments. Accession numbers of insect core feedback loop genes used for comparison are indicated (Suppl. Table S4). The neighbor-joining method was used with 1000 bootstrap replicates, and evolutionary distances were computed using Poisson correction. Gaps and missing sequences were eliminated for the analysis, resulting in 360 positions in PER and 421 in CRY2 being used.
Quantitative PCR
For LD 12:12 experiments, male R. maderae were dissected every 4 hours starting at ZT0. Tissue containing the AMe and associated neurons (n = 10 per pool), brains (without the AMe and associated neurons, photoreceptor cells, and neurohemal complex; n = 10 per pool), and Malpighian tubules (n = 10 per pool) were snap frozen in liquid nitrogen right after dissection. In the dark phase, dissections were performed under red LED light (λ > 610 nm). At each time point, 3 pools (3 × n = 10) were used.
For long- and short-day experiments, brains including the AMe and associated neurons, but without the neurohemal complex and photoreceptor cells, were used (n = 3). Samples were stored at −80 °C until use. At each time point, 3 pools (3 × n = 3) were analyzed, and the same samples were used for each gene. With RNeasy Mini columns (Qiagen), mRNA was extracted, and genomic DNA was digested on-column using RNase-free DNase (Qiagen). With random hexanucleotides (Roche), cDNA was primed and reverse transcribed using Sensiscript reverse transcriptase (Qiagen). RNA quality control was performed by determining the spectrum and 260/280 ratio using a NanoDrop ND-1000 UV-Vis spectrophotometer (Peqlab Biotechnologie GmbH, Erlangen, Germany). FAM-labeled TaqMan probes and primers were designed and synthesized by Applied Biosystems (Foster City, CA) (Suppl. Table S3). A short fragment of a R. maderae actin (Suppl. Fig. S1) was cloned and was used as a reference gene. TaqMan Universal PCR Master Mix, No AmpErase UNG (Applied Biosystems) containing the passive reference dye ROX was used with Applied Biosystems 7300 and 7500 qPCR cyclers according to the manufacturer’s manual. Triplica were analyzed for each cDNA sample. Relative mRNA expression ratios were calculated as described by Pfaffl (2001) and normalized to the arithmetic mean of the threshold cycle values across all time points of the respective gene of interest:
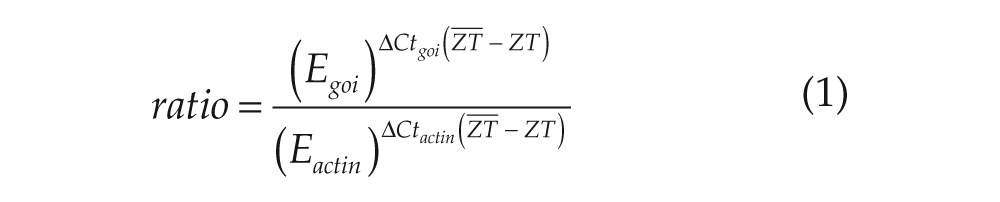
(E = efficiency, goi = gene of interest, ratio = relative expression ratio, ZT = sampled time point,
One-way ANOVA followed by a least significant difference (LSD) post hoc test (α = 0.05) was employed to determine significant differences between time points. The result of the post hoc test is indicated by characters above time points in the figures. Two time points with completely different characters above are significantly different. Cosinor analysis (Nelson et al., 1979) was performed by cosine wave curve fitting (ϕ = 24 hours) and zero amplitude tests (Suppl. Table S5).
To compare the daily mean of expression levels of the genes, the percentage of actin expression was calculated as follows:
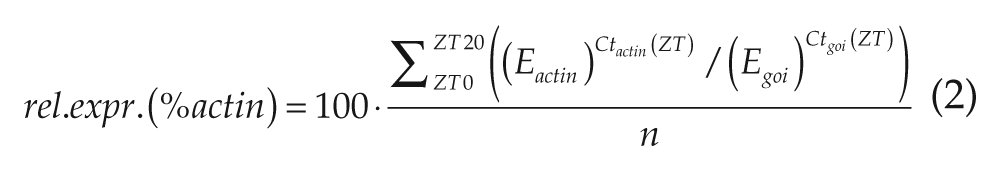
(E = efficiency, goi = gene of interest, n = number of time points, Ct = threshold cycle, and ZT = sampled time point).
One-way ANOVA (α = 0.05) was employed to determine significant differences. All values were computed and graphs generated using Matlab R 2012a (MathWorks, Natick, MA).
Results
To analyze molecular constituents of the nuclear circadian pacemaker of the Madeira cockroach R. maderae, RT-PCR and quantitative PCR were employed. First, using a degenerate primer PCR strategy and RACE, putative core genes of the circadian feedback loop of R. maderae, period (rmPer), cryptochrome 2 (rmCry2), and timeless 1 (rmTim1) were cloned. Then, it was examined whether these circadian genes show ZT-dependent rhythms of mRNA expression in different tissues: brain, AMe, and Malpighian tubules of animals kept in LD 12:12 were examined. In the same manner, we also assayed the expression levels of these genes in the brain of animals raised in different photoperiods: long-day (LD 18:6) and short-day (LD 6:18).
Period
The cDNA cloning and sequencing of rmPer resulted in a sequence of 4261 base pairs (accession no. JX235363) including part of the 3′ and 5′ untranslated regions (UTRs), an initial ATG (pos. 133-135) and stop codon (TAA, pos. 3961-3963). The protein encoded by the open reading frame (ORF) has a predicted size of 1276 amino acids and an estimated molecular weight of 138.7 kDa. The ORF contained c1 to c6 domains as described for drosophilid species (Colot et al., 1988) as well as the c7 domain defined by Lin et al. (2002) for B. germanica, which is also present in many nondipteran species and mammals (Fig. 1A and 1C). A SMART and CDD search revealed PAS-A, PAS-B (PER-ARNT-SIM), and PAC (PAS-associated C-terminal) domains, or PAS fold as it was redefined by Hefti et al. (2004). The PAC domain is part of the cytoplasmic localization domain (CLD) (Saez and Young, 1996). The PeriodC domain, including the c7 domain, was also found. Nuclear localization signal (NLS) prediction (NLStradamus) resulted in 2 predicted sites: One at the end of the c1 domain is well defined in D. melanogaster (Vosshall et al., 1994; Saez and Young, 1996). A second NLS (residues 1239-1242) was predicted near the C-terminal end with a similar motif in some insects with the c7 domain (Fig. 1A and 1C). In addition to the 2 NLS predicted by NLStradamus, a bipartite NLS previously described in D. melanogaster at the end of the c3 domain (Chang et al., 2003) is also conserved in rmPER (Fig. 1A and 1C). In cockroaches, there is no threonine-glycine repeat (TG repeat) as described for D. melanogaster, but serine-glycine repeated 4 times at a similar position, as known for other insects including B. germanica (Lin et al., 2002). Phylogenetic analysis places rmPER within the cockroach PER proteins (Fig. 1B). Schematic representation of rmPER compared with PER from different insect species (Fig. 1A) and the alignments of NLS from different species (Fig. 1C) show the high degree of conservation of different sequence motifs for this circadian gene.
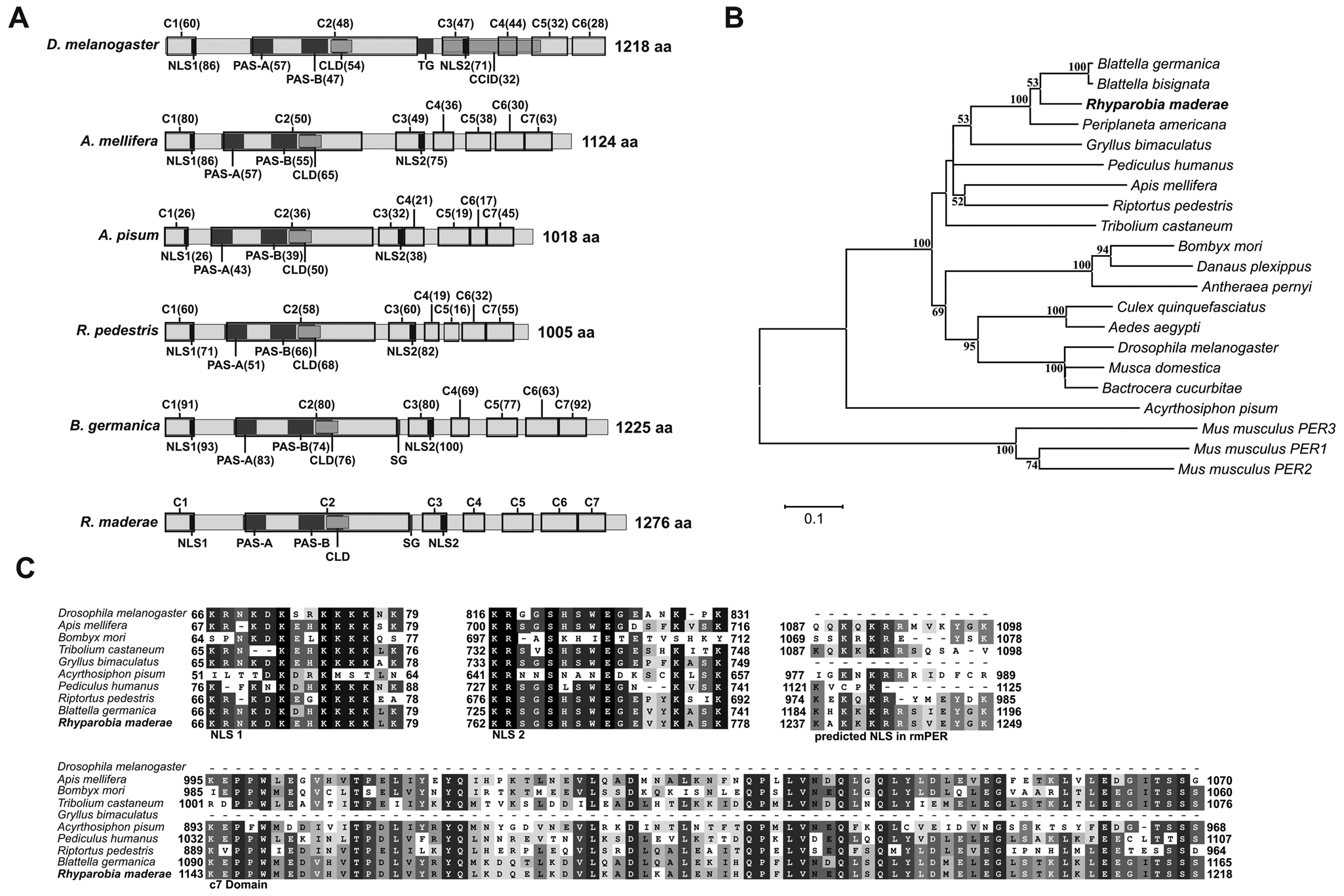
(A) Schematic representation of rmPER and various insect PER proteins. c1 to c6 domains (Colot et al., 1988) and the c7 domain (Lin et al., 2002) are indicated by black boxes. PAS-A and PAS-B domains are shown as identified by SMART (Letunic et al., 2012; Schultz et al., 1998) in rmPER. NLS, CLD, and CCID are indicated as identified in Drosophila melanogaster (Chang et al., 2003; Saez and Young, 1996; Vosshall et al., 1994). Numbers in brackets represent the percentage of amino acid identity with rmPER. Protein size is shown at the end of each diagram. (B) Phylogenetic relationship of various insect (including rmPER) and mouse PER proteins. The neighbor-joining algorithm as implemented in MEGA5 with Poisson-corrected distances was used. Numbers shown next to the branches represent the percentage of replicate trees in the boostrap test (1000 replicates), shown only if higher than 50%. (C) Alignments of 2 conserved PER NLS, a NLS predicted in the C-terminal region of rmPER and the c7 domain (Lin et al., 2002) sequences from different insect species. Numbers at the beginning and end of each alignment box indicate the position of the first and last residues with respect to the N-terminus. CCID = dCLK:CYC inhibition domain; SG = serine-glycine repeat; TG = threonine-glycine repeat.
Cryptochrome 2
Cloning and sequencing of rmCry2 cDNA yielded a sequence of 2221 base pairs (accession no. JX266618), with an initial ATG (117-119) and a stop codon (TGA, 1875-1877). The predicted protein size was 586 amino acids, with an estimated molecular weight of 67.3 kDa. This is the first cry reported in cockroaches. A CDD search predicted a DNA photolyase (amino acids ~10-175) and a flavin adenine dinucleotide (FAD) binding domain (amino acids ~217-492). Domains necessary for CLOCK:BMAL interaction (RD-2a, RD-1, and RD-2b) in mice are conserved in rmCRY2 as well as a CLD domain (Hirayama et al., 2003) and coiled-coil (C-C) region (Chaves et al., 2006) (Fig. 2A and 2C). Phylogenetic analysis places rmCRY2 within the insect CRY2 proteins next to another hemimetabolous insect, R. pedestris (Fig. 2B). Schematic representation of rmCRY2 compared with CRY2 from different insect species as well as with CRY1 from D. melanogaster (Fig. 2A) and the alignments of RD-1, RD-2b, and C-C domains from different species (Fig. 2C) show a high degree of conservation and show that the CRY examined is indeed an insect CRY2.
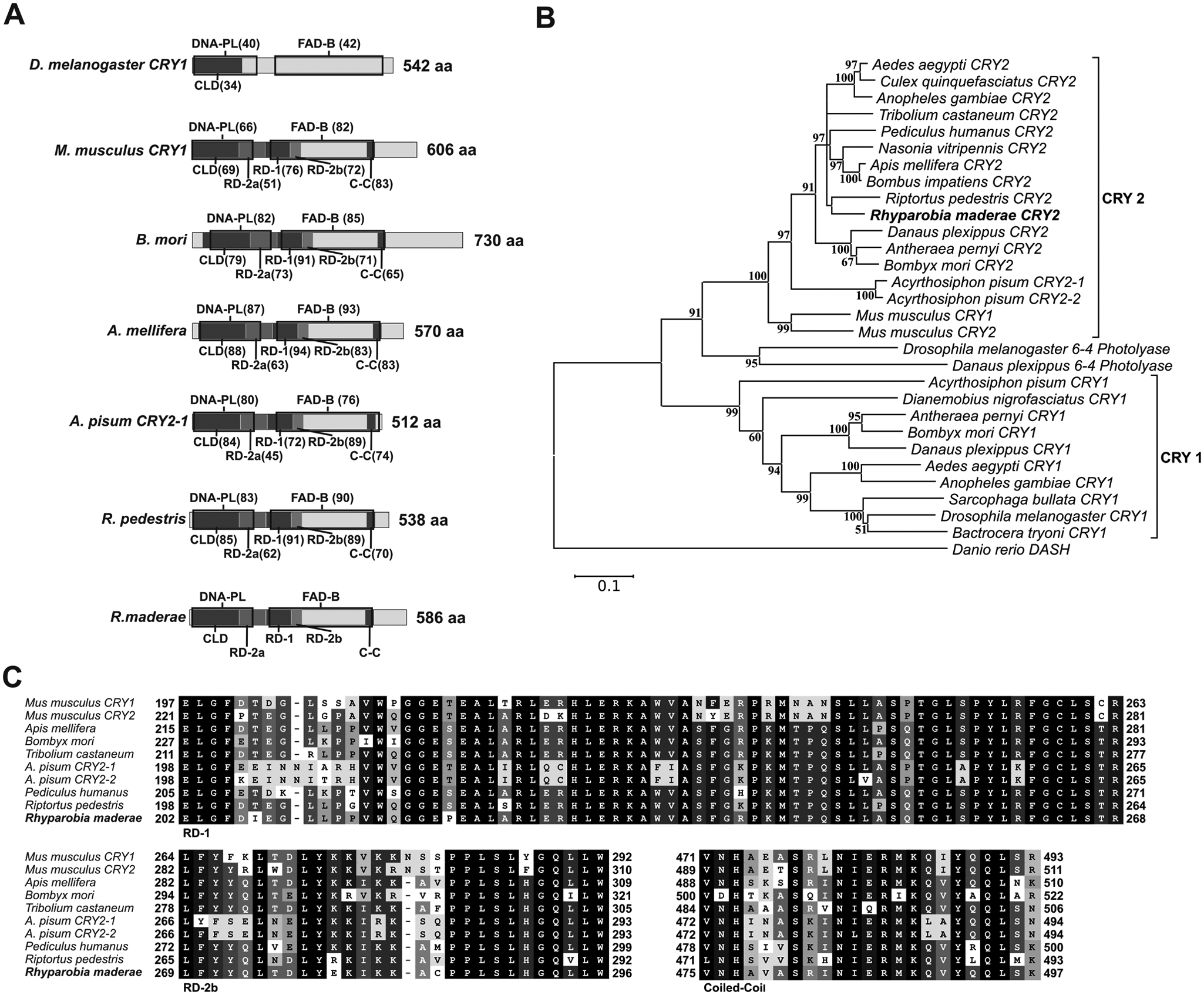
(A) Schematic representation of rmCRY2, various insect CRY2 proteins, and D. melanogaster CRY1. DNA photolyase conserved domain and FAD binding site are indicated by black boxes and were identified by CDD (Marchler-Bauer et al., 2011). CLD, RD-2a, RD-1, and RD-2b domains as identified in mouse CRY1 (Hirayama et al., 2003; Kume et al., 1999) and a C-C region (Chaves et al., 2006) are also shown. Numbers in brackets represent the percentage of amino acid identity with rmCRY2. Protein size is shown at the end of each diagram. (B) Phylogenetic relationship of different insect (including rmCRY2) and mouse cryptochromes and insect photolyases. For details, see the legend to Figure 1. (C) Alignment of RD-1, RD-2b, and C-C domain sequences from rmCRY2 and various insect CRY2 proteins. Numbers at the beginning and end of each alignment box indicate the position of the first and last residues with respect to the N-terminus. DNA-PL = DNA photolyase conserved domain; FAD-B = flavin adenine dinucleotide binding site.
Timeless 1
A clone with a 591-bp partial fragment (accession no. JX266619) of rmTim1 was obtained, sufficient for quantitative PCR. Due to the very low expression level of rmTim1 (Fig. 7A and 7B), a full-length sequence was not identified. The fragment obtained encodes for a 196-aa-long part of rmTIM1 (Suppl. Fig. S2A). If aligned with the 1421-aa-long isoform D of dTIM1, the obtained rmTIM1 fragment extends from aa 645 to aa 839. It starts 35 aa 3′ of the first PER dimerization domain (which includes the NLS) identified in dTIM1 (Saez and Young, 1996) and spans 93 aa into the second PER dimerization domain (Suppl. Fig. S2B).
Putative Core Feedback Loop Gene Expression in Different Tissues
First, it was examined whether there are daily oscillations in expression levels of the putative R. maderae core feedback loop genes rmPer, rmTim1, and rmCry2 in neuronal (Fig. 3) and nonneuronal tissues (Malpighian tubules) (Suppl. Fig. S3) in LD 12:12 and the first day in constant darkness (DD) (Fig. 4).
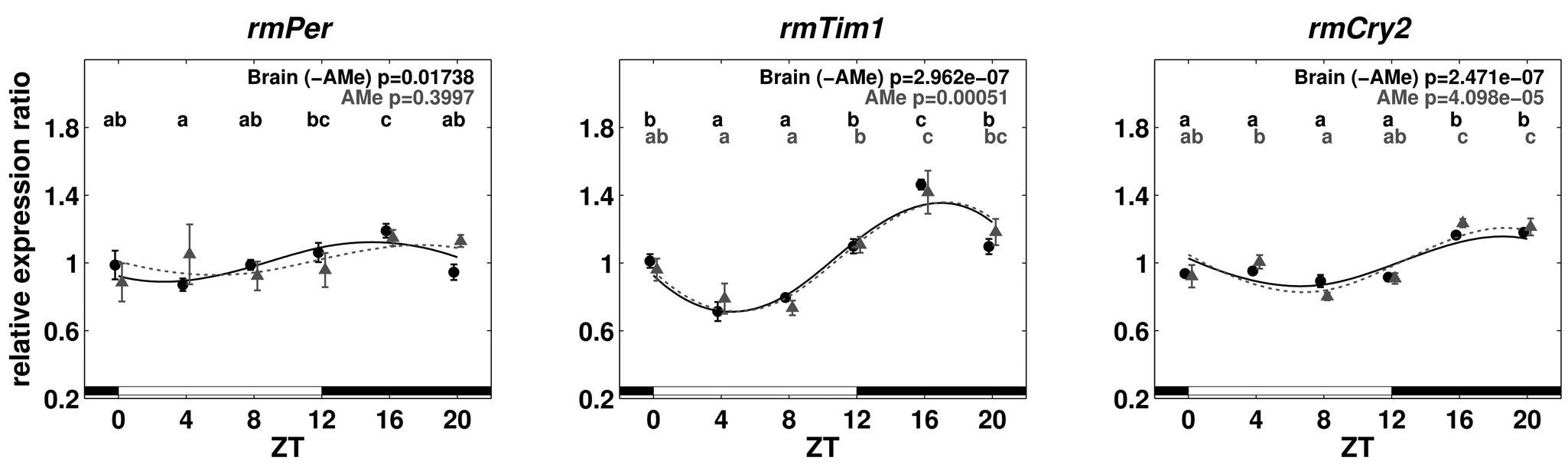
Relative expression ratios of rmPer, rmTim1, and rmCry2 for brain (without AMe, black) and AMe (gray) tissues in the LD 12:12 cycle. The bars at the bottom of the plots indicate light (white) and dark (black) phases. Expression levels of 2 zeitgeber times with different characters above are significantly different (1-way ANOVA with post hoc LSD test, p < 0.05). Error bars represent standard error;
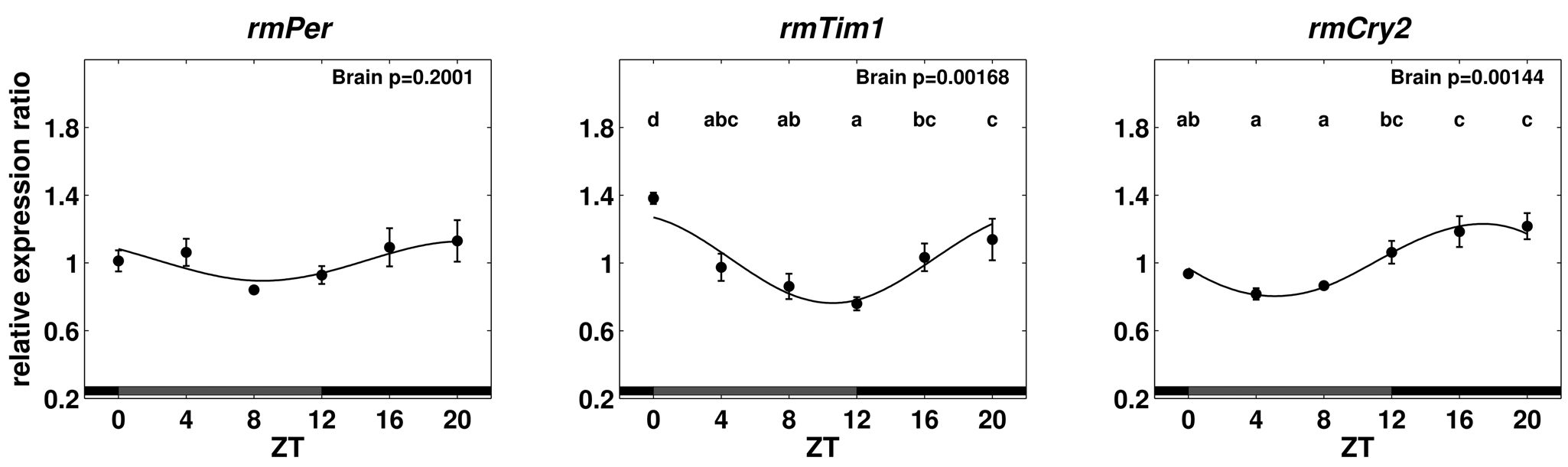
Relative expression ratios of rmPer, rmTim1, and rmCry2 for brain tissue on the first day in DD. For details, see the legend to Figure 3.
In LD 12:12, brain (with the AMe excised) as well as AMe mRNA levels cycled with the highest values occurring in the first half of the night and lowest values in the first half of the day (Fig. 3). Significant differences in mRNA levels between time points for all 3 genes in brain and AMe tissues were obtained (1-way ANOVA with post hoc LSD test, p < 0.05) (Suppl. Table S5), except for rmPer from AMe tissue (Fig. 3). This lack of significance is due to a single pool of tissue at ZT4. This pool was not excluded as an outlier since the same sample grouped with the other pools for rmTim1 and rmCry2. In Malpighian tissue, variation between pools was much higher than in other tissues, and no significant differences between time points in any of the 3 genes were observed (Suppl. Fig. S3 and Suppl. Table S5).
Since there was hardly any difference in expression levels between AMe tissues and the remaining brain, it was decided to use complete brains to monitor the expression levels in DD. Cycling of rmTim1 and rmCry2 mRNA in brain tissue continued the first day in DD with significant differences between time points. Highest values of rmPer expression occurred during the subjective night and lowest during the subjective day but were not significantly different (Fig. 4).
Putative Core Feedback Loop Gene Expression in Different Photoperiods
After examining the daily expression of putative core feedback loop genes in LD 12:12 conditions, the expression levels in cockroaches raised in different photoperiods were compared to examine the influence of day length on clock gene expression in R. maderae. In both short- (LD 6:18) as well as long-day (LD 18:6) conditions, maxima occurred in the first half of the night with a higher amplitude under long-day conditions (Fig. 5). Significant differences between time points were found (1-way ANOVA with post hoc LSD test, p < 0.05) for rmPer, rmTim1, and rmCry2 in short- and long-day conditions. Aligned at ZT0, the long-day oscillation is phase delayed with respect to the short-day oscillation by about one sampling interval (4 hours). Cycling continued the first day in DD with significant differences between time points, except for rmPer and rmCry2 in short-day conditions (1-way ANOVA with post hoc LSD test, p < 0.05) (Fig. 6). The amplitude in long-day conditions was generally higher in the LD cycle than the first day in DD. Strikingly, the expression level of rmTim1 from animals raised under short-day conditions suddenly dropped to low levels at ZT12 the first day in DD.
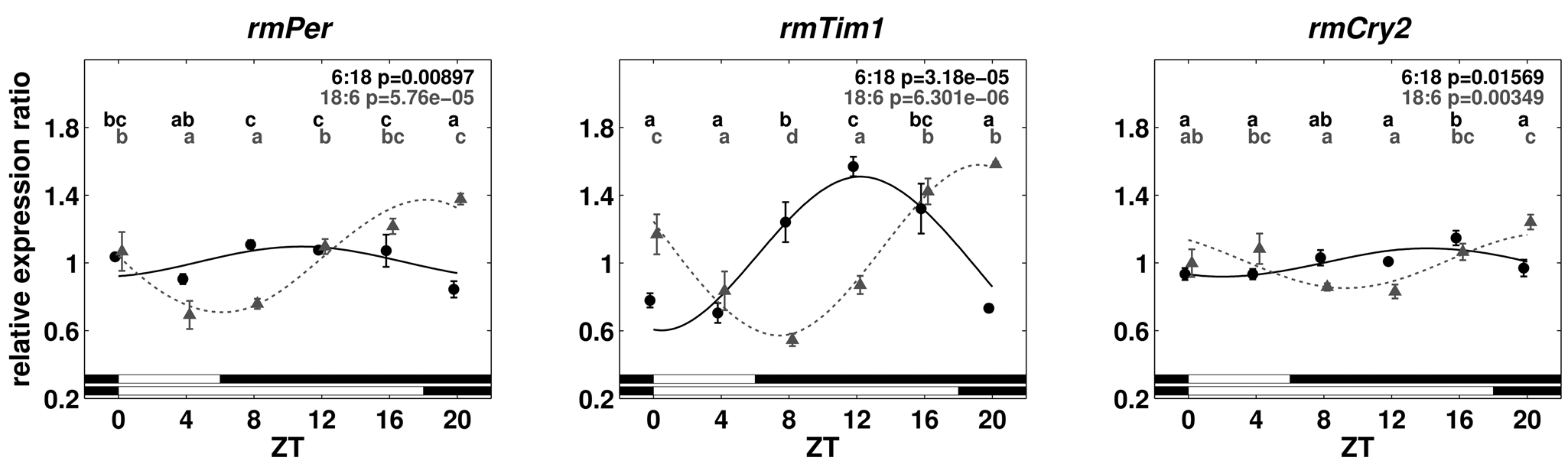
Relative expression ratios of rmPer, rmTim1, and rmCry2 for brain tissue under short- (6:18 LD) and long-day (18:6 LD) conditions. For details, see the legend to Figure 3.
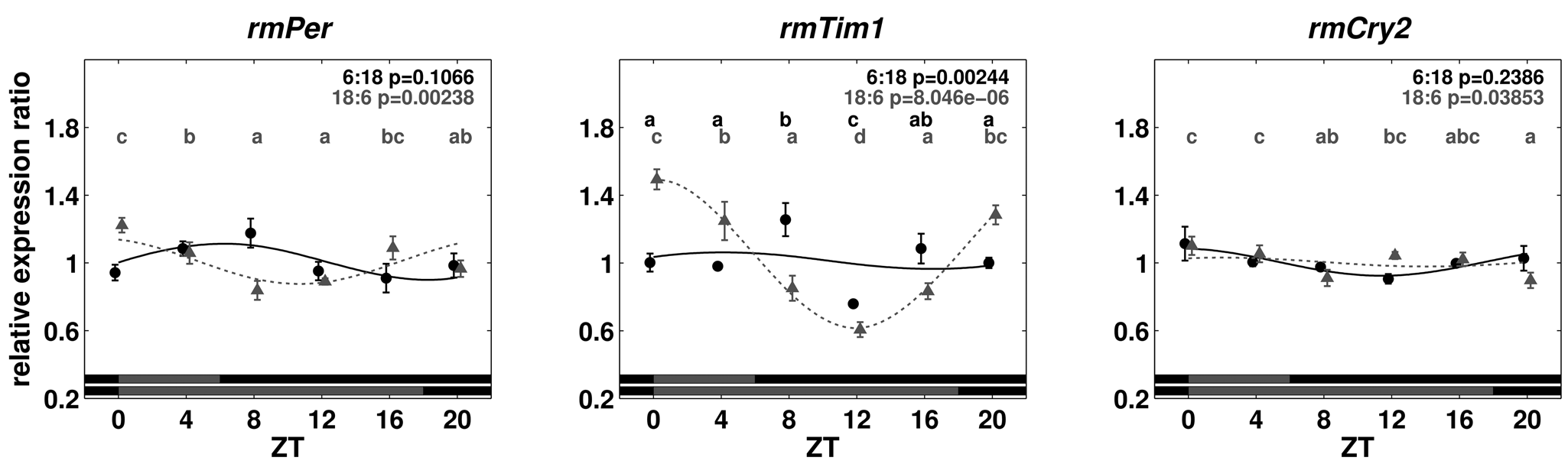
Relative expression ratios of rmPer, rmTim1, and rmCry2 for brain tissue of animals entrained to short- (6:18 LD) and long-day (18:6 LD) conditions on the first day in DD. For details, see the legend to Figure 3.
The daily mean of rmPer, rmTim1, and rmCry2 expression was significantly higher in long- than in short-day conditions (1-way ANOVA, p < 0.05) (Fig. 7B). The daily mean of rmPer and rmCry2 expression was much higher than the rmTim1 expression level in all tissues (Fig. 7A). When moved from short day to DD, the expression levels significantly increased for rmPer and rmTim1 and did not change in rmCry2 (1-way ANOVA, p < 0.05). Reversely, when moved to DD from long-day conditions, expression levels significantly dropped for all 3 genes (1-way ANOVA, p < 0.05) (Fig. 7B).
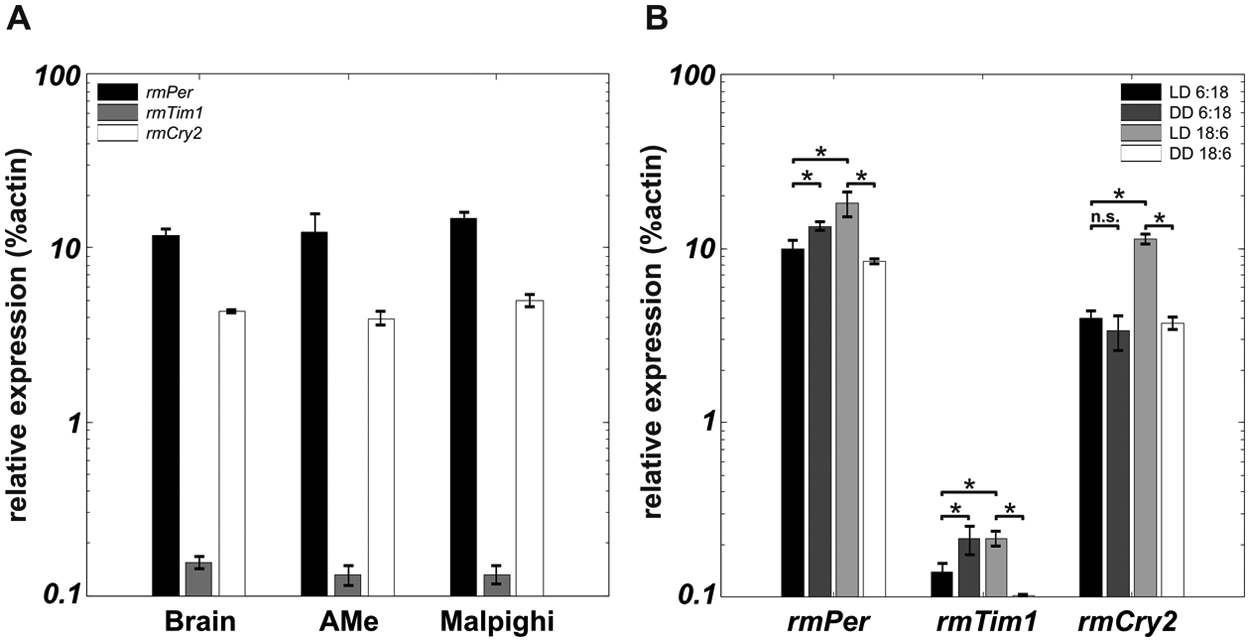
Daily mean of the expression levels of rmPer, rmTim1, and rmCry2 for different tissues (A) and different photoperiods (B). Error bars represent standard error. Asterisks indicate significant differences (1-way ANOVA, p < 0.05). ns = not significant.
Discussion
As a first step towards the characterization of the molecular circadian clock of the Madeira cockroach, full-length coding sequences of R. maderae per and cry2 and a short fragment of tim1 were obtained. In addition, it was shown that rmTim1 and rmCry2 genes are expressed in a circadian rhythm in the brain as well as in the circadian pacemaker center, the AMe, which controls circadian locomotor activity rhythms. Expression of rmPer was not significantly rhythmic in constant conditions. Interestingly, clock gene expression rhythms of this equatorial species are phase shifted in cockroaches raised in different photoperiods.
Circadian Clock Genes in the Madeira Cockroach
Phylogenetic analysis places rmPER next to other hemimetabolous insect species and within cockroach PER proteins (Fig. 1B). Domains known to be necessary for specific protein function in dPER, except the TG repeat, are conserved in rmPER. Instead of a TG repeat, rmPER contains a short repeat of serine-glycine residues, of unknown function, which was also reported in other species. The c7 domain, defined in B. germanica (Lin et al., 2002), is conserved in rmPER as well as in other nondrosophilid insects (Fig. 1C). In mammals, a C-terminal PER region comparable to c7 was shown to bind mCRY2-1 (Tomita et al., 2010). Possibly, the c7 domain binds CRY2 in R. maderae and in other insects as well. However, nematoceran insects like Culex, Anopheles, and Aedes lack a conserved c7 domain but express CRY2. Thus, there may be other CRY2 binding sites in these insects.
Amino acid composition and domain structure identify rmCRY2 as a “mammalian-type” insect CRY2. Functional domains first determined in mammals (Hirayama et al., 2003; Chaves et al., 2006) are conserved in rmCRY2 (Fig. 2A). A region necessary in mammals for CRY2 interaction with PER2 (Ozber et al., 2010) is also well conserved (amino acids 475-501; 81% identity). Even the amino acids (Arg-487 and Lys-489), which were shown to be necessary for PER2 interaction (Ozber et al., 2010), are identical in the Madeira cockroach. Together with the rhythmic expression in DD, this indicates that rmCRY2 is part of the core feedback loop rather than a blue light receptor only. PER-TIM interactions with CRY2 were shown in D. plexippus (Zhu et al., 2008), and although CRY2 proteins can be activated by light (Hoang et al., 2008), it is not clear whether CRY2 function is light dependent, independent, or both in insects.
rmPer, rmTim1, and rmCry2 expression is rhythmic and cycles in the same phase in the brain of the Madeira cockroach in LD 12:12 conditions, with peaks in the first half of the dark phase (Fig. 3), as in D. melanogaster, D. plexippus (Zhu et al., 2008), and A. mellifera (Rubin et al., 2006). This indicates that these expression rhythms are driven by the same regulatory elements in these various insect species. Since cycling of rmTim1 and rmCry2 continued the first day in DD (Fig. 4), these are very likely circadian clock genes in R. maderae. Differences between time points were not significant for rmPer the first day in DD. Possibly, the already low amplitude cycling in LD 12:12 further damped due to uncoupling oscillators in the brain. Rhythmic expression occurred with the same phase in tissue samples containing the isolated AMe with associated neurons as well as in the rest of the brain without the AMe. Therefore, rmPer, rmTim1, and rmCry2 cycle with the same phase in the majority of cells in both tissues. However, not all cells appear to be phase locked: Higher variation between pools in AMe-containing tissues, especially in rmPer, may be caused by different circadian pacemaker cells in the AMe, which cycle out of phase. Furthermore, there must be additional circadian pacemaker cells in the brain next to the AMe pacemakers since we observed rhythmic expression in brain tissue not containing the AMe. For example, PDF immunoreactive neurons occur in 2 clusters of cells in the lamina and in 2 clusters, the anterior (aPDFMes) and the posterior cells, next to the AMe (Homberg et al., 1991; Petri et al., 1995). While only the aPDFMes were shown to be circadian pacemaker neurons, which control locomotor activity rhythms, it is likely that also the other PDF immunoreactive neurons are circadian pacemaker cells as well (Homberg et al., 2003). Also, as has been shown in D. melanogaster (Zerr et al., 1990; Ewer et al., 1992; Suh and Jackson, 2007), glial cells of the Madeira cockroach brain may express circadian genes as well. Indeed, preliminary immunostainings employing antibodies raised against PER from different species as well as against rmPER showed widespread PER expression within the brain (unpublished results), indicating that the majority of cells express rmPER. Since these findings are beyond the scope of this study, they will be published elsewhere. Interestingly, the expression level of rmTim1 is much lower than the expression level of rmPer or rmCry2 (Fig. 7A and 7B). It nevertheless exhibits the highest amplitude of cycling. Of the available TIM1 sequences, the fragment we cloned shares the highest identity with P. americana TIM1 (93%), followed by G. bimaculatus (75%) and Thermobia domestica TIM1 (74%). Knockdown experiments showed that the role of TIM1 can be quite diverse. Whereas in G. bimaculatus the clock seems to work without TIM1 (Danbara et al., 2010), it is an essential component in T. domestica (Kamae and Tomioka, 2012). Peak values of rmTim1 expression occurred during the first half of the night with a similar amplitude as in G. bimaculatus optic lobes. However, with only part of rmTim1 cloned and with the experiments performed, it cannot be decided whether rmTim1 is an essential component of the circadian clock or only has a modulatory role.
The rather weak circadian modulation in expression levels of rmPer, rmCry2 (<2-fold), and rmTim1 (~2.5-fold) was also reported in other hemimetabolous insects (Hodkova et al., 2003; Syrová et al., 2003; Ikeno et al., 2008; Cortés et al., 2010). This is in contrast to many holometabolous insects in which there are changes of often more than 3-fold (Reppert and Sauman, 1995; So and Rosbash, 1997; Cyran et al., 2003; Rubin et al., 2006; Zhu et al., 2008) and up to 7-fold in A. mellifera cry2 (Rubin et al., 2006). It was suggested that the weak circadian modulation in hemimetabolous insects is due to masking effects of peripheral tissue, when complete or large compartments of heads were used (Hodkova et al., 2003; Ikeno et al., 2008). However, if masking effects are responsible for low-amplitude cycling in R. maderae, differently phased oscillators must reside inside the neuronal tissue itself since we used precisely explanted tissue in our experiments.
Rhythmic expression of per and tim1 in Malpighian tubules was shown in D. melanogaster (Giebultowicz et al., 2000). In R. maderae, we did not find any significant differences in expression levels of rmPer, rmTim1, and rmCry2 between all time points in Malpighian tubules (Suppl. Fig. S3). Also, variation between pools was higher than in neuronal tissues. This is similar to G. bimaculatus, where also no clear rhythmic expression of per and tim1 could be observed (Uryu and Tomioka, 2010). A tissue-specific role of these clock genes could result in a noncircadian expression, or possibly differently phased molecular clocks exist in Malpighian tubules of some species.
Photoperiod-Dependent Plasticity
Phase and amplitude of clock gene expression are clearly light-dependently regulated: First, as in D. melanogaster (Majercak et al., 1999) and other insects (Sarcophaga bullata: Goto and Denlinger [2002]; P. terraenovae: Muguruma et al. [2010]), peak levels of rmPer, rmTim1, and rmCry2 adjusted relative to the onset of the scotophase, with maxima at the beginning of the night in all photoperiods tested (Figs. 3 and 5). Second, the rhythm’s amplitude is lower in short days versus long days. Third, the daily mean of expression levels is higher in long days versus short days. In contrast, in other insects, clock gene expression levels were either higher (Dolezel et al., 2007; Cortés et al., 2010) or remained unchanged in short days (Ikeno et al., 2008). In P. apterus, strong changes in per levels reflected changes in physiological conditions rather than being a direct consequence of the photoperiod (Dolezel et al., 2007). Possibly, also upregulation of circadian genes in short days may be due to masking via changed physiological conditions in these diapausing insects. Whether the circadian clock works similar in R. maderae and in D. melanogaster remains to be studied. Either light directly or indirectly activates clock gene expression, or higher mean expression levels in long days are due to slower protein accumulation to lower levels. Since the gene products inhibit their own expression, this results in a higher daily mean of expression rate. Expression levels in LD conditions are influenced mainly by the photic input and the feedback loop system itself. In constant conditions, expression levels are a result of the endogenous feedback loop system alone. Thus, if moved to DD from different photoperiods, the mean daily expression level should move towards its endogenous state uninfluenced by light, which means an increase from short-day conditions and a decrease from long-day conditions. Since this was exactly observed in the Madeira cockroach, it appears to possess conserved circadian clockworks, which are regulated tightly by light.
Footnotes
Acknowledgements
The authors thank Dr. Penelope Higgs for providing laboratory instruments, Prof. Dr. Ralf Stanewsky for critically reading the article, and Prof. Dr. Horst Bohn for his remarks on R. maderae nomenclature.
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.