
Abstract
Locusts demonstrate remarkable phenotypic plasticity driven by changes in population density. This density dependent phase polyphenism is associated with many physiological, behavioral, and morphological changes, including observations that cryptic solitarious (solitary-reared) individuals start to fly at dusk, whereas gregarious (crowd-reared) individuals are day-active. We have recorded for 24-36 h, from an identified visual output neuron, the descending contralateral movement detector (DCMD) of Schistocerca gregaria in solitarious and gregarious animals. DCMD signals impending collision and participates in flight avoidance maneuvers. The strength of DCMD’s response to looming stimuli, characterized by the number of evoked spikes and peak firing rate, varies approximately sinusoidally with a period close to 24 h under constant light in solitarious locusts. In gregarious individuals the 24-h pattern is more complex, being modified by secondary ultradian rhythms. DCMD’s strongest responses occur around expected dusk in solitarious locusts but up to 6 h earlier in gregarious locusts, matching the times of day at which locusts of each type are most active. We thus demonstrate a neuronal correlate of a temporal shift in behavior that is observed in gregarious locusts. Our ability to alter the nature of a circadian rhythm by manipulating the rearing density of locusts under identical light-dark cycles may provide important tools to investigate further the mechanisms underlying diurnal rhythmicity.
Keywords
Circadian rhythms present in almost all organisms permit behavioral and physiological preparations for predictable events. We have sought to determine whether circadian rhythmicity in the responses of a visual neuron of the locust Schistocerca gregaria differs in individuals in the swarming (gregarious) and nonswarming (solitarious) phases, which are active at different times of the day.
In the insect visual system, endogenous circadian changes in eye structure and function prepare the visual system to cope with predictable daily cycles of light intensity. Changes in ambient light intensity in turn induce further modifications of eye structure, and these processes together permit eyes to function effectively over a large range of light intensities (Fleissner and Fleissner, 2006). Endogenous circadian rhythms can also be entrained by changes in ambient light intensity, thus providing a mechanism for plastic changes in their expression. In Drosophila, for example, the timing of behavior in the 24-h period is shifted to correspond with seasonally changing day length. Such plasticity means that an ongoing circadian rhythm can give rise to differing rhythmic patterns of physiology or behavior that are appropriate to the ambient conditions (Stoleru, 2007; Dubruille and Emery, 2008).
A second type of plasticity in circadian rhythmicity is evident in eusocial insects where there is typically division of labor between different castes or ages. Different individuals express different patterns of circadian behavior appropriate to their caste-specific function in the colony, and single individuals can express different patterns at different times during their life. For example, young worker bees typically nurse the brood in a dark hive without showing circadian patterns of activity, but they develop diel rhythms of activity before beginning to forage outside the hive in strongly diurnal patterns as they get older (Moore et al., 1998). In Drosophila, which is not a eusocial insect, interactions between conspecifics can also influence the circadian clock, with olfactory cues playing a key role (Levine et al., 2002a). Pairs of adult Drosophila housed together alter the timing of their locomotory behavior from the normal rhythm, driven by male courtship and copulatory behavior that is elicited by stimuli emitted from the female (Fujii et al., 2007).
The desert locust (Schistocerca gregaria) shows remarkable phenotypic plasticity, expressed as differences in behavior, endocrine physiology, neurophysiology, neurochemistry, morphology, and life history. This suite of changes is driven by fluctuations in population density brought about by changes in the environment (Uvarov, 1966; Simpson et al., 1999). At low population densities, locusts are solitarious, being cryptic in both appearance and behavior. They walk slowly with a creeping gait, fly mostly at night, have a restricted dietary range, and avoid contact with conspecifics. When solitarious locusts are forced together, typically as a result of increasing population size or a reduction in food availability, they become gregarious. Gregarious locusts walk with a rapid upright gait and have a broad diet. Juveniles show bright warning colors, and adults fly during the day. Gregarious locusts are actively attracted to conspecifics, which promotes the formation of swarms. The key change from solitarious to gregarious behavior takes just a few hours (Roessingh et al., 1993; Anstey et al., 2009), but morphological changes take several generations to become established (Pener, 1991). Density dependent polyphenism is accompanied by large changes in the amounts of many neuromodulators and neurotransmitters in the CNS (Rogers et al., 2004).
Solitarious and gregarious locusts express different diel patterns of activity in both the field and the laboratory (Roffey and Popov 1968; Uvarov, 1977; Michel, 1980; Steedman, 1990; Ely et al., 2011). Gregarious locusts are generally day-active, whereas activity of solitarious locusts peaks 1-2 h after dusk (Steedman, 1990; Ely et al., 2011). Evening and night activity of solitarious locusts might reduce their exposure to diurnally active avian predators (Steedman, 1990).
In locusts, as in many other animals, evasion of predators is facilitated in part by activation of visual pathways that are tuned to the detection of looming objects (Santer et al., 2005, 2006). One such pathway consists of a well characterized pair of brain interneurons, the lobula giant movement detector (LGMD) and its postsynaptic target the descending contralateral movement detector (DCMD). LGMD responds most strongly to objects looming on a direct collision course (Gabbiani et al., 1999, 2002; Hatsopoulos et al., 1995; Rind and Simmons, 1992, 1997; Schlotterer, 1977; Simmons and Rind, 1992) and excites DCMD at a synapse that transmits all presynaptic action potentials with 1:1 fidelity (O’Shea and Williams, 1974; Rind, 1984). Spikes in the descending axon of DCMD are the largest visible in extracellular recordings made from the longitudinal connectives of the ventral nerve cord, which facilitates their detection and analysis. DCMD makes excitatory synapses onto leg motor neurons and flight motor neurons in the thorax (Pearson and Goodman, 1979; Simmons, 1980). It has previously been shown that the visual receptive field of the LGMD/DCMD system differs in solitarious and gregarious locusts (Rogers et al., 2010) and that the nature of the output synapse of DCMD onto a leg motor neuron also differs in a way that tunes the collision-detecting pathway to the different lifestyles of the solitarious and gregarious animals (Rogers et al., 2007; Matheson et al., 2004).
In the present article we show that the responsiveness of the LGMD/DCMD system to looming objects changes in a circadian rhythm under constant light. Moreover, the strongest responses in solitarious and gregarious locusts occur at different times within the 24-h period, matching the timing of peak flight activity.
Materials and Methods
Solitarious and gregarious locusts (Schistocerca gregaria Forskål) were maintained under a 12-h/12-h light-dark regimen in conditions matched for temperature (36 °C light period, 25 °C dark period), illuminance (75-150 lux), and food availability (fresh wheat seedlings and bran flakes ad libitum). Solitarious locusts (reared in isolation for 3-4 generations) were obtained from the colony maintained in the Department of Zoology, University of Cambridge. They were kept as isolated individuals in small (10 × 10 × 20 cm) containers, whereas gregarious locusts were kept at densities of 100-200 per cage (50 × 50 × 50 cm). These rearing conditions reliably produce solitarious and gregarious locusts in our laboratory and elsewhere (Simpson et al., 1999; Matheson et al., 2004).
Using a CRT (cathode ray tube) monitor, we simulated the looming approach of a dark square (contrast 72%) against a green background. The refresh rate (160 Hz) and spatial resolution (<0.22° visual angle) of the monitor were sufficiently high to ensure that locusts could not perceive the stepwise increases in stimulus size (Howard, 1981; Horridge, 1978). The rate of expansion of the object is given by the ratio of the half size of the object (l) to the absolute value of the approach velocity (v). For all of these experiments, a value of l/v = 40 sec−1 was used, with the approach simulating the last 5 sec before the predicted time of collision with the eye. Stimuli approached the eye from directly orthogonal to the animal, level with the center of the eye.
All experiments were carried out under constant low-level lighting (105 lux) at approximately 25 °C. Locusts of both sexes were used. Previous work has shown no differences in DCMD responses between males and females (Matheson et al., 2004; Rind et al., 2008). Experiments were started at different times of day (between 1000 and 1700 h). The first looming approach was presented at least 30 min after the preparation was set up, and subsequent approaches were presented at 1-h intervals for the following 24 h.
Recordings lasting 24-36 h were made from 9 solitarious and 8 gregarious locusts. Of the solitarious recordings, 3 were arrhythmic and 6 showed robust circadian rhythms (2 male, 4 female). Among the gregarious recordings, 2 were arrhythmic and 6 showed robust circadian rhythms (2 male, 4 female). Constant light conditions can cause behavioral arrhythmia in several organisms due to inhibitory effects on components of the molecular circadian clock (Marrus et al., 1996, Emery et al., 2000), so only the 12 records in which the periods of the fitted sine curves fell within the circadian range (18-31 h) were used in the subsequent analyses.
In both solitarious and gregarious locusts, DCMD responded to the simulated approach of a looming object (Fig. 1, lower trace) with a train of spikes (Fig. 1, middle trace) recorded from the cervical connectives using a bipolar silver hook electrode, a Neurolog AC-coupled extracellular amplifier, and a micro1401plus data acquisition system running Spike 2 software (both Cambridge Electronic Design) (Rogers et al., 2007). The DCMD spikes could be readily identified as they have the largest amplitude of any spikes in these recordings (Matheson et al., 2004). The firing rate increased throughout the approach and then declined rapidly just before the anticipated collision (Fig. 1). We convolved the spike train with a Gaussian smoothing function having a standard deviation of 20 msec and amplitude of 25.1 Hz and computed the number of spikes in each approach, the peak firing rate, and the time of peak firing relative to the predicted time of collision using custom scripts in Spike 2 (Gabbiani et al., 1999). The data were fitted with 4 parameter sine curves using the equation Y = Y0 + a·sin[(2πx / b) + c], where Y0 = offset, a = amplitude, b = period, and c = phase (SigmaPlot: Systat Software Inc.). Such sine functions are routinely used to describe circadian data (Barth et al., 2010).
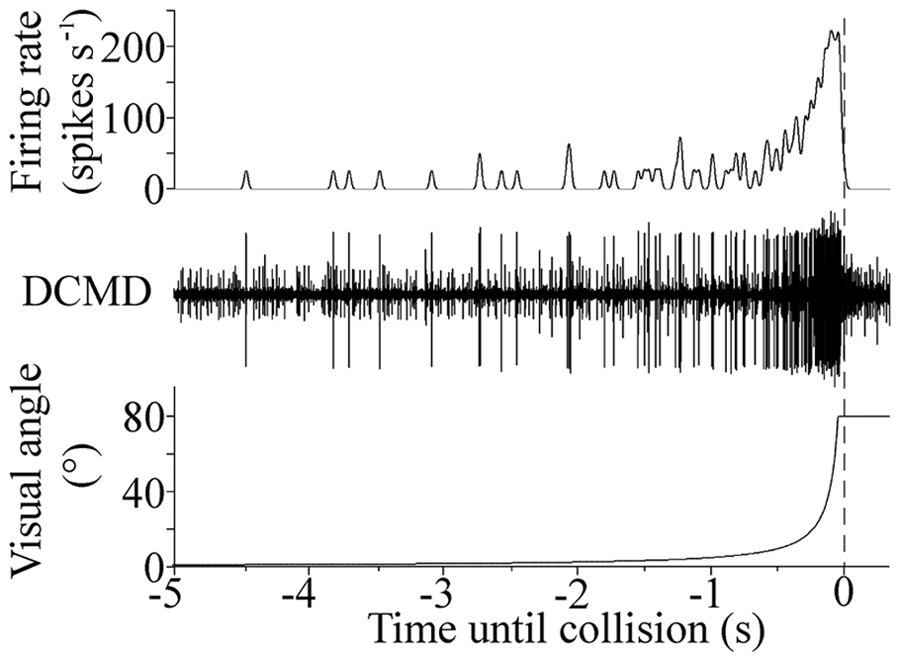
Response of DCMD to a single looming stimulus. Lower trace: angular size of the simulated approaching object. Middle trace: a recording from the cervical connective containing the axon of DCMD revealed the neuron’s characteristic response (largest spikes). Upper trace: smoothed firing rate of DCMD. The dashed line represents the predicted collision time.
We applied a discrete Fourier transform to the unfiltered data to inspect the underlying basis for diel patterns of activity in DCMD (Chatfield, 1989; Dowse, 2009; Dowse and Ringo, 1989). For this part of the analysis we made the assumption that the data were cyclical in nature and that the pattern would repeat with a 24-h period. We first removed the mean from the signal and detrended it by fitting a regression line and subtracting it. The power in the series was thus the same as the variance. Fourier analysis decomposes the signal orthogonally and partitions the variance by period; hence, each coefficient is the power attributable to that period, expressed as relative power without units. We calculated Fourier coefficients for periods ranging from 2 h, the shortest period that can be treated given the 1 h–1 sampling rate (Nyquist frequency), up to 12 h, which is the longest period justifiable with 24 h of data but restricted our further analysis to periods of 4-12 h (Bennett, 1983).
Results
Circadian Rhythms in the Responses of Individual Animals
To determine whether the output of DCMD is modulated by an endogenous circadian rhythm, approaches by a looming stimulus were simulated at 1-h intervals for at least 24 h during maintained recordings of the DCMD response. We analyzed the number of spikes, peak firing rate, time of peak firing, and background firing rate for each approach. We present here the analysis for number of spikes per approach. The further analyses are presented in the supplementary information.
DCMD recordings from a solitarious locust (Fig. 2A) and a gregarious locust (Fig. 2B) showed that the number of spikes evoked by a 5-sec approach varied approximately sinusoidally over the 24-h period of constant low level light, with a minimum around subjective dawn and a maximum in the subjective afternoon or evening. The mean number of spikes per approach averaged across the entire 24-h sampling period did not differ significantly between gregarious and solitarious locusts (gregarious, 120 ± 15 spikes per approach; solitarious, 127 ± 21 spikes per approach, t-test, df = 10, p = 0.56). A continuous recording of 36 h (Fig. 2C) showed that the circadian rhythm persisted into a second period.
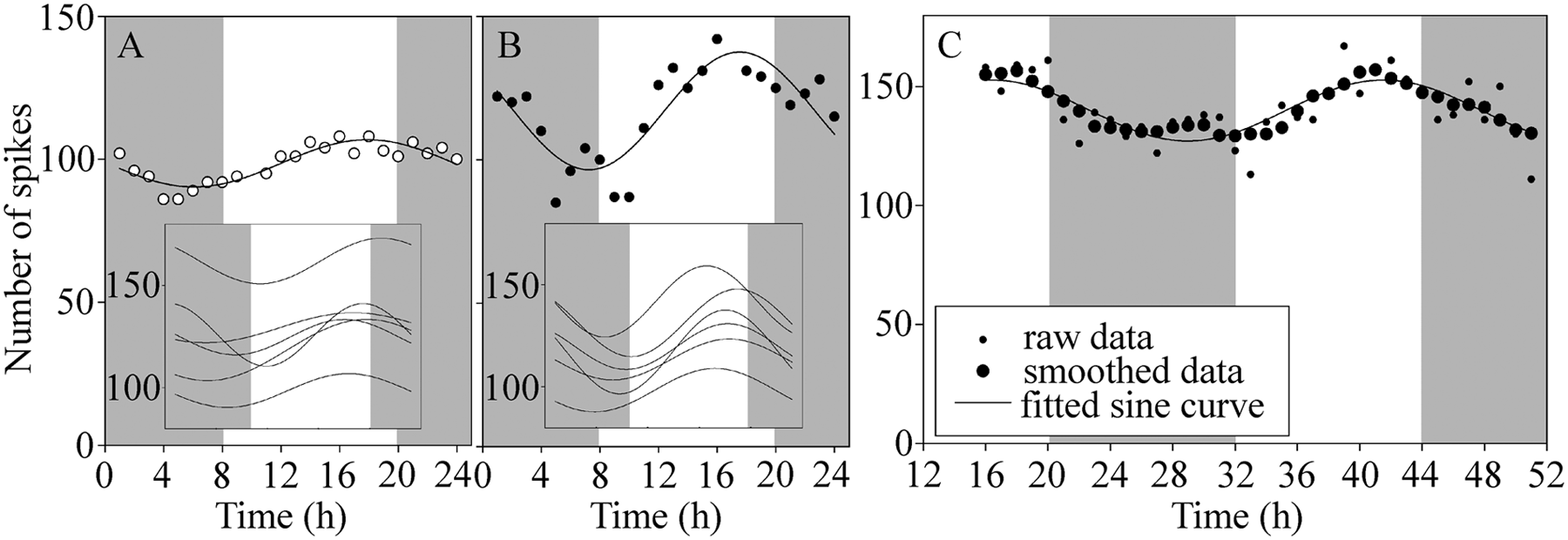
DCMD’s response varied over 24 h under constant light in both solitarious and gregarious locusts, as shown by plots of the number of DCMD spikes elicited by a looming stimulus presented at 1-h intervals over 24-36 h. (A) Responses of a single solitarious locust over 24 h with a fitted sine curve. Inset: fitted sine curves for all solitarious locusts used in subsequent analyses. (B) Responses and fitted sine curve for a gregarious locust. Inset: fitted sine curves for all gregarious locusts used in subsequent analyses. (C) Number of spikes (small symbols = raw data; large symbols = 5-point moving average) and sine curve fitted to the smoothed data for a single gregarious locust recorded for 36 h. Shaded areas represent time periods when the locusts would have normally experienced darkness. The start times for these experiments were (A) 1100 h, (B) 1000 h, (C) 1600 h.
Comparison of the Responses of Solitarious and Gregarious Locusts
To compare the patterns of DCMD response between solitarious and gregarious locusts, we normalized the data for each neuron by dividing each measurement by the mean obtained from responses over the whole recording. The resulting values are plotted as means ± SD in Figure 3 for 6 solitarious and 6 gregarious animals. We then took 2 approaches to assessing the data. First, we fitted the averaged normalized data with sine functions to provide an initial estimate of rhythmicity (solid lines in Fig. 3). Second, we filtered the normalized data for each animal with a low-pass 1-2-3-2-1 weighted sliding mean that reflected at the beginning and end of the measurement period (Hamming, 1983), and we determined the times of maximum and minimum response for each animal. We plotted these values as means ± SD in Figure 3. We then averaged the filtered data and plotted the mean curves as dashed lines, indicating their peaks with arrows in Figure 3. Carrying out these approaches in parallel enabled us to identify deviations from a purely sinusoidal pattern of response. We later use a Fourier analysis to examine the basis for these patterns in more detail.
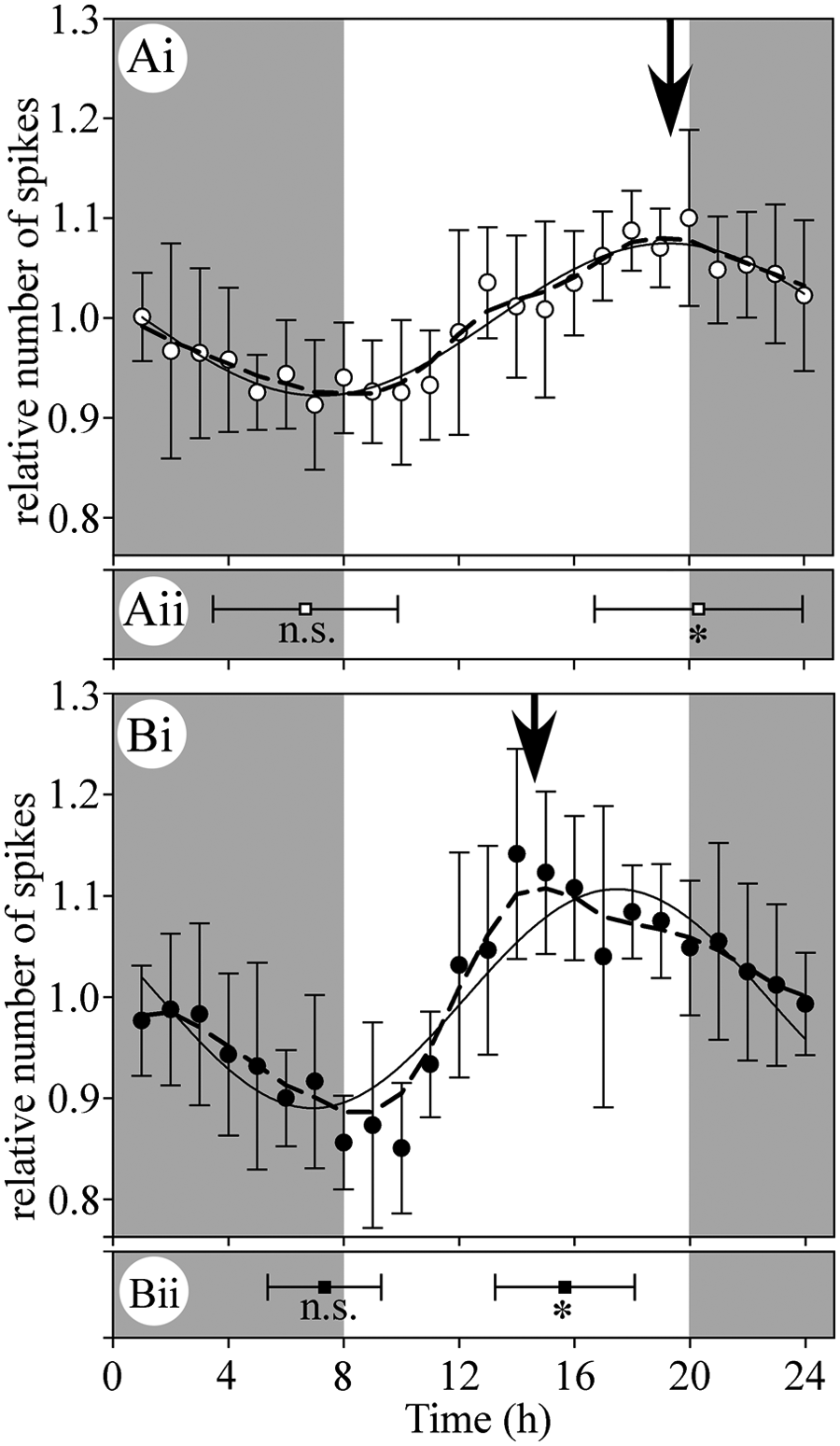
Differences in circadian rhythms of responsiveness in a visual interneuron. Circadian rhythms in DCMD responses of solitarious (open symbols) and gregarious (solid symbols) locusts peaked at different times of subjective day. (Ai) Relative number of spikes in solitarious animals (R2 = 0.9221). (Bi) Relative number of spikes in gregarious animals (R2 = 0.7871). Each point is the mean ± SD of 6 animals, and the data are fitted with 4 parameter sine functions (solid lines). Vertical arrows mark the maxima of the averaged filtered data (dashed lines). The mean and standard deviation of the times of minimum and maximum responses calculated from the filtered normalized data for each locust are shown for solitarious (Aii) and gregarious (Bii) locusts. The times of the morning minima in Aii and Bii did not differ in solitarious and gregarious locusts (ns), but the times of the afternoon maxima did (asterisks, p = 0.025). Shaded areas represent time periods when the locusts would normally have experienced darkness. Corresponding data for peak firing rate are shown in Supplementary Figure S1.
Both solitarious and gregarious locusts had minimum responses around expected dawn, but the time of subjective day at which their maximum responses occurred differed by several hours. As shown for the individual data illustrated in Figure 2, the mean data across animals were well fitted by a sine function for solitarious locusts (Fig. 3A). For gregarious animals, however, there were clear deviations from a purely sinusoidal pattern (Fig. 3B), with the responsiveness remaining high throughout the second half of the subjective day rather than rising to a discrete peak. The strength of the response increased relatively rapidly to a maximum value, followed by a slower decline over the rest of the subjective day. The maximum response for solitarious animals occurred at the time of expected dusk (1900 h, arrow in Fig. 3A), whereas that for gregarious animals occurred 5 h earlier, approximately halfway through the anticipated light period (at 1400 h, arrow in Fig. 3B). Note that this peak is earlier than the maximum in the sine curve because the pattern of response in gregarious animals deviates from a simple sine function (see “Periodicity of Response Modulation” below). The minimum response for both solitarious and gregarious locusts occurred around the time that dawn would have been expected (0800 h and 0830 h, respectively, Fig. 3A, B). We used the maximum and minimum times of subjective day extracted from the filtered data for each individual to test for differences between solitarious and gregarious animals. The time of maximum response for solitarious animals was later in the subjective day than that for gregarious animals (Fig. 3: t-test, df = 10, p = 0.025), but there was no significant difference in the time of minimum response (Fig. 3: t-test, df = 10, p = 0.673). The difference in response amplitude between the morning minimum and the afternoon/evening maximum was 23% ± 4% for gregarious locusts and 17% ± 6% for solitarious locusts, relative to the mean response over 24 h (t-test, df = 10, p = 0.052). Identical analyses of DCMD’s peak firing rate during each simulated approach revealed patterns similar to those described above for the number of spikes per approach (Suppl. Fig. S1A, B).
The largest divergence in strength of DCMD response between solitarious and gregarious locusts occurred during the expected daytime. The number of spikes per approach was relatively larger in gregarious animals between 1100 h and 1645 h, and there was a smaller peak approximately 12 h earlier (Fig. 4). A very similar pattern was seen for the peak firing rate per approach (Suppl. Fig. S2).
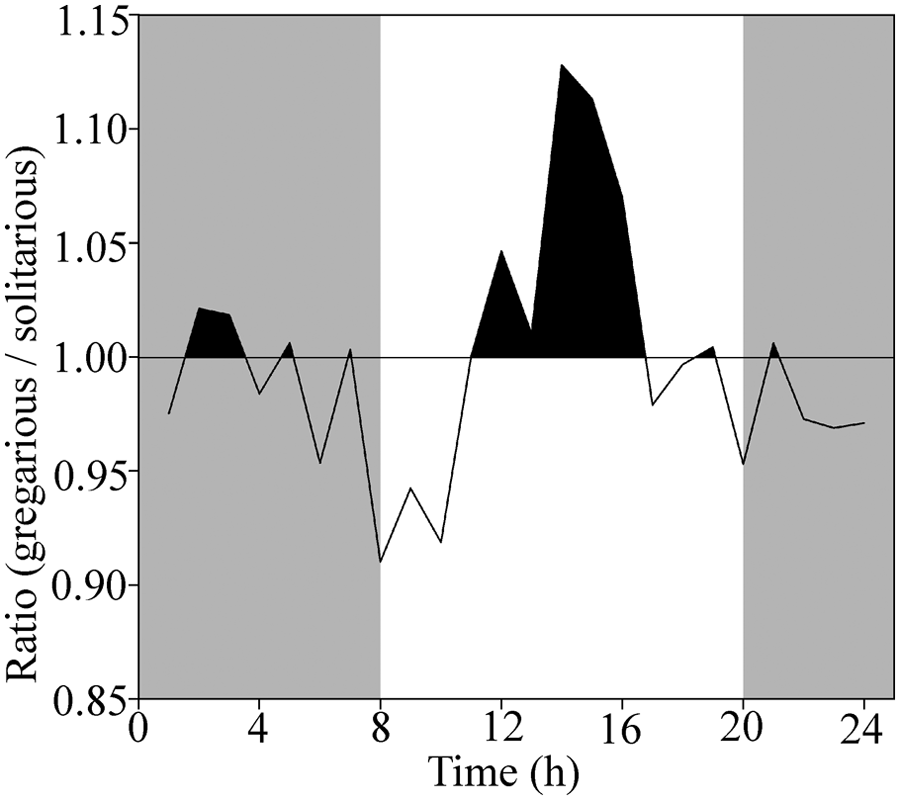
Temporal differences in response. The number of spikes elicited in DCMD by the sight of a looming object was greater in gregarious locusts than solitarious locusts during the subjective daytime (areas shaded black). Areas shaded gray represent time periods when the locusts would normally have experienced darkness. The values on the ordinate show the ratio of the number of spikes in gregarious locusts to the number in solitarious locusts. Corresponding data for peak firing rate are shown in Supplementary Figure S2.
For each individual approach we also measured the time (relative to predicted collision) at which the DCMD firing rate reached a maximum (see Fig. 1). There was no clear circadian pattern of variation in this time of peak firing in either solitarious or gregarious locusts (Suppl. Fig. S1C, D). DCMD had a low spontaneous firing rate (solitarious median 0.15 spikes per second, gregarious median 0.267 spikes per second) that also did not exhibit a circadian rhythm (Suppl. Fig. S1E, F). An apparent increase in the variability of background firing during the subjective day was caused by occasional brief spontaneous bursts of spikes in DCMD (up to 40 spikes per second).
Periodicity of Response Modulation
The highly sinusoidal circadian pattern of DCMD response seen in solitarious locusts (Fig. 3A) was reflected in relatively low Fourier coefficients for all periods of less than 24 h (open symbols in Fig. 5). The more complex circadian pattern of response for gregarious animals (Fig. 3B) was associated with higher Fourier coefficients at each period from 4 h to 12 h (solid symbols in Fig. 5). The sets of Fourier coefficients differed in solitarious and gregarious animals (ANOVA for periods of 4-12 h, F = 5.094, p = 0.048), indicating that the patterns of response in gregarious animals had more power in these periods than did those of solitarious locusts. A similar pattern in all respects was seen for peak firing rate (Suppl. Fig. S3).
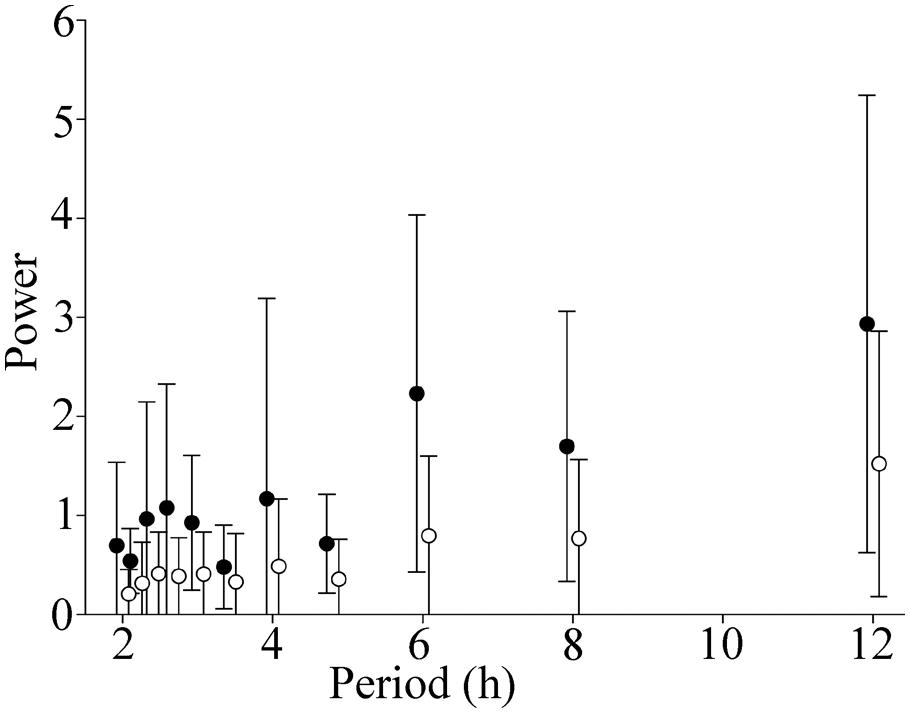
Ultradian rhythms contribute to the complexity of the circadian rhythm in DCMD. Fourier coefficients are expressed as normalized power, without units. The circadian pattern of response of DCMD of gregarious animals had more power in 4- to 12-h components than did that of solitarious animals, indicating the presence of stronger additional underlying rhythms. Values are the mean ± SD Fourier coefficients for relative number of spikes in gregarious (filled circles) and solitarious (open circles) locusts. Corresponding data for peak firing rate are shown in Supplementary Figure S3.
Discussion
The remarkable density-dependent phenotypic plasticity of locusts is well documented (Applebaum and Heifetz, 1999) with striking differences in morphology, ecology, physiology, and behavior. Solitarious locusts do not engage in spontaneous flight during the day (Waloff, 1963) although they are commonly seen flying between dusk and midnight. The flights begin in response to rapidly decreasing solar radiation and discontinue as the temperature falls below 25 °C (Roffey, 1963). In contrast, gregarious locusts fly during the day, with swarms finally settling in the late afternoon (Roffey, 1963). There is thus a good correlation between the observed times for the initiation of flight by solitarious and gregarious locusts and their corresponding high levels in DCMD’s response to impending collision. Our data provide the first evidence for alterations in neuronal function associated with different temporal patterns of behavior induced by density-dependent polyphenism in locusts (Pener and Simpson, 2009).
Colliding with an object in the environment or failing to detect the approach of a predator can be a fatal mistake for any animal. To avoid such outcomes, animals must detect objects approaching on a collision course, determine the likely time of collision, and take appropriately timed and directed avoidance action. Visual neurons tuned to detect looming objects are found in both vertebrates (Sun and Frost, 1998) and invertebrates (Gabbiani et al., 1999; Rind and Simmons, 1992). DCMD and its presynaptic neuron LGMD (O’Shea and Williams, 1974) fulfill all the requirements of such looming detector neurons, being highly tuned to the approach of objects on a direct collision course, having outputs to the flight and leg motor systems of the thorax, and being involved in turning or escape responses during flight (Santer et al., 2006). DCMD has also been implicated in the control of jumping escape behaviors, acting in synergy with other descending visual interneurons (Fotowat et al., 2011). Gregarious locusts show stronger DCMD responses, which are more resistant to habituation in the face of repeated looming stimuli, than those of solitarious locusts (Rogers et al., 2010; Rogers et al., 2007; Matheson et al., 2004).
Although collision avoidance is presumably important at all times, flying insects such as locusts may well benefit from increased responsiveness at times of peak flying activity. Arousal has been shown to have a significant effect on DCMD responsiveness (Rind et al., 2008) with locusts aroused either by leg stimulation or enforced flight showing an elevated spike frequency and reduced habituation. Both mechanical stimulation prior to takeoff and increased arousal during flight would result in an increase in DCMD responsiveness, probably due to raised levels of octopamine (Rind et al., 2008).
Changes in the sensitivity of eyes during a diurnal light-dark cycle occur at the level of the retina in several species of insects, for example, Dytiscus (Jahn and Wulff, 1943), butterflies (Swihart, 1963), Drosophila (Chen et al., 1992), and cockroach (Wills et al., 1985). There is an increase in sensitivity at night in the locust Valanga (Horridge et al., 1981). The sensitivity of locust photoreceptors follows a diurnal rhythm as a result of changes in photoreceptor membrane properties (Cuttle et al., 1995) and the extent of rhabdom turnover (Williams, 1983). Circadian changes under constant dark conditions have also been recorded in the optic neuropils of insects, in the lamina and lobula plate of flies (Pyza and Meinertzhagen, 1993; Bult et al., 1991), and in the lobula complex of cockroaches (Bult and Mastebroek, 1993) and bees (Kaiser and Steiner-Kaiser, 1983). The circadian rhythmicity we have demonstrated in DCMD spiking activity could therefore arise from changes in LGMD and/or DCMD or from changes in neurons presynaptic to them.
In the majority of our experiments, DCMD neurons of both solitarious and gregarious locusts showed a clear 24-h pattern of activity with response curves that were congruent over most of the subjective day. A clear difference in the timing of the peak response was due not just to a simple temporal phase shift but also to superimposed rhythms in responsiveness with higher frequencies. In particular, there was evidence of a 12-h ultradian rhythm in gregarious locusts, both in the relative spike rate (Fig. 4) and in the peak firing time (Suppl. Fig. S2). Both 24-h and 12-h rhythms occur together in other arthropods such as Norway lobster (Nephrops norvegicus) (Aguzzi et al., 2004) and Antarctic krill (Euphausia superb) (Gaten et al., 2008). In krill, the expression of a secondary 12-h rhythm is thought to be enhanced when food supply is limited (Godlewska, 1996) or when normal intraspecific schooling interactions are prevented (Gaten et al., 2008), whereas in our experiments the difference was induced by different rearing densities. Ultradian rhythms with periods of a few hours have received little attention in insects. There is a clear rhythm of feeding with a period of approximately 15 min in locusts (Simpson, 1981), but whether this is linked to the circadian clock or to other independent physiological processes is not known.
In Drosophila, an intraspecific olfactory cue acts to synchronize circadian rhythms, so that flies reared in groups tend to show more highly synchronized rhythms than those reared alone (Levine et al., 2002a). In the gregarious cockroach Leucophaea maderae, however, social interactions do not entrain circadian rhythms of behavior (Knadler and Page, 2009). In contrast to both these studies, our data suggest that crowding (gregarization) of locusts increases the complexity of a circadian pattern of neuronal activity with a consequent advance in the time of subjective day at which DCMD is maximally responsive. One possibility is that the higher frequency (ultradian) rhythms are under the control of clock neurons, the activity of which is enhanced by gregarization. In other words, density-dependent polyphenism might alter the relative levels of activity and coupling between groups of clock neurons to modify visual function alongside the behavioral changes in patterns of flight activity in solitarious and gregarious individuals. Whether the activity and neurochemistry of pigment dispersing factor (PDF)-containing neurons of the central clock circuits (Homberg et al., 1991; Helfrich-Förster, 2003; Park et al., 2000; Lee et al., 2009; Grima et al., 2004; Stoleru et al., 2004) differ in solitarious and gregarious locusts is not known, but these cells provide an obvious focus for further study.
The earlier rise in responsiveness of DCMD seen in gregarious locusts could be due to entrainment by social interaction. Social behavior can influence the functioning of circadian clocks in both Drosophila (Levine et al., 2002a) and honey bees (Toma et al., 2000), probably due to chemosensory effects resetting the clock. Interactions between locusts in the crowded cages used to house gregarious animals in our experiments may therefore have contributed to changes in the diurnal pattern of DCMD responsiveness that persisted as after-effects in our recordings. After-effects are slowly decaying changes in the period of an oscillator following a change from a fixed photoperiod to constant conditions (Pittendrigh and Daan, 1976). Although the circadian clock has a genetically determined state (probably as described here for solitarious locusts), it is also strongly influenced by the conditions in which the animals were maintained prior to the experiment. This could be addressed in future work by retaining the locusts in constant light for 1 day before repeating the experiments described here.
The precision with which the period of a circadian rhythm can be estimated improves as the number of observed cycles increases (Chui, 1992; Levine et al., 2002b). Our extracellular recordings of DCMD permitted sampling over at most 36 h, so we cannot demonstrate directly that the patterns we observed repeat on subsequent subjective days. Despite this, the consistent sinusoidal nature of the data evident in recordings started at different times of day is implicit evidence that the 24-h patterns we observed reflect a persistent circadian rhythm in DCMD responsiveness. Although the strength of the DCMD response to a looming object was modulated in a circadian rhythm, the time of peak firing (i.e., the point during a single looming approach at which the DCMD firing rate reached its maximum) did not vary in a circadian fashion, providing both solitarious and gregarious locusts with a consistent cue to initiate avoidance behaviors (Gabbiani et al., 1999).
Our work demonstrates that there is a circadian rhythm in the response of a descending visual interneuron and that this rhythm is modified during density-dependent polyphenism. The changes in response to imminent collision mirror a key behavioral difference in the timing of flight activity in solitarious and gregarious locusts, thus providing a neural correlate of the reported behavioral plasticity.
Footnotes
Acknowledgements
This work was funded by BBSRC research grants to T.M. and E.G. We thank Drs. I.A. Barber and M.L. Head for discussions about the data, and Dr. S.M. Rogers, Dr. E. Rosato, and Professor C.P. Kyriacou for commenting on a draft of the manuscript.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.