
Abstract
We studied locomotor activity rhythms of C57/Bl6 mice under a chronic jet lag (CJL) protocol (ChrA6/2), which consisted of 6-hour phase advances of the light-dark schedule (LD) every 2 days. Through periodogram analysis, we found 2 components of the activity rhythm: a short-period component (21.01 ± 0.04 h) that was entrained by the LD schedule and a long-period component (24.68 ± 0.26 h). We developed a mathematical model comprising 2 coupled circadian oscillators that was tested experimentally with different CJL schedules. Our simulations suggested that under CJL, the system behaves as if it were under a zeitgeber with a period determined by (24 – [phase shift size/days between shifts]). Desynchronization within the system arises according to whether this effective zeitgeber is inside or outside the range of entrainment of the oscillators. In this sense, ChrA6/2 is interpreted as a (24 − 6/2 = 21 h) zeitgeber, and simulations predicted the behavior of mice under other CJL schedules with an effective 21-hour zeitgeber. Animals studied under an asymmetric T = 21 h zeitgeber (carried out by a 3-hour shortening of every dark phase) showed 2 activity components as observed under ChrA6/2: an entrained short-period (21.01 ± 0.03 h) and a long-period component (23.93 ± 0.31 h). Internal desynchronization was lost when mice were subjected to 9-hour advances every 3 days, a possibility also contemplated by the simulations. Simulations also predicted that desynchronization should be less prevalent under delaying than under advancing CJL. Indeed, most mice subjected to 6-hour delay shifts every 2 days (an effective 27-hour zeitgeber) displayed a single entrained activity component (26.92 ± 0.11 h). Our results demonstrate that the disruption provoked by CJL schedules is not dependent on the phase-shift magnitude or the frequency of the shifts separately but on the combination of both, through its ratio and additionally on their absolute values. In this study, we present a novel model of forced desynchronization in mice under a specific CJL schedule; in addition, our model provides theoretical tools for the evaluation of circadian disruption under CJL conditions that are currently used in circadian research.
Keywords
In mammals, daily rhythms are driven by a circadian clock located in the suprachiasmatic nuclei (SCN) of the hypothalamus. Synchronization of the clock with the external environment is achieved mainly by photic information provided by the daily light-dark (LD) cycle (Golombek and Rosenstein, 2010). This process involves both period and phase locking (i.e., entrainment) of the oscillator to the LD cycle in order to compensate the difference between the zeitgeber and the circadian endogenous period, T and τ, respectively (Aschoff, 1981).
Alterations in zeitgeber period can generate major disruptions in behavioral and physiological functions. For instance, it was recently shown that the exposure of mice to a 20-hour LD cycle, a condition that does not allow stable circadian entrainment, negatively affects metabolic variables, behavior, and brain physiology (Karatsoreos et al., 2011). In addition, re-entrainment to a phase-shifted LD cycle is accompanied by alterations in the internal synchrony of the circadian system at distinct levels of biological organization. Transient desynchrony occurs between SCN and peripheral circadian oscillators (Davidson et al., 2009; Yamazaki et al., 2000), among neuronal subpopulations within the SCN (Albus et al., 2005; Liu and Borjigin, 2005; Nagano et al., 2003), at the neuronal level in the SCN between its molecular core and electrophysiological rhythmic outputs (Vansteensel et al., 2003), and finally, at the molecular level between different SCN clock protein cycling levels (Reddy et al., 2002). Indeed, there are severe consequences of subjecting an organism to repeated phase shifts, such as those experienced by frequent fliers or rotating shift workers. An experimental chronic jet lag (CJL) experience affects complex phenomena such as the immune system (Castanon-Cervantes et al., 2010), tumor progression (Filipski et al., 2004; Filipski and Levi, 2009), or even life expectancy (Davidson et al., 2006) in animal models. We have designed experimental protocols to study recurring phase shifts based on a systematically repeating LD-shifting schedule, which provide a controlled schedule in which the effects of phase shift and intershift interval can be manipulated.
Materials and Methods
Animals
Male C57/Bl6 mice (n = 91; 3-5 months) from the National University of La Plata, Argentina, were used in the study. Animals were housed individually in cages equipped with infrared motion detectors, with food and water ad libitum. The experimental protocols for this study were approved by the local Ethics Committee according to the National Institutes of Health (NIH) Guidelines for the Care and Use of Laboratory Animals.
Behavioral Studies
General locomotor activity was recorded through infrared motion detectors (Archron, Buenos Aires, Argentina), and counts were stored automatically every 5 minutes. All animals were kept under a 12-hour:12-hour LD schedule (300 lux at cage level) until stable entrainment was verified before entering any experi mental procedure. For chronic advancing experimental jet lag (ChrA6/2), 53 mice were subjected to a schedule of 6-hour advances of the LD cycle every 2 days. This was accomplished through a 6-hour shortening of every second dark phase. Thirty of the animals were kept under this ChrA6/2 schedule for 45 to 60 days. The remaining 23 mice were kept under ChrA6/2 for 25 to 30 days, after which they were released into constant darkness (DD). In another experiment, 10 mice were subjected to an asymmetrical 21-hour LD cycle (T21; 12 hours:9 hours L:D) for 30 to 35 days before being released into DD. In a third experiment, 9 mice were subjected to a schedule of 9-hour advances of the LD cycle every 3 days (ChrA9/3) before being released into DD. This was accomplished through a 9-hour shortening of every third dark phase. For the last experiment, 10 mice were subjected to a chronic delay jet lag (ChrD), with 6-hour delays in the LD cycle every 2 days, through a 6-hour lengthening of every second light phase for 25 to 30 days, after which animals were released into DD. All advancing CJL experiments were designed so that photophase in all of them remained equal (12 hours). A group of 9 littermate control mice was used to establish the endogenous period of locomotor activity rhythms of the C57/BL6 strain of mice in DD under our experimental conditions.
Behavioral Data Analysis
To determine the significant periods of the general activity components found under each LD condition, Bonferroni-corrected Sokolove-Bushell (SB) (Sokolove and Bushell, 1978) periodograms were calculated for each animal during the schedules, covering a range from 20 to 27 hours (for the advancing schedules) or from 20 to 28 hours (for the delay schedule). This method allowed us to detect different rhythmic components that either had a period equal to that of the LD schedule (under ChrA6/2, T21, and ChrD) or were longer than 24 hours (under ChrA6/2, ChrA9/3, and T21).
To evaluate the phase locking of free-running activity rhythms to the activity components detected under each schedule, we calculated the phase relation ships between the free-running rhythm and these components. To check whether the LD cycle had any effect on the outcome of the phase in DD, we visually extrapolated the free-running rhythm onset to the last LD cycle for all animals and calculated the phase difference to the last ZT12. To determine the influence of the nonentrained components on the DD phase, we visually extrapolated the center of the activity interval to the last LD cycle and calculated the difference between this time point and the extrapolated onset in DD (Cambras et al., 2004). Finally, the phase differences were entered into Rayleigh z tests (Batschelet, 1981), and clustering was evaluated.
SB periodograms, double-plot actograms, and Rayleigh z tests were performed and built with El Temps software (A. Díez-Noguera, Barcelona, Spain). Statistical analysis of results was performed with GraphPad Prism software (La Jolla, CA). Data are presented as mean ± standard deviation.
Computer Simulations
Simulations were performed using 1 or 2 coupled Pittendrigh-Pavlidis equations. In these equations, R and S are state variables, and a, b, c, and d are oscillator parameters. Zeitgeber L is represented by square waves that are set at zero except for intervals of duration Ldur, when they are set to a fixed amplitude value. These equations differ from the original Pavlidis equation (Pavlidis, 1967) by a parameter K, which is a small nonlinear term (K = 1/[1 + 100R2]) that tends to prevent the R variable from approaching zero. In 2 coupled Pittendrigh-Pavlidis oscillator systems, parameters C12 and C21 correspond to the coupling strengths of oscillator 1 on 2 and of oscillator 2 on 1, respectively. Oscillators affect each other continuously through the C parameter. At each time unit, the value of oscillator 2 affects the rate of change of oscillator 1 (and vice versa) through this coupling coefficient that feeds each oscillator with the value of the other.
L represents lights-on and, when “on”, instantaneously decreases the level of one oscillator parameter, which in turn decreases the level of the amplitude of the oscillator. L is programmed to be “on” and “off” according to the simulated schedule. In this model, light does not affect coupling directly.
Oscillator equations:
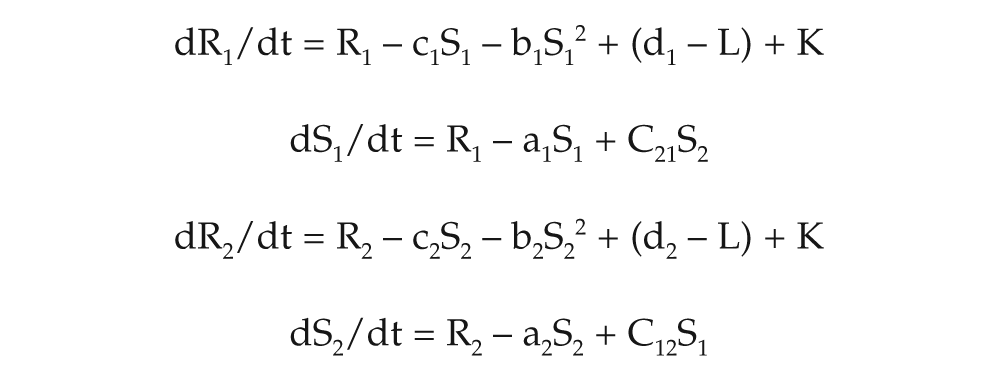
We used the Euler method for numerical integration, with 1000 integration steps per 24-hour day. Variable R was explicitly constrained from achieving negative values. Locomotor activity occurred every time the S variable in either oscillator 1 or 2 rose above some threshold value, which we set to two thirds of the maximum amplitude of this variable (Oda et al., 2000).
Simulations were performed using the Circadian Dynamix software, which is an extension of NeuroDynamix II (Friesen and Friesen, 2009). NeuroDynamix II is freely available for download at http://www.neurodynamix.net. The specific CircadianDynamix model is available free of charge by contacting W.O. Friesen.
Periods for the components found in the simulations were determined by χ2 periodograms built with the ClockLab software (Actimetrics, Wilmette, IL).
Results
Experimental Data 1: Chronic 6-Hour Advances Every 2 Days
Under ChrA6/2, we found that a mean of 58% of mice in each experiment displayed 2 activity rhythms as determined by both actogram observation and periodogram analysis: a short-period component (21.01 ± 0.04 h), following the LD schedule, and a second component with a period greater than 24 hours (24.68 ± 0.26 h) (Fig. 1A and 1B). Animals were considered to be internally desynchronized only when these 2 components, and only these, were detected in the SB periodogram analysis.
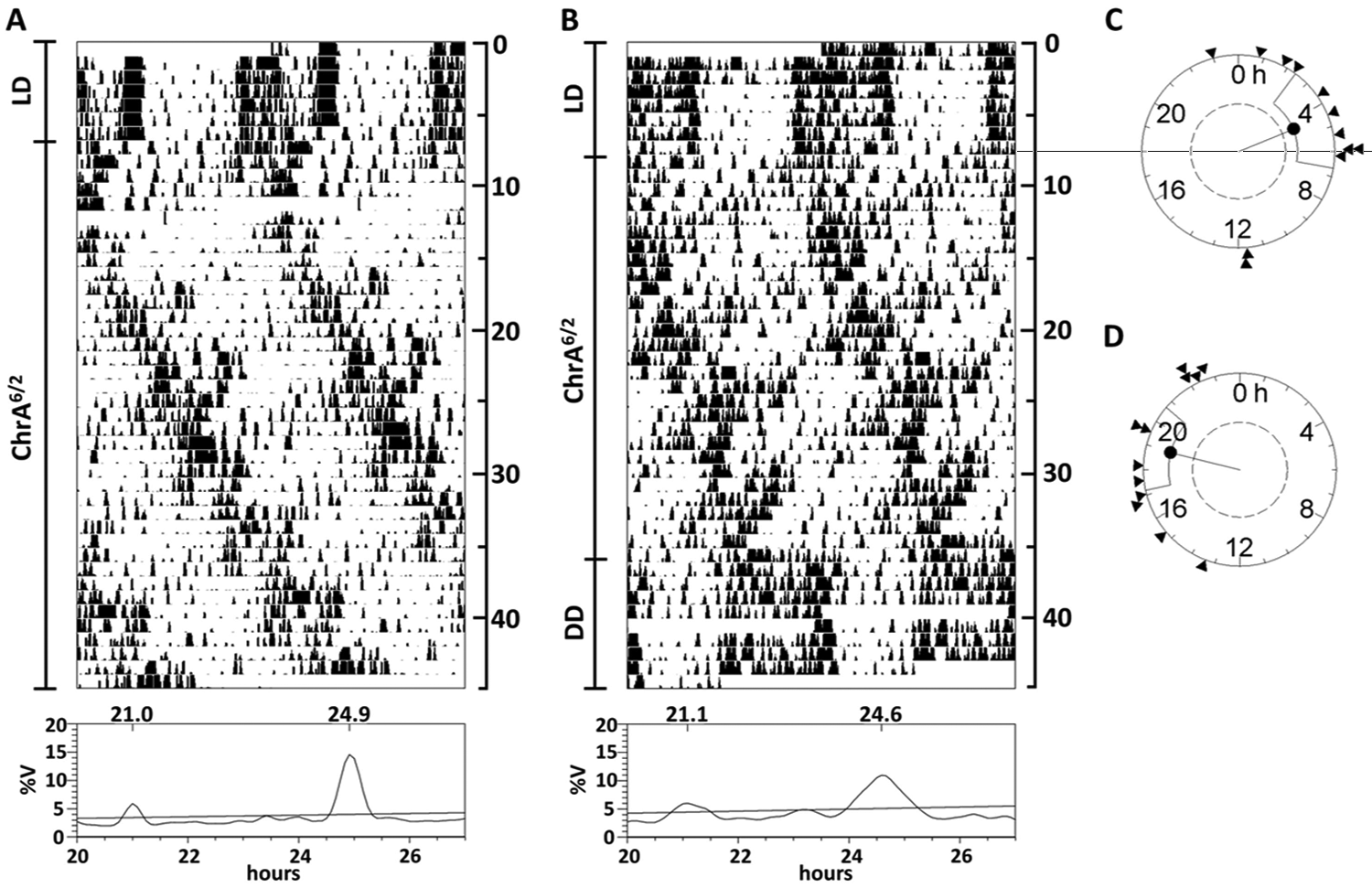
A schedule of 6-hour advances of the LD cycle every 2 days (ChrA6/2) gives rise to 2 components of locomotor activity rhythm in mice. (A, B) Representative double-plot actograms plotted at modulo 24 hours, and Sokolove-Bushell periodograms of animals under the ChrA6/2 schedule. (A) An animal showing 2 activity rhythms, both maintained for 40+ days under ChrA6/2. (B) An animal showing 2 rhythms of activity under ChrA6/2 and a single fused circadian rhythm after being released into DD. Days of recording are shown at the right of each actogram. Significance threshold in periodograms is set to p = 0.05, after Bonferroni correction. %V = percentage of variance. (C, D) Rayleigh z test analysis of the onset phase differences between the DD free-running activity rhythm and either (C) the previous LD cycle or (D) the nonentrained component for animals displaying the described 2 rhythms of activity under ChrA6/2. Clustering is significant for both tests.
The animals that did not meet the above criterion fell into one of these categories (mean percentages along the different experiments and periods of the rhythmic components are indicated): 3 components (11%; 21.03 ± 0.04 h, 23.38 ± 0.15 h, and 24.97 ± 0.37 h), only the short component (11%; 21.01 ± 0.03 h), a single long component (17%; 24.86 ± 0.22 h), and 2 components different from the described (3%; 21.13 ± 0.18 h and 23.42 ± 0.00 h). Nevertheless, desynchronization of the 2 activity components described before was the only pattern present in all the different experiments. When desynchronized mice were released into DD, both components rapidly fused, and the free-running activity rhythm had a period of 23.89 ± 0.38 h, not statistically different from that of control mice under DD (Table 1).
Circadian periods of general activity detected by Sokolove-Bushell periodogram analysis for each light-dark condition tested.
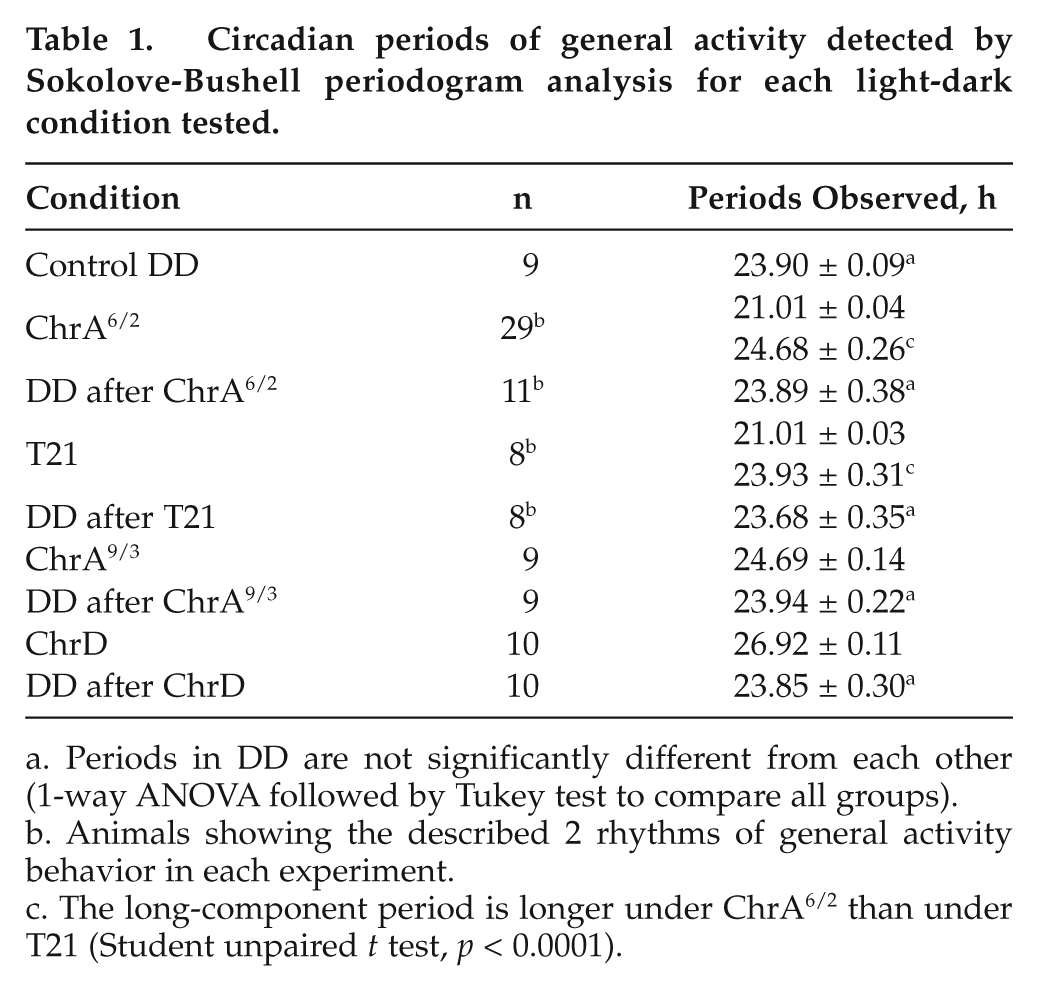
Periods in DD are not significantly different from each other (1-way ANOVA followed by Tukey test to compare all groups).
Animals showing the described 2 rhythms of general activity behavior in each experiment.
The long-component period is longer under ChrA6/2 than under T21 (Student unpaired t test, p < 0.0001).
The onset phase in DD was found to be predicted by both the previous LD cycle and the long-period component, as significant clustering of phase differences was detected in the Rayleigh z tests (Fig. 1C). Based on these findings, we will designate the short-period component as a light-entrained component (LEC) and the long-period component as the non–light-entrained component (NLEC), as described previously for T22-forced desynchronization in rats (Campuzano et al., 1998).
Modeling Section I: Interpretation of Chronic Phase Shifting of 2 Coupled Oscillators
A chronically phase-shifted 24-hour zeitgeber resembles a new zeitgeber with a different, well-defined period (Suppl. Fig. S1A). In the supplementary material, we present model simulations in which the dynamics of a single circadian oscillator under CJL schedules is studied. Basically, it is shown that repeatedly phase-shifted 24-hour LD cycles (i.e., CJL schedules) are processed by the model circadian oscillator as new zeitgebers with periods that differ from 24 hours. The oscillator can either entrain or display relative coordination based on whether this emerging zeitgeber period is within or outside its range of entrainment (Suppl. Fig. S1B and S1C). The effective period (T′) of the emergent zeitgeber will be given by
T′ = 24 – step,
where step results from the division of the size or amplitude of the phase shift (in hours; PS) divided by the number of days between the phase shifts, the intershift interval (in days; ISI); these will be designated as the step components. The unit for step is h, and it is positive for advances and negative for delays. Then, T′ = 24 – PS/ISI.
According to the model, ChrA6/2 is a CJL schedule with a step:
ChrA6/2: 6/2 = +3 h
and hence is interpreted by the circadian system as an effective zeitgeber with a period equal to 24 − 3 = 21 h.
Our activity data of mice under the ChrA6/2schedule reveal an emergent pattern of forced desynchronization (Campuzano et al., 1998; de la Iglesia et al., 2004; Schwartz et al., 2009). This pattern occurs when a system of dual, weakly coupled oscillators is subjected to a zeitgeber that is outside the range of entrainment of one of the oscillators only. We performed simulations of the behavior of a system of 2 coupled oscillators (oscillator 1: τ1 = 23.5 h; oscillator 2: τ2 = 24.5 h) under CJL schedules according to the previous considerations. Symmetrical coupling parameters were used in the simulations because forced desynchronization models are robust enough to be later generalized to asymmetrically coupled systems (Schwartz et al., 2009).
The following schedules with step = +3 were considered: +3/1, +9/3, and +12/4. Under the +3/1 and +9/3 step schedules (and also under +6/2), oscillators dissociate and hence give rise to desynch ronized patterns (Fig. 2). These patterns varied systematically with the step components: the average period of the NLEC component becomes closer to T′ = 21 h under step +3/1 than under step +9/3. Now under step +12/4, the 2 oscillators resynchronize and display relative coordination with an intermediate average period.

Simulation of coupled circadian oscillators subjected to chronic advancing jet lag schedules with steps +3/1, +9/3, and +12/4. Weakly coupled oscillators (τ1 = 23.5 h, τ2 = 24.5 h) desynchronize for steps +3/1 and +9/3 while presenting relative coordination without dissociation for step +12/4. The nonentrained component periods under +3/1 and +9/3 are 25.0 h and 25.2 h, respectively. Oscillator parameters: a1 = 0.85; b1 = 0.3; c1 = 1.0; d1 = 0.5; a2 = 0.85; b2 = 0.3; c2 = 0.7; d2 = 0.5; C12 = C21 = 0.02. Zeitgeber parameters: L = 2.0; Ldur = 1.
When relative coordination of oscillator 2 occurs while oscillator 1 remains entrained to the LD schedule, a forced internal desynchrony is generated under CJL. Forced desynchronization occurs when the zeitgeber is inside the range of entrainment of oscillator 1 only (as in the patterns generated under steps 3/1 and 9/3), and the period of oscillator 2 will be closer to the period of the zeitgeber the closer the latter is from the limits of the range of entrainment.
Internal synchronization of a 2-oscillator system results when it is inside or outside the ranges of entrainment of both oscillators (Schwartz et al., 2009). Accordingly, the single nonentrained component pattern generated under step 12/4 does not dissociate because the effective zeitgeber is outside the range of entrainment of both oscillators.
Based on the simulations, the model predicts that under a CJL schedule with a step = +3/1, the occurrence of 2 desynchronized rhythms of activity might be induced in C57/BL6 mice. One component should be entrained to T′ = 21 h, while the nonentrained one should display a period closer to T′ = 21 h than that found under ChrA6/2. On the other direction, under a jet lag schedule with a step = +9/3, our simulations predict that either 2 desynchronized components or a single component in relative coordination (as in the +12/4 simulation in Fig. 2) should arise. In order to test these predictions, we designed 2 behavioral experiments applying the indicated +3/1 and +9/3 steps. We expected that these experiments would also allow us to test the limits of the proposed hypothesis on the adaptation of the circadian system to CJL schedules.
Experimental Data 2: 21-Hour LD Schedule through Daily 3-Hour Advances of Light Onset
Under the asymmetric 21-hour LD cycle (T21), 8 of 10 animals displayed an activity pattern with 2 components as determined by the periodograms: a short-period one (21.01 ± 0.03 h) and a long-period one (23.93 ± 0.31 h) in relative coordination (Fig. 3A). The long-period component was significantly shorter, and therefore closer to T′ = 21 h, than the one found under ChrA6/2 (Student unpaired t test, p < 0.0001), confirming our model prediction. As reported for ChrA6/2, when animals were released into DD, a single free-running rhythm emerged, with a circadian period of 23.68 ± 0.35 h, not different from that of control mice under DD. Rayleigh z test analysis determined that the onset phase of the free-running rhythm in DD was predicted by both the light LD schedule (which confirms the short component found is indeed a LEC) and the component in relative coordination (Fig. 3C and 3D, respectively).
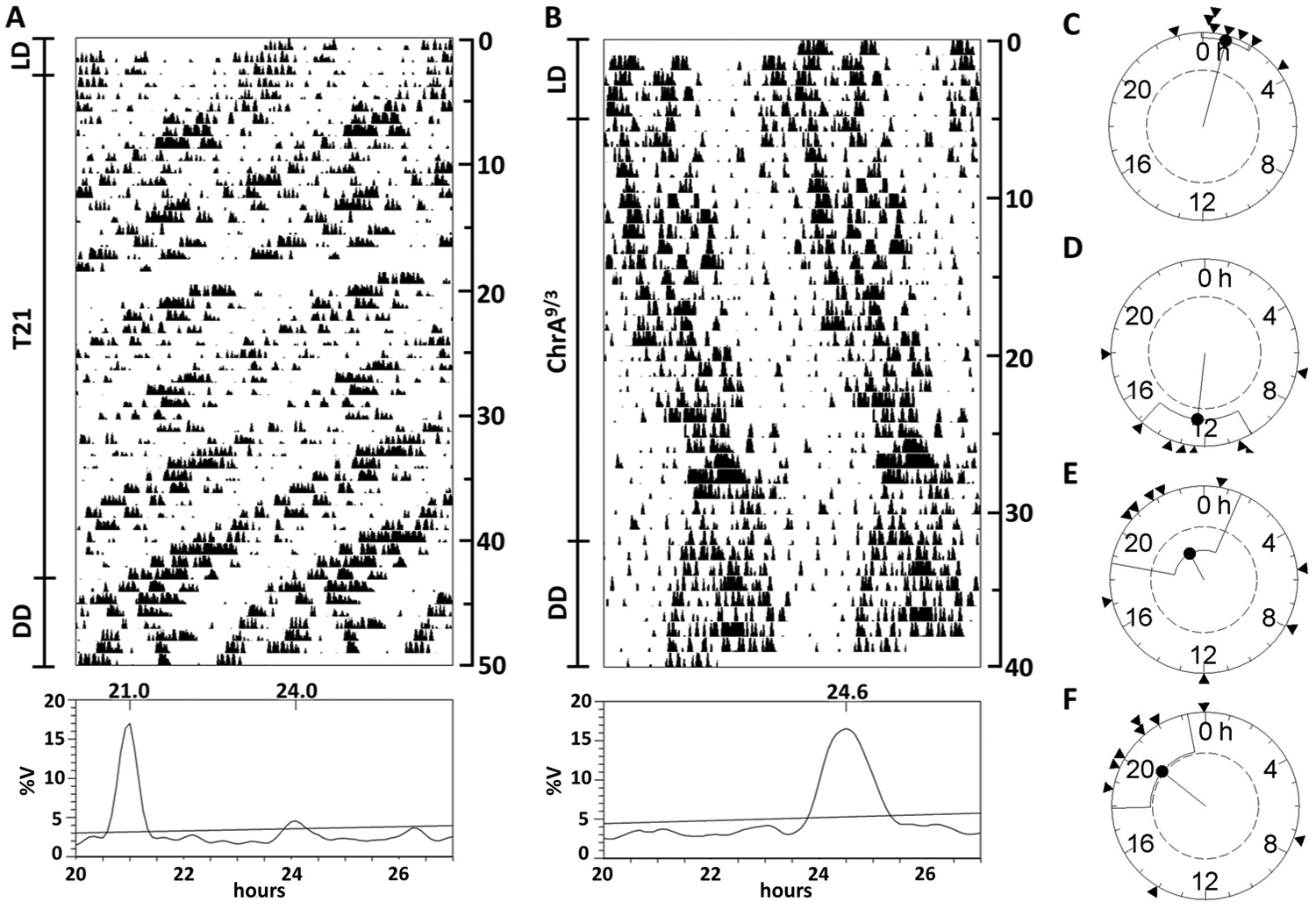
Behavior under different CJL schedules confirms model predictions. (A, B) Representative double-plot actograms plotted at modulo 24 hours, and Sokolove-Bushell periodograms of animals kept under (A) the T21 or (B) the ChrA9/3 schedule and then released into DD. Days of recording are shown at the right of each actogram. Significance threshold in periodograms is set to p = 0.05, after Bonferroni correction. %V = percentage of variance. (C, D) Rayleigh z test analysis of the onset phase differences between the DD free-running activity rhythm and either (C) the previous LD cycle or (D) the nonentrained component for animals displaying the described 2 rhythms of activity under T21. (E, F) Rayleigh z test analysis of the onset phase differences between the DD free-running activity rhythm and either (E) the previous LD cycle or (F) the nonentrained component for animals under the ChrA9/3 schedule. Significant clustering is found in the z tests shown in C, D, and F.
Experimental Data 3: Chronic 9-Hour Advances Every 3 Days
Under the ChrA9/3 schedule, we found no evidence of a light-driven component in the periodograms of all 9 mice tested. An activity component in relative coordination with an average period of 24.69 ± 0.14 h was detected in all animals (Fig. 3B). After release into DD, a free-running rhythm with a 23.94 ± 0.22 h period arose, and its onset was phase locked to the previously described activity component under ChrA9/3 (Fig. 3F), while no clustering due to the LD cycle was detected (Fig. 3E).
Modeling Section II: Asymmetry of Advancing and Delaying CJL
Several species, such as the C57/Bl6 mice used in this study, have phase response curves with larger delay than advance areas (Pittendrigh and Daan, 1976; Schwartz and Zimmerman, 1990). Accordingly, the ranges of entrainment of such species are asymmetrical to each side of the 24-hour period, as also shown by simulations in Supplementary Figure S2. To uncover the consequences of this asymmetry for our CJL model, we applied the model of 2 coupled oscillators to the case of a chronic delay jet lag schedule, with step = −6/2, corresponding to a zeitgeber with period T′ = 27 h. Importantly, each emergent zeitgeber (27 hours for phase delays and 21 hours in the case of phase advances) and oscillator circadian periods (see below) is symmetrical with respect to 24 hours.
Comparison between the dynamics of coupled oscillators 1 and 2 (τ1 = 23.5 h and τ2 = 24.5 h) under CJL schedules with the steps −3 and +3 is shown in Figure 4. While advancing CJL (upper traces) induced forced desynchronization, there was entrainment of the coupled system under the delaying CJL schedule (lower traces).
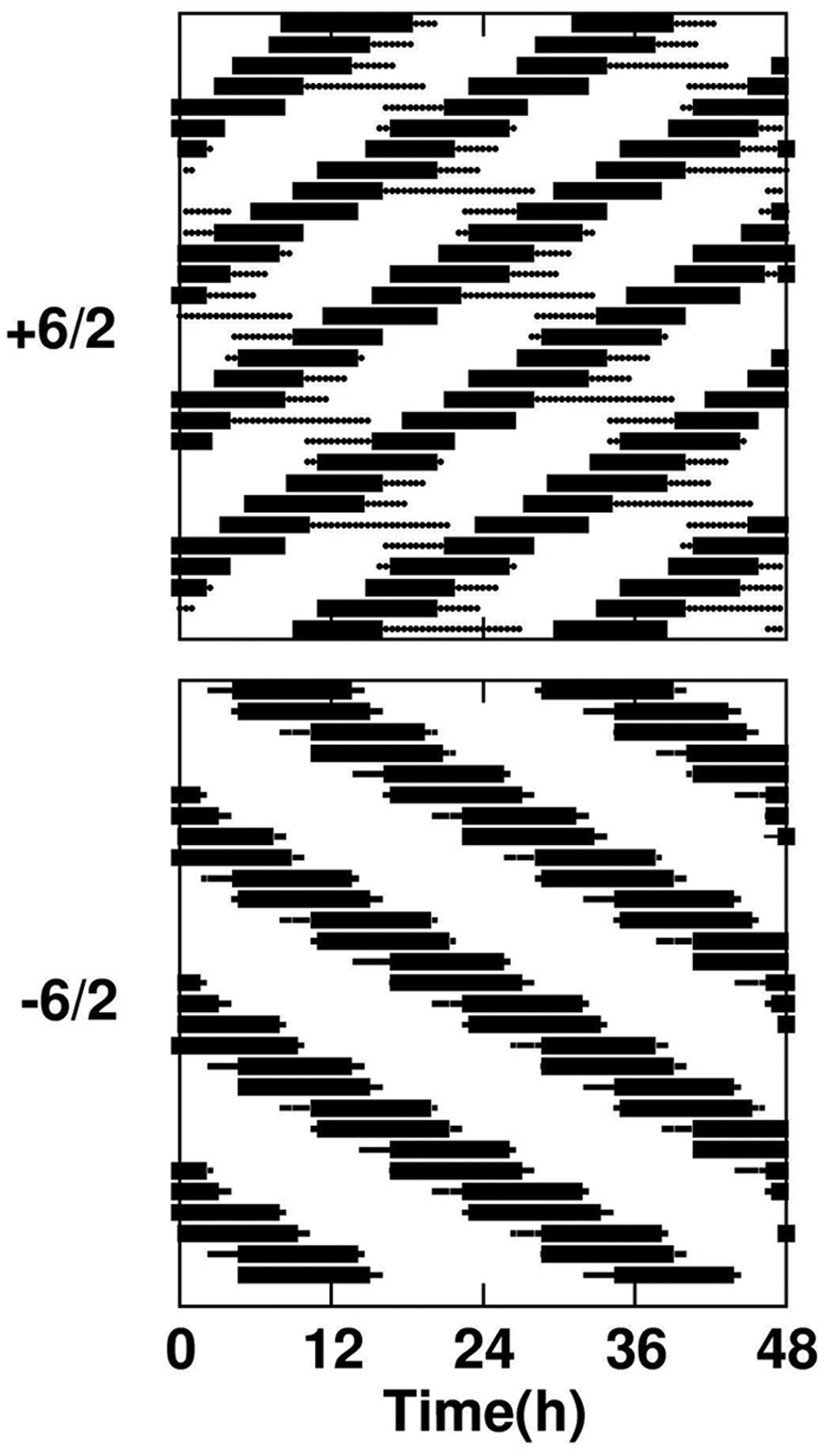
Comparative simulations of oscillators under ChrA6/2 and ChrD. Simulation of 2 coupled circadian oscillators (τ1 = 23.5 h, τ2 = 24.5 h) subjected to CJL schedules with steps equal to +6/2 and −6/2. No desynchronization, but correct entrainment of the system, is found under −6/2. The nonentrained component period under +6/2 is 25.2 h. Oscillator parameters: a1 = 0.85; b1 = 0.3; c1 = 1.0; d1 = 0.5; a2 = 0.85; b2 = 0.3; c2 = 0.7; d2 = 0.5; C12 = C21 = 0.02. Zeitgeber parameters: L = 2; Ldur = 1.
Besides the larger delays portion of the phase response curve, another feature of our model oscillator system that contributes to the asymmetry between advancing and delaying schedules is the stronger coupling that the longer period usually oscillator, with its larger amplitude (in limit-cycle oscillator models, an increase in the period usually implies an increase in amplitude), exerts on the shorter period component. In our model, although the coupling constants are the same, they are multiplied by the amplitudes of each oscillator, thus generating stronger coupling of oscillator 2 with respect to 1.
The above arguments predict that under a delaying CJL schedule symmetrical to the previously studied ChrA6/2, desynchronization would be unlikely, and animals might be able to entrain to this 27-hour emergent zeitgeber.
Experimental Data 4: Chronic 6-Hour Delays Every 2 Days
All 10 animals under the ChrD schedule displayed a dominant long-period rhythm consistent with the 27-hour predicted emergent zeitgeber (26.92 ± 0.11 h) (Fig. 5A). The activity patterns of 5 mice also exhibited a minor peak in the periodogram (at 25.13 ± 0.36 h), attributable to infrequent relative coordination events, but no component was evident in the actograms (not shown). After release into DD, a free-running rhythm with a period of 23.85 ± 0.30 h was detected. When the onset phase was analyzed in the animals with a single activity component by means of a Rayleigh z test, there was a significant clustering of phase values relative to the LD cycle, demonstrating that these animals were entrained by the ChrD schedule (Fig. 5B). The phase angle of the onset under DD was found to be advanced with respect to the light offset, as would be predicted by the τ – T difference. Hence, we considered the activity of these mice to be entrained by the light cycle.
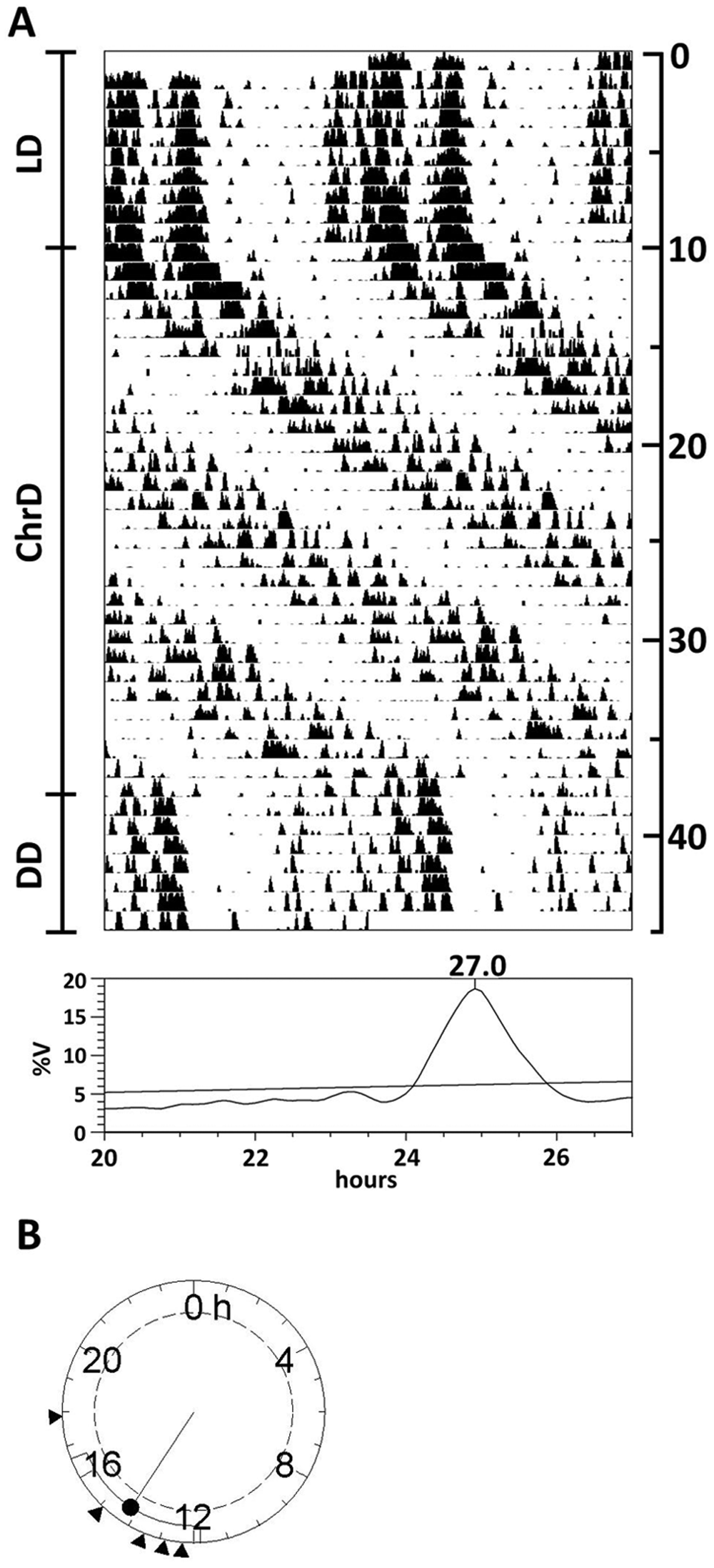
A schedule consisting of 6-hour delays every 2 days of the LD cycle does not lead to desynchronization of locomotor activity rhythms. (A) Representative double-plot actogram plotted at modulo 24 hours, and Sokolove-Bushell periodogram of an animal under the ChrD schedule. Days of recording are shown at the right. Significance threshold in periodograms is set to p = 0.05, after Bonferroni correction. %V = percentage of variance. (B) Rayleigh z test analysis of the onset phase differences between the DD free-running activity rhythm and the previous LD cycle.
Discussion
Experimental CJL protocols provide a controlled LD schedule in which phase-shift size and sign (advance or delay) and interval between shifts can be manipulated in the laboratory. By recording activity under a CJL schedule for 40+ days, our behavioral data reveal an emergent pattern of forced desynchronization, which suggests a new perspective for the systematic evaluation of different CJL schedules. The resulting disturbances in the circadian timing system are better understood by the overall effects of repeated phase shifts and not by the sum of individual disturbing effects of each phase shift.
The experimental protocols designed here test the ability of the circadian system of C57/Bl6 mice to entrain to specific LD conditions. The ChrA6/2 protocol produces a forced desynchronization of locomotor activity into 2 components in most of the animals. One component shows an approximately 21-hour period, which follows the ChrA6/2 schedule, with a stable phase and which is able to predict the phase of the activity rhythm after release into DD. Also, the onset phase in DD had a delayed phase angle as related to the dark phase onset, a fact that could be predicted from the τ – T difference between the biological oscillator and the emergent zeitgeber. The other component exhibits relative coordination with an average period longer than 24 hours and also predicts the phase of activity onset in the subsequent release into DD. We conclude, as it was also observed in the T22-rat forced desynchronization model (Cambras et al., 2004), that both circadian components are reliable markers of desynchronized oscillator components. We found a very similar pattern under T21, with both an entrained and a nonentrained component. The nonentrained component under T21 displayed a shorter period than the one found under ChrA6/2, a difference that was essentially predicted by the simulations.
Our simulations unveil several layers of the disruption of the circadian system under CJL regimes. The main cause of this disruption seems to be sustained desynchrony, in contrast to the transient desynchrony that characterizes jet lag. This sustained desynchrony can occur between zeitgeber and oscillator and also between component oscillators, giving rise to internal desynchrony. In a system of strongly coupled oscillators, the CJL schedules cause desynchrony between the LD schedule and the circadian oscillator, when light schedules give rise to an emergent zeitgeber whose period is outside the range of entrainment of the system as a whole (as in route A in Suppl. Fig. S2). Importantly, our simulations indicate that this desynchrony is not due to the phase-shift size or to the interval between shifts in the light cycle alone but to their combined effects. Moreover, in a weakly coupled multioscillator system, CJL schedules elicit internal desynchronization. In this case, some components may undergo entrainment to the LD schedule, while others display relative coordination. When desynchronization occurs, the nature of the step components is very important: the larger such components are, the higher the values for the nonentrained period (Fig. 2). This is observed in the CJL schedules studied here (i.e., T21 v. ChrA6/2), which were designed with the same photoperiod. Thus, a parametric effect of light cannot explain this difference, as it was reported in forced desynch ronized rats under T22 cycles (Cambras et al., 2004). A counterfactual, nonparametric hypothesis, based on the relevance of the LD transitions present in the CJL schedules, should be constructed in further studies.
The first interpretation of this modulation in desynchrony patterns is that increasing step components are effectively processed by the system as a decreasing amplitude of the emergent zeitgeber in a CJL schedule (as in route B in Suppl. Fig. S2). This is in accordance with the observed output of the coupled system to lose the entrained component when step components were increased. Entrainment of at least one component oscillator (Fig. 2) is achieved under the schedules with lower step components, suggesting stronger zeitgebers. Experimental results have followed these model interpretations. On one hand, the ChrA6/2 and T21 schedules both showed desynchronized patterns with different NLEC periods (longer under ChrA6/2 than under T21) as predicted. On the other, under the ChrA9/3 schedule, no entrained activity component was detected but a single component in relative coordination, which indicates a weak zeitgeber that falls out of the range of entrainment of each oscillator. While we have chosen a 2 coupled oscillator’s model in the present work, it should be considered that either amplitude or frequency modulation of a single circadian oscillator can also generate activity patterns composed by up to 3 components, as demonstrated with model simulations of behavioral data of rats under T22 (Granada et al., 2011). However, a third long-period component was not observed in any of the periodograms studied here.
A second interpretation is related to the recently reported statement that for a fixed period and amplitude of the zeitgeber, the range of entrainment increases when the relaxation rate of the oscillator is increased (Abraham et al., 2010), that is, how fast it recovers its amplitude after an external perturbation. Comparing 2 CJL schedules with same step and different step components, the lower these compo-nents, there is less time for amplitude recovery of the oscillator after each periodic phase shift of the zeitgeber. This fact could cause, throughout the schedule, an increase in the range of entrainment.
Regarding the limits of our model interpretation of the response of real circadian systems to CJL schedules, some points should be taken into account. We have considered repeated jet lag schedules that induce a permanent state of disrupted entrainment and not episodic re-entrainment, as would happen if the system is able to re-entrain to the new LD phase well before the next phase shift takes place. The model would be limited according to this when intershift intervals are long enough for the system to entrain before the next phase shift occurs. Our experiments testing 3 different combinations of step components (+3/1, +6/2, and +9/3), giving rise to 21-hour emergent zeitgebers, have consistently followed the possible predictions of the model simulations.
Our model shows that the range of entrainment is significantly skewed towards the delaying region, as previously shown and as is predicted by an asymmetric phase response curve. Consequently, delaying jet lag schedules should be less effective in generating internal desynchronization than advancing ones. We found a major periodogram peak equal to the zeitgeber period, corresponding to an entrained component in all of the animals subjected to a CJL schedule with step = −6/2, and no desynchronization of circadian components, confirming this prediction of the model. The lack of desynchrony under delaying schedules might explain the results found by Davidson et al. (2006), who showed that old mice had a shorter life expectancy under a +6/7 step CJL schedule than under one with a −6/7 step. However, it should be mentioned that these schedules were closer to 24-hour entrainment conditions (23.14- and 24.86-hour emerging zeitgeber periods, respectively) than the ones used in our present work.
Arrhythmic variables are more commonly observed when CJL effects are assayed at the molecular level, using other schedule steps. Filipski et al. (2004) have used a schedule with step = +8/2, which corresponds to an emergent period of 20 hours. Based on these data, computer simulations suggested that these arrhythmic data may be quasiperiodic or chaotic (Leloup and Goldbeter, 2008). In forced oscillator systems, there have been reports of transitions among periodic, quasiperiodic, and chaotic dynamics when periods and/or amplitudes of the driving cycles are manipulated (D’Humieres et al., 1982; Ding, 1987; Glass and Mackey, 1988; Mackey and Glass, 1977). Our behavioral data and the ensuing mathematical model indicate that this period and amplitude are defined by each particular CJL schedule. Several CJL steps are shown in Supplementary Figure S3, showing the corresponding period of the CJL schedule according to our simulations. This is useful for evaluating which schedule is potentially more disrupting based solely on the effective period of the schedule, considering that as the period differs more from 24 hours, it is more likely for unstable patterns to appear (Chiesa et al., 2005). Our present work provides a significant approach to the design of appropriate CJL protocols and the evaluation of the disruption associated to them. This becomes more important, as there has been no systematic evaluation of different CJL protocols before, and because of this, most research carried out so far on this subject has been done using steps chosen based on little information on the diverse degrees of disruption they may cause.
Under advancing CJL schedules, a sustained, forced desynchronization of a dual oscillator system may be established at the behavioral and molecular levels. This was first demonstrated using LD cycles outside the range of entrainment (Wever, 1979) and later demonstrated to occur among ventrolateral and dorsomedial neuronal oscillators within the SCN (de la Iglesia et al., 2004). Indeed, multiple components of overt behavioral rhythmicity might arise from changes in coupling among SCN subregions (Aton and Herzog, 2005; Welsh et al., 2010; Albus et al., 2005; Nagano et al., 2003). Also, these effects were shown to be more pronounced and sustained in rats when the LD cycle is advanced as compared to delays, and this correlates with a slower re-entrainment rate (Nakamura et al., 2005). In rats, the forced desynchronization protocol, which uses a 22-hour LD cycle, results in the dissociation of the circadian rhythm into 2 components that are related to the activity of vlSCN and dmSCN, respectively (de la Iglesia et al., 2004). Our murine forced desynchronization model could also be reflected in the differential activity of SCN subregions in this species, a possibility that remains to be tested.
To conclude, we present a novel protocol of forced desynchronization of activity rhythms in a murine model through the ChrA6/2 CJL schedule, which has significant implications for further studies dealing with the circadian clocks and physiology. Chronic desynchronization has profound effects in health and quality of life, and understanding its behavioral and molecular bases will certainly shed light into the elusive mechanism of malfunctioning circadian clocks.
Footnotes
Acknowledgements
This work was supported by grants from the National Science Agency (ANPCyT, Argentina), National Research Council (CONICET, Argentina), National University of Quilmes (Argentina), FAPESP (Brazil), and FIRCA (NIH, USA).
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.