
Abstract
The ability to adapt to different environmental conditions including seasonal changes is a key feature of the circadian clock. Here, we compared the ability of 3 Drosophila melanogaster wild-type strains to adapt rhythmic activity to long photoperiods simulated in the laboratory. Fruit flies are predominantly crepuscular with activity bouts in the morning (M) and evening (E). The M peak follows dawn and the E peak follows dusk when the photoperiod is extended. We show that this ability is restricted to a certain extension of the phase angle between M and E peaks, such that the E peak does not delay beyond a certain phase under long days. We demonstrate that this ability is significantly improved by simulated twilight and that it depends additionally on the genetic background and the ambient temperature. At 20 °C, the laboratory strain CantonS had the most flexible phase angle between M and E peaks, a Northern wild-type strain had an intermediate one, and a Southern wild-type strain had the lowest flexibility. Furthermore, we found that the 3 strains differed in clock light sensitivity, with the CantonS and the Northern strains more light sensitive than the Southern strain. These results are generally in accord with the recently discovered polymorphisms in the timeless gene (tim) that affect clock light sensitivity.
A key task of endogenous clocks is to adapt the activity of animals to different environmental conditions and especially to seasonal changes that are accompanied by different photoperiods. This adaptation can nicely be followed in animals showing bimodal activity patterns: The timing of the morning (M) and evening (E) activities of finches correlates with dawn and dusk, meaning that the M activity occurs earlier and the E activity later under long summer days (Aschoff and Wever, 1965). On the other hand, M and E peaks are closer together under short-day conditions. This seasonal change in phase angle between M and E peaks is also observed in other animals (Aschoff, 1966; Daan et al., 2001; Pittendrigh and Daan, 1976) including the fruit fly Drosophila melanogaster (Majercak et al., 1999; Rieger et al., 2003; Shafer et al., 2004). For flies and other insects, optimal timing of their activity is most important during long and hot summer days when they are most at risk of overheating and desiccation. Indeed, the flies respond to increased temperature and light intensity by prolonging their siesta and by shifting M and E activity peaks into the early morning and late evening, respectively, already at a 12-hour day length (conventional 12-hour:12-hour light:dark cycles) under laboratory conditions (Majercak et al., 1999; Rieger et al., 2007; Yoshii et al., 2009). Under long photoperiods, the increased phase angle between M and E peaks is most evident (Rieger et al., 2003). Nevertheless, the previous studies also showed that this increase is limited, indicating that M and E peaks are mutually coupled and not working independently. In the present study, we intended to reveal whether simulated twilight modifies the adaptation of the flies to long photoperiods and whether the flexibility between M and E activity peaks under long days depends on the genetic background of wild-type strains caught in the field.
In nature, gradual changes in light intensity and spectral composition characterize the light transitions of dawn and dusk. Several studies have shown that the clock’s entrainment characteristics differ if twilight transitions are present (Wever, 1967; Kavaliers and Ross, 1981; Boulos et al., 1996a, 1996b, 2002; Fleissner and Fleissner, 1998, 2002; Gorman and Zucker, 1998; Danilenko et al., 2000; Boulos and Macchi, 2005; Comas and Hut, 2009). In these studies, twilight was usually defined as gradual, natural-like changes of light intensity without modifying the spectral composition of light (exception: Wever, 1967). Nevertheless, this kind of simulated twilight provoked a lower day-to-day variability of activity on-sets and thus a higher precision of the rhythms in many species (Boulos et al., 2002; Boulos and Macchi, 2005; Fleissner et al., 1996; Gorman and Zucker, 1998). In hamsters and mice, simulated twilight additionally changed the phases of the rhythms, and this was correlated with period changes in subsequent constant darkness (DD), suggesting that twilight influences the clock instead of merely provoking masking effects (Boulos and Macchi, 2005; Comas and Hut, 2009).
In fruit flies, simulated twilight (natural-like change of light intensity over 1.5 hours without modifying color) provoked an almost complete shift of the activity into dawn and dusk (Rieger et al., 2007) (Fig. 1B): The M activity was advanced and the E activity delayed in comparison to rectangular light-dark cycles. Interestingly, M and E activities peaked when light reached an intensity of approximately 7.5 lux, independently of the final irradiance during the day (10, 100, 500, or 1000 lux) (Rieger et al., 2007). This result suggested that the flies prefer to be active at rather dim light. The previous study was performed with the laboratory strain CantonS under LD 12:12. Longer photoperiods have not yet been tested with twilight simulation, and it is also unknown whether wild D. melanogaster strains respond in the same way to artificial twilight.
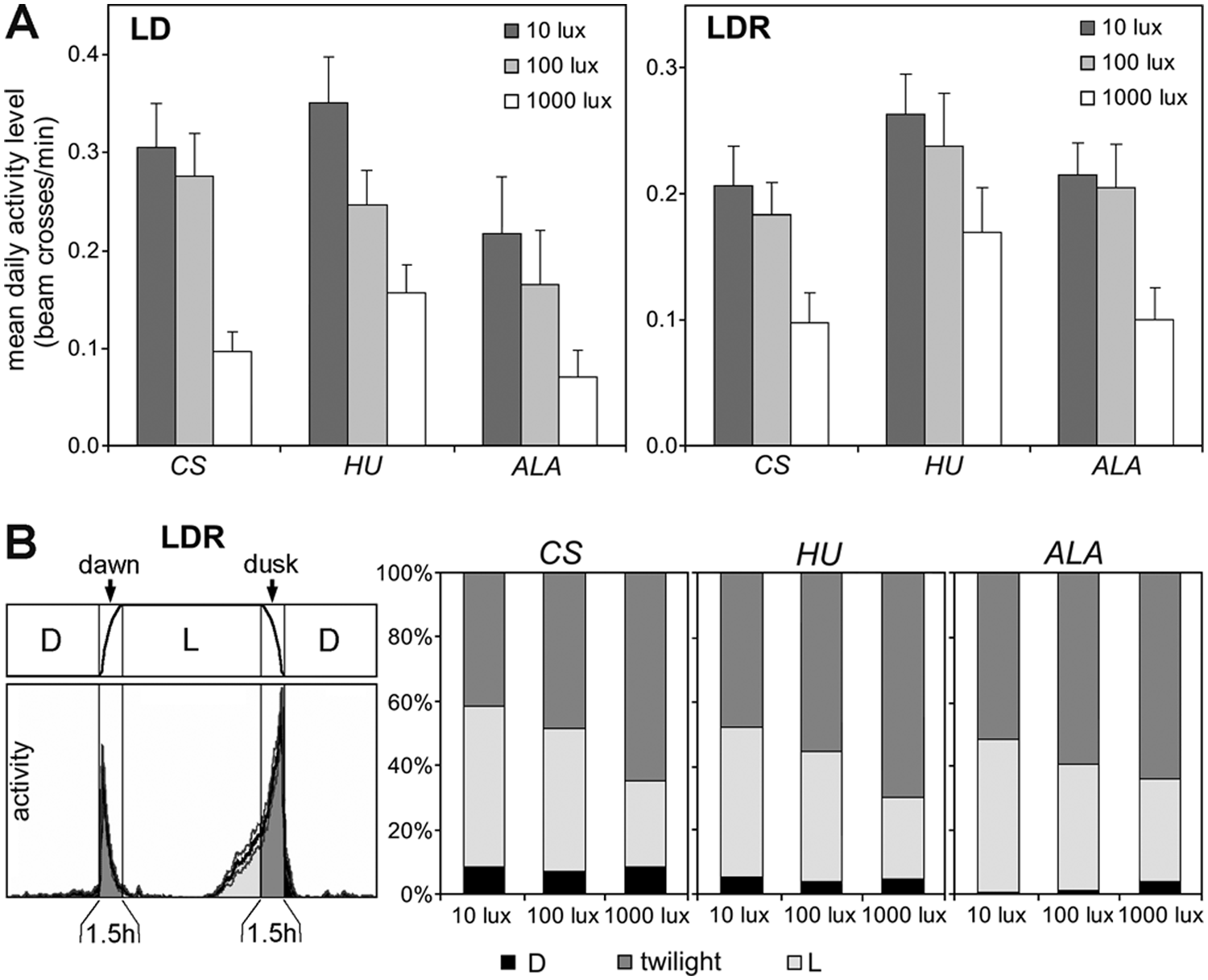
Activity level of CantonS (CS), HU, and ALA flies under rectangular 12:12 LD cycles and 12:12 cycles with simulated twilight (LDR) at 3 different irradiances during the light phase. (A) Mean daily activity level in the 3 strains at 10, 100, and 1000 lux. In all 3 strains, the activity level decreased with increasing irradiance (LD: F2,165 = 9.778; p = 0.0001; LDR: F2,167 = 5.838; p = 0.0035). (B) Under LDR conditions, the flies shifted their activity into dawn and dusk. The left diagram shows the schedule of simulated twilight together with the average activity profile of 30 CantonS flies at 1000 lux. The right diagrams show the percentage of whole day activity each strain spent during twilight (sum of dawn and dusk), the light phase (L), and the dark phase (D) at the different irradiances. ANOVA revealed a significant effect of irradiance on twilight activity (F2,167 = 30.520; p < 0.0001), which was the same in each strain (no interaction between irradiance and strain; F4,167 = 1.211; p = 0.308).
To reveal the influence of the genetic background on the adaptation to 12:12 LD cycles and long photoperiods, we compared the behavior of the laboratory strain CantonS with that of 2 wild-type strains caught in moderate Southern (46°N) and Northern (52°N) Europe. In the South, the longest photoperiod in summer was around 16 hours, and the flies were observed to be active until early November when photoperiods were already shorter than 12 hours (Bhutani, 2009). In the North, active adult flies were exposed to photoperiods up to 18 hours, and they rarely experience photoperiods shorter than 12 hours because they enter diapause in autumn when the temperature drops below a certain value (Bhutani, 2009). We therefore expected the Northern strain to adapt more easily to long photoperiods than the Southern strain.
Polymorphisms in several clock genes appear to facilitate the adaptation to certain environmental conditions, and among those, the short (s) and long (ls) forms of the timeless (tim) gene play a major role in the response to light (Kyriacou et al., 2008). The clock of s-tim flies is more light sensitive than that of ls-tim flies (Sandrelli et al., 2007), but ls-tim flies have a higher incidence to enter diapause, likely increasing the flies’ fitness in the North (Tauber et al., 2007). Therefore, it is believed that the ls-tim allele, which originated in the South and is still more frequent there, will slowly infiltrate the North (Kyriacou et al., 2008).
Here, we show that the simulation of twilight helped all 3 wild-type strains to adapt to more extreme photoperiods. Nevertheless, the maximal phase angle between M and E activity peaks was limited, genetically determined, and only partly dependent on the tim allele.
Materials and Methods
Strains and Rearing
Locomotor activity was recorded in the laboratory strain CantonS that was isolated from a wild strain near Canton, Ohio, in 1930 (Bridges and Brehme, 1944) as well as in 2 wild-type strains that were caught from the wild in Northern and Southern Europe (Sandrelli et al., 2007; Tauber et al., 2007). The Northern wild-type strain stemmed from one isofemale line caught in the Netherlands, close to the town Houten (52°N). The corresponding strain was named HU(4-1). The second strain originated from 37 isofemale lines caught in Val Venosta (Alto-Adige) in North Italy (45°N) and was named ALA. The isogenic ALA lines contributed to a mixed culture, which was regularly refreshed by adding a few flies from the isofemale lines that were maintained separately. The flies were raised on Drosophila medium (0.8% agar, 2.2% sugar-beet syrup, 8.0% malt extract, 1.8% yeast, 1.0% soy flour, 8.0% corn flour, and 0.3% hydroxybenzoic acid) at 20 °C in LD 12:12.
Recording
At the age of 1 to 3 days, individual male flies were transferred into the recording chambers (photometer cuvettes that were placed with one end in an infrared light beam; on the opposite end, the flies had access to water and sugar). Locomotor activity was recorded photoelectrically in 1-minute intervals at 20 °C (or 30 °C) as described (Rieger et al., 2007). Illumination was derived from “white” LEDs (Lumitronix LED-Technik GmbH, Jungingen, Germany) and was adjusted to 100 lux (19 µW/cm2). Either rectangular LD cycles (lights were shut on and off) were applied or dawn and dusk (LDR cycles) was simulated by increasing/decreasing irradiance in 1-minute steps within 1.5 hours (Rieger et al., 2007).
In the first experiment, the activity of the flies was recorded at 20 °C under LD/LDR 12:12 cycles of different light intensity (10, 100, and 1000 lux; 2.4, 19, and 150 µW/cm2, respectively) to judge the response of the flies to increasing irradiances.
In the next experiment, activity was recorded at 20 °C for about 1 week under LD/LDR 12:12 cycles. Subsequently, the duration of the photoperiod was increased symmetrically to 14 hours (or 16 hours). After another week of recording, the photoperiod was further increased (to 18 hours or 20 hours). One experiment was performed under LDR 18:6 cycles at 30 °C.
In a further experiment, the wild-type flies were transferred into DD after initial entrainment to 12-hour:12-hour LD cycles at 20 °C to determine the free-running period of the 3 different strains.
A last experiment was performed to determine the light sensitivity of the clock in the 3 wild-type strains. For this purpose, the flies were entrained to rectangular light-dark cycles of low light intensity at 20 °C. Intensity was adjusted by neutral density filters (Rosco Laboratories Ltd., London, UK). In the first week of recording, light intensity was 6.0 × 10−3 µW/cm2 (~0.003 lux). After all flies had entrained to this intensity, the LD cycle was phase advanced by 6 hours, and simultaneously, light intensity was reduced to 3.6 × 10−4 µW/cm2 (~0.0002 lux, equal to starlight). In the next step, the LD cycle was again phase advanced by 6 hours, and simultaneously, light intensity was reduced to 2.2 × 10−5 µW/cm2 (~0.00001 lux). The last light intensity was at the limit of entrainment, and only few flies could re-entrain (compare Hirsh et al., 2010). Therefore, the experiment was repeated at a light intensity of 4.7 × 10−5 µW/cm2 (~0.00003 lux). After the end of the experiments, the number of days the flies needed to re-entrain for each shift was determined as well as the percentage of flies that were able to re-entrain. All this was judged on the original actograms by 3 different persons who were “blind” to the genotype. These persons were advised to determine the number of days each fly needed until its M and E activity peaks had reached exactly the same phase they had before the phase shift. Determined values were averaged and plotted.
For all experiments and for each condition, 20 to 50 flies were recorded and analyzed.
Data Analysis
The raw data were displayed as actograms using ActogramJ (http://actogramj.neurofly.de) (Schmid et al., in press). Flies were judged to be entrained when they exhibited 24-hour periods, and M and E activity peaks showed stable phase angles to lights-on and lights-off (or to dawn and dusk), respectively. If this was the case, mean activity profiles of individual flies and of groups of flies were calculated for each chosen light intensity and light program as done previously (Helfrich-Förster, 2000; Rieger et al., 2003). The average days shown in the diagrams are smoothed by a moving average filter (over 11 values). To reveal the phase angle of the M and E peaks to lights-on or lights-off, the maxima of both peaks were determined and the phase angle between M and E peaks calculated (Helfrich-Förster, 2000; Helfrich-Förster et al., 2001; Rieger et al., 2003). The maxima of the peaks were determined manually by clicking on them after the data had smoothed over 101 values by a nonrecursive linear digital low-pass filter in the second canonical form (detailed description in Helfrich-Förster, 2000). This filter moved the actual peaks towards their center of gravity. Data were analyzed for statistical differences by a 1- or 2-way ANOVA after testing for normal distribution (Kolmogorov-Smirnov). In case of nonnormal distribution, p values were adjusted by multiplication with 10 according to Glaser (1978).
tim Polymorphism
The timeless gene of the different Drosophila strains was partly sequenced using genomic DNA. To distinguish between s-tim and ls-tim animals, we used the following oligonucleotide primers to amplify (and later to sequence) the genomic DNA: sense TAGGTATCGCCCTCCAAG and antisense TAGGCAGCTCCACAATCA.
Results
Activity Pattern and Activity Level of the 3 Wild-Type Strains under Increasing Irradiances in 12:12 Light-Dark Cycles
Under conventional rectangular LD cycles, CantonS flies reduce overall activity with increasing irradiance and shift parts of activity into the dark phase. Under simulated twilight, they shift most activity into dawn and dusk, and this behavior is more pronounced the higher the irradiance during the light phase. To test whether the Northern and Southern wild strains show the same behavior as the laboratory strain, we recorded their activity pattern under rectangular 12:12 LD cycles and cycles with simulated twilight (12:12 LDR cycles) at irradiances of 10, 100, and 1000 lux during the light phase.
We found that all 3 strains reduced the overall activity level with increasing irradiance during the light phase (Fig. 1). This reduction was very pronounced under LD cycles but also present under LDR cycles. Under the latter, the flies of all 3 strains shifted activity into dawn and dusk, and this tendency increased with increasing irradiance during the light phase (Fig. 1). In the laboratory strain and the Northern HU strain, the peaks of M and E activity occurred at a light intensity of approximately 5 lux, whereas M and E activities peaked slightly later/earlier at approximately 10 to 20 lux in the Southern strain ALA (not shown).
Entrainment of the 3 Wild-Type Strains to Long Photoperiods
In the next experiment, all 3 strains were recorded under long photoperiods of either LD or LDR cycles. We found that all flies of the 3 wild-type strains entrained to the long photoperiods and increased the phase angle between M and E peaks (YM,E) with increasing day length (Fig. 2), but we observed clear differences between rectangular LD cycles and simulated twilight as well as differences between the strains that will be described in the following.
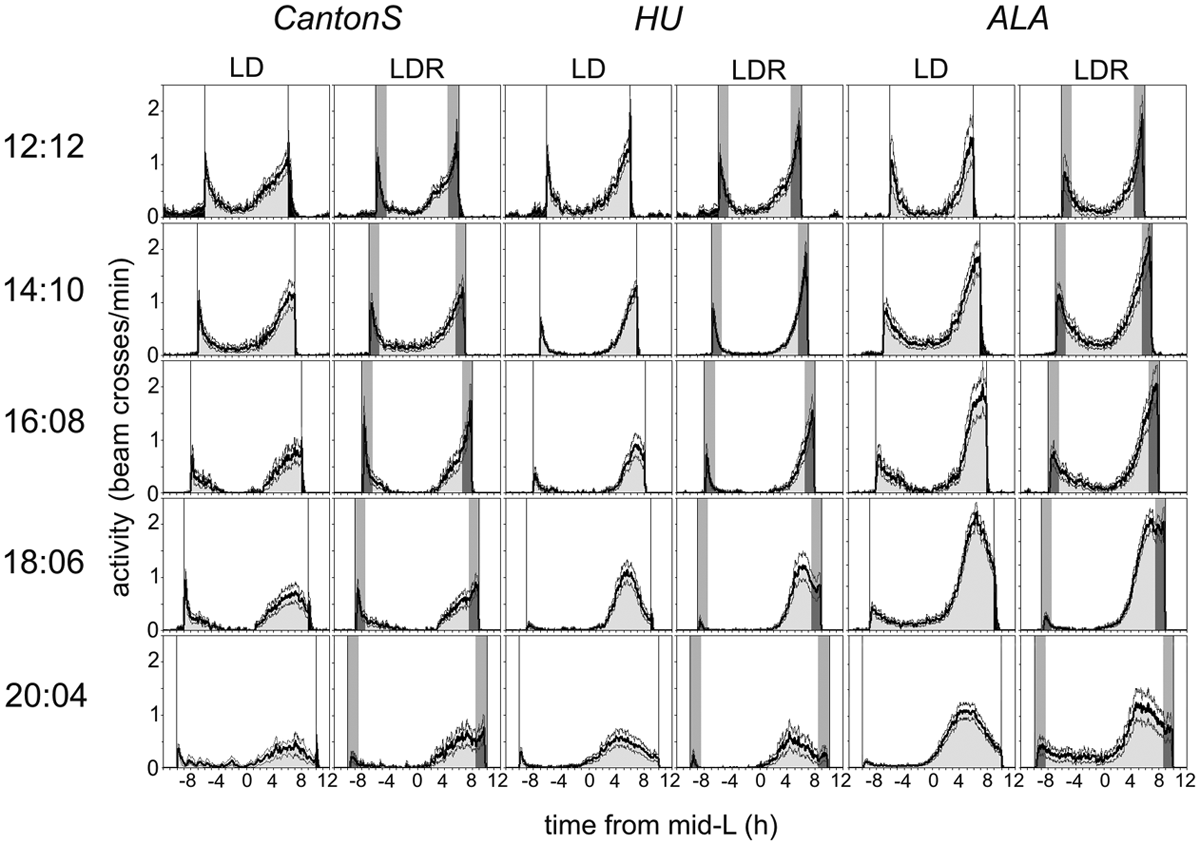
Mean activity profiles (±SEM) of CantonS, HU, and ALA flies under 12-hour:12-hour light-dark cycles and under long photoperiods with and without simulated twilight. Activity during the light phase is indicated in light gray, during dawn and dusk phases in darker gray, and during the dark phase in black. For details, see text. The mean activity level is indicated as infrared beam crosses per minute (the interval between the ticks on the y-axis is 0.2). Note that the scale is different for the ALA strain (extends until 3.0 at 14:10, 16:8, and 18:6) because these flies were more active under long photoperiods (see Fig. 4).
Under rectangular 12:12 LD cycles, M and E activity peaks were close to lights-on and lights-off, respectively, in the 3 strains, whereby the Northern HU strain showed a significant earlier M peak than the other 2 strains (Fig. 3A). Under the longer photoperiods, the M peaks of HU and CantonS occurred consistently earlier than those of ALA, but the M peaks followed lights-on quite well in all 3 strains. Just the amplitudes of the M peaks decreased with increasing photoperiod (Fig. 2), and in approximately 10% of the flies, the M peak was not visible at longer photoperiods (χ2 analysis revealed no significant differences in the frequency of M peak presence between all 3 strains: χ2 = 0.442; p = 0.802). For the E peak, the situation was different: It remained present in each single fly, but it did not track lights-off at longer photoperiods (Figs. 2 and 3). With increasing day length, the E peak occurred clearly before lights-off, and this tendency was the same in all strains. Nevertheless, differences were evident in the ability to delay the E peak towards lights-off in the 3 strains. The laboratory strain CantonS delayed its E peak most, followed by the Northern HU strain and the Southern ALA strain (Fig. 3A). We calculated a maximal YM,E of 15.6 ± 0.2 h for the laboratory strain CantonS, 14.9 ± 0.2 h for the Northern strain HU, and 14.4 ± 0.1 h for the Southern strain ALA (Fig. 3C). A 2-way ANOVA revealed that YM,E was significantly dependent on day length and the strain (day length: F4,423 = 431.194; p < 0.0001; strain: F2,423 = 24.823; p < 0.0001). Furthermore, day length and strain showed a significant interaction, meaning that the photoperiod influenced YM,E in the 3 strains differently (CantonS flies increased YM,E more than the other strains; F8,423 = 5.627; p < 0.0001).
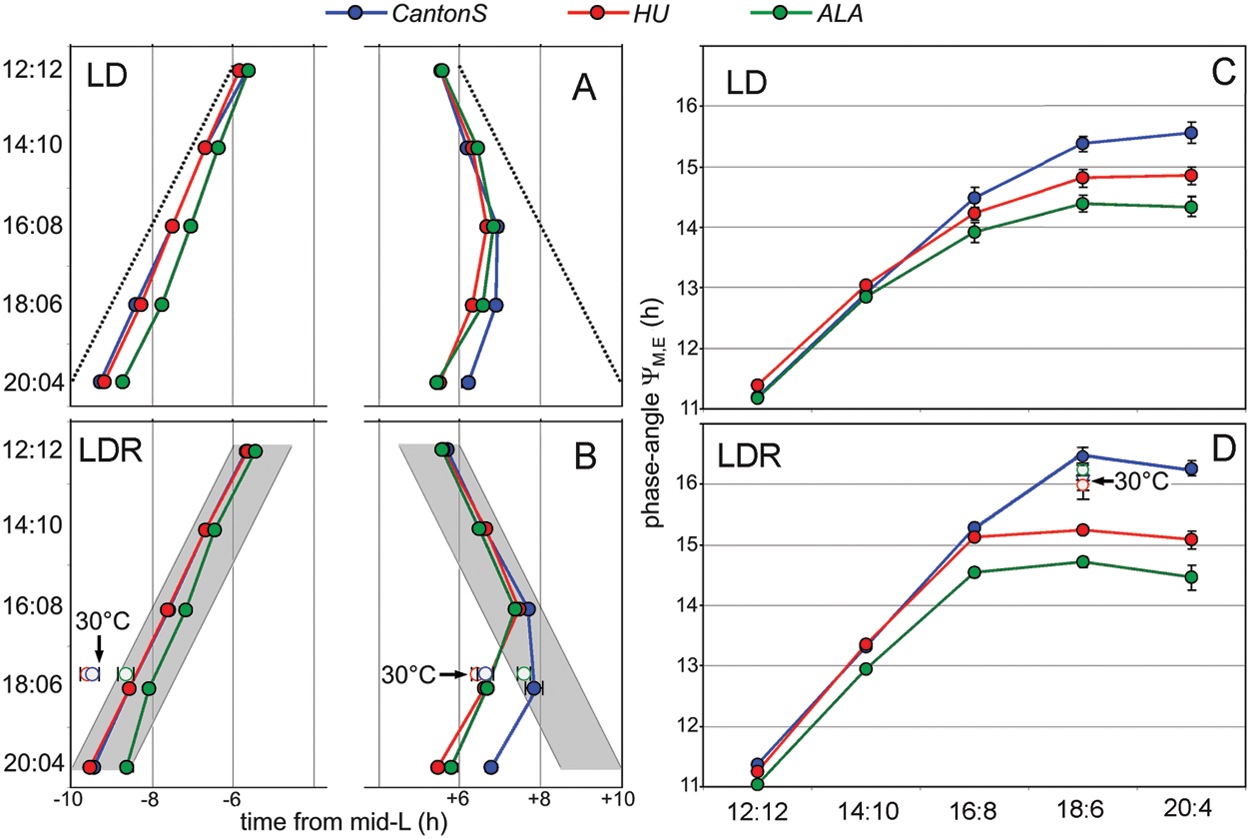
Phases of M and E peaks in relation to lights-on/lights-off (A) or to dawn and dusk (B) as well as the phase angle YM,E between M and E peaks (C, D) under the different photoperiods. Lights-on and lights-off are indicated as a broken line (A), and dawn and dusk are indicated as gray shadow (B). Faint vertical lines (A, B) have a distance of 2 hours. Simulated twilight led to a larger YM,E in each strain and at each photoperiod (CantonS: F1,157 = 97.918; p < 0.0001; HU: F1,266 = 50.264; p < 0.0001; ALA: F1,223 = 9.533; p = 0.0023). Open symbols at 18:6 LDR cycles indicate the phases of M and E peaks (B) and YM,E (D) at a constant temperature of 30 °C. All other values derive from experiments performed at 20 °C. For details, see text.
Under LDR cycles with simulated twilight, all 3 strains did extend YM,E to larger values (Figs. 2 and 3B and 3C). If visible, the M peaks perfectly tracked dawn under all photoperiods in the laboratory strain CantonS and the Northern strain HU and almost perfectly, although with a later phase, in the Southern strain ALA (as found under LD, ~10% of the flies lost the M peak under long photoperiods) (Fig. 3B). The E peak tracked dusk until a day length of 16 hours in all 3 strains (Fig. 3B). At longer day lengths, the E peaks of the Northern and Southern wild strains could not further track dusk but occurred clearly before dusk. Nevertheless, a second activity peak occurred at late dusk (Fig. 2), which we interpreted as a direct response to the rapid decrease in light intensity (see Discussion and Kempinger et al., 2009). At LDR 18:6, the dusk peak had almost the same amplitude as the E peak before dusk in ALA but was considerably smaller in HU. At LDR 20:4, the dusk peak was almost invisible in ALA but still evident in HU.
In the laboratory strain CantonS, the E peaks remained on average close to dusk until a day length of 18 hours (Fig. 3B). At the level of individual flies, we found that the E peak of 68% of the CantonS flies still tracked dusk at LDR 18:6. The remaining 32% of the flies showed an E peak before dusk that is visible as a shoulder in the average activity profile calculated from all flies (Fig. 2). In addition to this earlier E peak, these flies showed a second increase in activity at late dusk, as did the Northern and Southern wild strains. At 20:4, none of the CantonS flies was able to track dusk, but the dusk peak remained clearly visible in the average day (Fig. 2).
YM,E was always calculated between the M peak and the first E peak (if the flies showed 2 peaks). As already revealed for rectangular LD cycles, we found that the laboratory strain CantonS showed the most flexible YM,E under simulated twilight (Fig. 3C). The maximal possible YM,E for CantonS was 16.5 ± 0.2 h, followed by a maximal YM,E of 15.3 ± 0.1 h for the Northern HU strain and 14.7 ± 0.1 h for the Southern strain ALA. A 2-way ANOVA revealed that YM,E was significantly dependent on day length and the strain (day length: F4,404 = 2248.242; p < 0.0001; strain: F2,404 = 215.081; p < 0.0001) and that CantonS flies increased YM,E more than the other strains (interaction between strain and photoperiod; F8,404 = 32.903; p < 0.0001).
To test the effects of simulated twilight on YM,E, we performed a 2-way ANOVA for each strain, comparing the values under LD and LDR at long days (14 hours to 20 hours). We found that simulated twilight led to a larger YM,E in each strain and at each photoperiod (F values in the legend of Fig. 3).
For the peak phase of the E activity, ANOVA revealed no significant difference in the adaptation to long photoperiods between the Northern and Southern wild strains (F1,320 = 0.102; p = 0.752) (Fig. 3B). This was surprising because one would expect the Northern strain to adapt “better” to longer photoperiods and not only shift the M peaks to earlier phases but also the E peaks to later phases. To shed light on this apparent discrepancy, we decided to consider the waveform of the rhythms in addition to the E peak phases. Figure 4 compiles the average activity profiles for each strain at the different photoperiods in one graph. This graph reveals clear differences between the 3 strains in adapting to long photoperiods. Whereas the E activity started later with increasing day length in CantonS and the Northern strain HU, the E activity started always at the same phase in the Southern ALA strain and became just broader with increasing day length, meaning that the flies did not increase their midday trough. As a consequence, the overall daily activity of ALA was higher under long photoperiods than under 12:12 LDR cycles (Fig. 4). In CantonS and HU flies, the mean activity level remained similar at all photoperiods. This finding suggests that the Southern strain may not adapt as well to long photoperiods as the other 2 strains do.
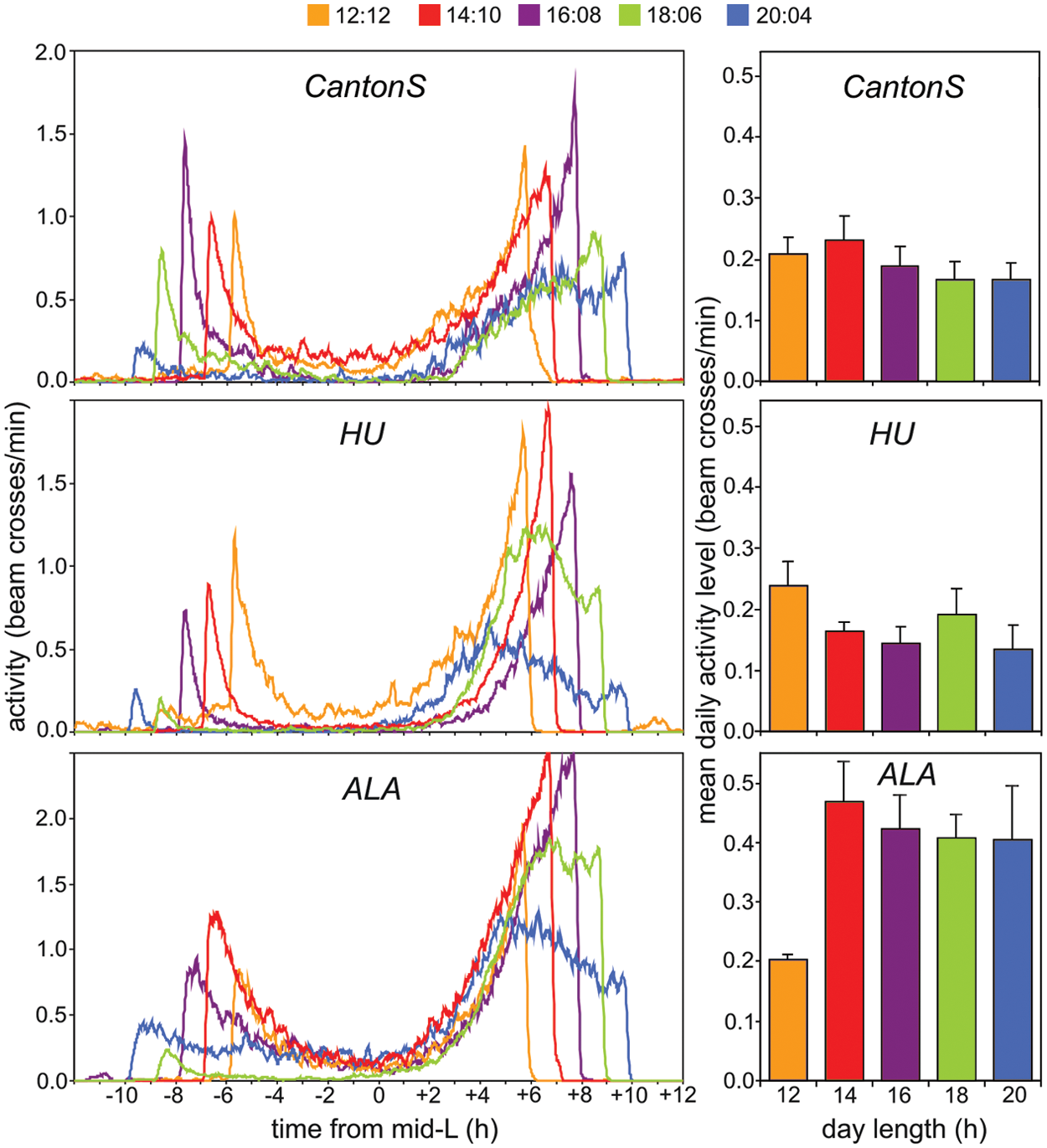
Overlay of mean activity profiles under LDR at all tested photoperiods for CantonS, HU, and ALA flies, respectively. Whereas the M peaks tracked dawn in all 3 strains, the E peaks did only partially. In CantonS and HU flies, the activity increase in the evening started later with increasing photoperiod (until 18:6 in CantonS and 16:8 in HU), but in ALA flies, the E activity increase occurred at the same time under all photoperiods. As a consequence, the E activity was longer under long photoperiods in ALA, resulting in a significantly higher mean activity level per 24 hours (right diagrams) (F1,114 = 8.839; p = 0.004; because the activity values at the different long photoperiods were not significantly different from each other, we pooled them and tested all values against the ones obtained for 12:12). In CantonS and HU flies, the mean daily activity level did not change with increasing day length (CantonS: F1,101 = 0.185; p = 0.668; HU: F1,132 = 2.227; p = 0.135; long photoperiods pooled).
The Influence of Temperature
High ambient temperatures are characteristic for summers in the South. To investigate whether high temperatures provoke a longer midday trough and a delayed E peak in ALA, we recorded the activity of the strains under 18:6 LDR cycles at 30 °C. Indeed, ALA flies exhibited now a longer siesta (not shown) and in addition delayed the E peak (and advanced the M peak) significantly as compared to 20 °C (p < 0.001) (Fig. 3B). This finding suggests that temperature is the predominant factor that determines the phase of the M and especially E peaks in the Southern strain.
The high temperature did also influence the phases of M and E peaks in the other 2 strains, but clearly in a different way. At 30 °C, the M peaks were strongly advanced in HU and CantonS flies, but the E peaks were not delayed. In CantonS flies, the E peaks were even significantly advanced at 30 °C as compared to 20 °C (p < 0.0001) (Fig. 3B). In spite of the different effects of temperature on M and E peaks in ALA and the 2 other strains, all 3 strains had a similar large YM,E at 30 °C that was not significantly different from YM,E of CantonS flies at 20 °C (F3,114 = 0.625; p = 0.600) (Fig. 3D). For ALA and HU flies, YM,E at 30 °C was significantly larger than YM,E at 20 °C (p < 0.0001).
Free-Running Period
To clarify whether the observed differences in adapting to long days are caused by a different speed of the endogenous clock, we determined the free-running periods of the 3 wild-type strains at 20 °C. We found no significant differences in period: CantonS flies showed a mean period of 24.2 ± 0.07 (n = 32), HU flies showed a mean period of 24.3 ± 0.05 (n = 56), and ALA flies showed a mean period of 24.3 ± 0.10 (n = 39).
Light sensitivity of the clock
Under simulated twilight at 20 °C, the laboratory strain CantonS and the Northern strain HU showed their M peaks (and to some extent also their E peaks) at lower irradiances than the Southern strain ALA. This suggests that the circadian clocks of CantonS and HU are more sensitive to light than the clock of ALA. To test this hypothesis, we determined the re-entrainability of the flies to very dim light, starting at an irradiance of 6.0 × 10−3 µW/cm2. At this irradiance, all flies were entrained and exhibited their typical bimodal activity pattern (not shown). When light intensity was gradually reduced and the LD cycle simultaneously phase advanced by 6 hours, obvious differences in the ability to re-entrain became evident. Whereas almost all flies of the strains CantonS and HU could re-entrain at a light intensity of 3.6 × 10−4 µW/cm2, only 70% of the ALA flies were able to re-entrain under the same conditions (Fig. 5A). The remaining ALA flies showed free run. The lower light sensitivity of ALA flies became also evident in the number of days the flies that could re-entrain needed to do so. ALA flies needed significantly longer to re-entrain than CantonS and HU flies (Fig. 5B). No significant difference was found between CantonS and HU flies, neither in the percentage of re-entrained flies, nor in the number of days the flies needed to re-entrain (Fig. 5A and 5B).
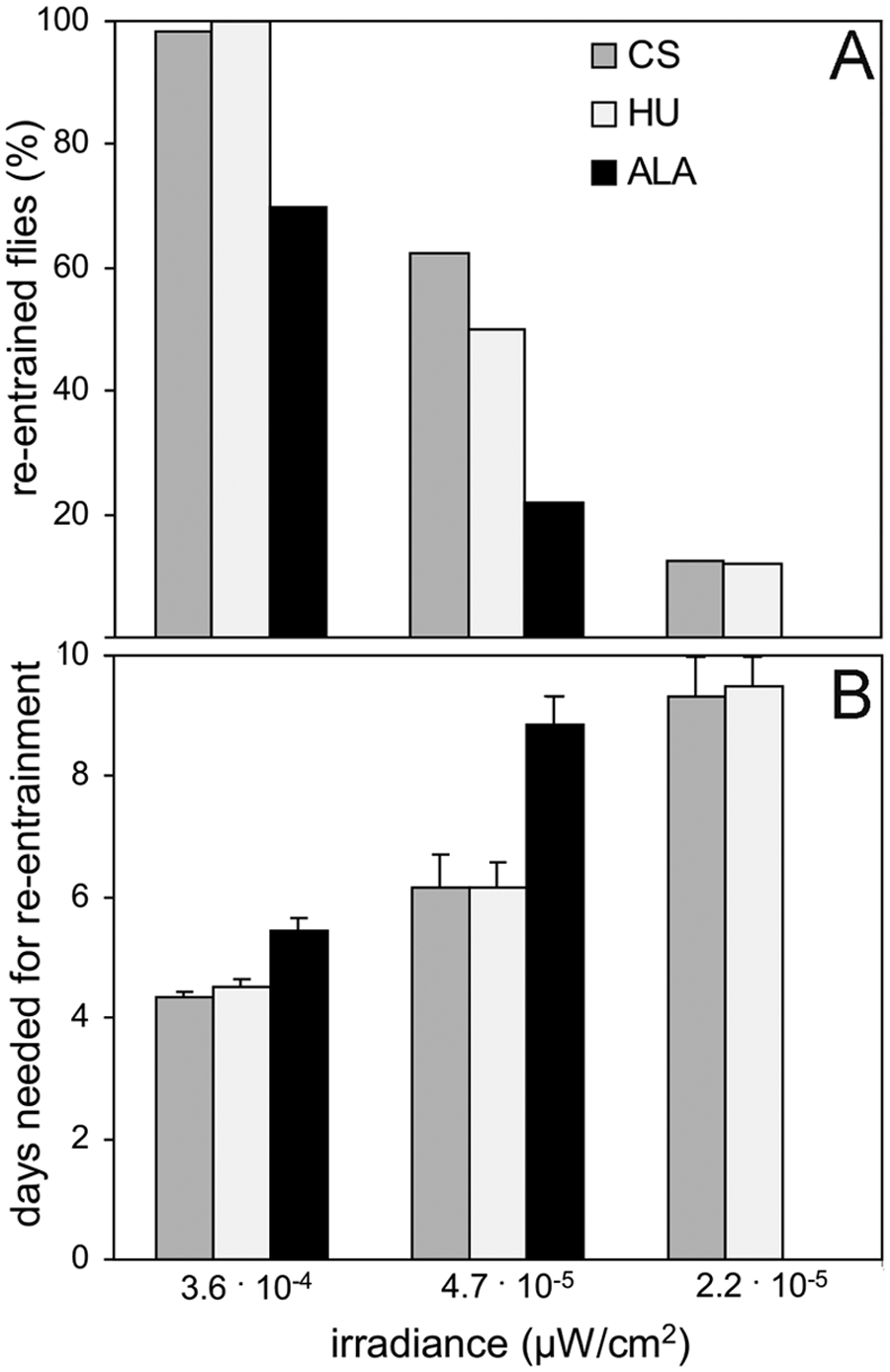
Capability of the 3 wild-type strains to re-entrain to a 6-hour phase advance of 12:12 dim-light LD cycles. (A) The percentage of flies that were able to re-entrain, and (B) the number of days (±SEM) the re-entrained flies needed to do so. As compared to CantonS and HU flies, a lower percentage of ALA flies re-entrained (χ2 = 14.87; p = 0.0006), and these needed significantly longer to do so (ANOVA and post hoc test; p < 0.0001).
tim polymorphism
Some tim polymorphisms contribute significantly to the light sensitivity of the circadian clock, for example, s-tim being more light sensitive than ls-tim (Sandrelli et al., 2007). Furthermore, the frequency of ls-tim shows a latitudinal cline with a higher frequency in Southern Europe (Tauber et al., 2007). As a consequence, the Northern HU strain had a high probability to carry the s-tim allelic form, what would be consistent with its high light sensitivity in the present experiments. On the other hand, the Southern ALA strain that consisted of a mixture of different isofemale lines should carry both tim forms with a higher incidence for ls-tim, and again, this would fit to the lower light sensitivity of this strain found in the present experiments. The laboratory strain CantonS was genotyped as ls-tim in former studies (Peschel et al., 2006), and this classification seems not to fit to the observed high light sensitivity of CantonS flies.
We determined the tim polymorphism in 3 to 10 flies of each strain to test the putative correlation of the s-tim form with a high light sensitivity. We found that the flies of the Northern HU strain carried indeed all the s-tim allele and those of the laboratory strain CantonS carried the ls-tim allele, whereas the Southern ALA strain contained as expected a mixture of both tim forms, but predominantly ls-tim (of 10 tested flies, 6 flies were ls-tim, and 4 flies were heterozygous).
Discussion
The main aims of the present study were to find out 1) whether wild D. melanogaster strains show a similar preference for dim light as does the laboratory strain CantonS, 2) whether Northern and Southern wild strains differ in the ability to adapt their locomotor activity rhythms to long photoperiods, and 3) whether simulated twilight can improve this ability. All 3 questions can be clearly answered with yes.
We found that all 3 D. melanogaster strains had crepuscular activity patterns with activity maxima during early dawn and late dusk. Furthermore, all 3 strains reduced their activity levels at higher irradiances (compare Rieger et al., 2007). We conclude that the well-described crepuscular behavior of D. melanogaster is not a unique feature of strains that were kept in the laboratory for decades but seems to be more generally true for this particular species. Similarly, the avoidance of being active at higher light intensities seems to be well fixed in the genes and not to change after many generations in the laboratory. These observations fit well with a recent study showing that D. melanogaster flies kept in complete darkness for 1340 generations (since 1954) still maintained their locomotor activity rhythms (Imafuku and Haramura, 2011).
In spite of the obvious stability of certain life history traits, we could reveal differences in the light sensitivity of the 3 strains. The clocks of the laboratory strain CantonS and the Northern strain HU were significantly more light sensitive than the clock of the Southern strain ALA, and as a consequence, the former 2 strains showed their activity peaks during simulated twilight at slightly lower irradiances. The high light sensitivity of the clock is at least partly caused by the short TIM isoform encoded by the s-timeless allele that still prevails in Northern Europe and was present in the HU line used here (the ls-tim allele originated in the South and is just about to expand to the North) (Sandrelli et al., 2007; Tauber et al., 2007). Nevertheless, these studies also showed that additional genetic factors determine the light sensitivity of Drosophila’s clock. When different natural fly strains were compared, HU flies carrying the ls-tim alleles turned out to be still more light sensitive than s-tim flies from a strain derived from Bitetto in Southern Italy (Sandrelli et al., 2007). Here, we found that CantonS flies are as light sensitive as HU flies, although the CantonS flies carried the ls-tim allele. Genes that affect the light sensitivity of the clock besides tim are, for example, jetlag and cryptochrome (Koh et al., 2006; Peschel et al., 2006, 2009) as well as putatively all other genes coding for proteins that are involved in the stability and degradation of the Tim protein.
In addition to the different light sensitivity, we could reveal significant strain differences in the ability of the flies to adapt to longer photoperiods. It was shown previously that the s- ls-tim polymorphism is also involved in seasonal adaptation (Tauber et al., 2007; Kyriacou et al., 2008): ls-tim flies have a higher incidence of entering diapause putatively, increasing the flies’ fitness in the North. Here, we found that CantonS flies had the most flexible YM,E, and as a consequence, M and E peaks followed dawn and dusk best at long days. This coincides well with the fact that CantonS flies carry the ls-tim allele. However, ALA flies that were also predominantly ls-tim showed the “poorest adaptation” to long days, whereas s-tim HU flies adapted clearly better, at least at 20 °C. Thus, the tim polymorphism can only partly explain the observed adaptation to long days.
Besides light, temperature is a major factor that influences the activity pattern of fruit flies (Tomioka et al., 1998; Majercak et al., 1999). The present study suggests that light and temperature have a different influence on the activity pattern in Southern and Northern strains. M and E activities of the Southern ALA flies had clear difficulties to follow dawn and dusk under long days at 20 °C but not at 30 °C: Whereas ALA flies started activity always about 3 to 4 hours after noon at 20 °C, they clearly delayed their E peak and consequently avoided activity after noon under long days (LDR 18:6) at 30 °C. Thus, ALA flies can perfectly adapt to the environmental constraints. The delay in E activity under long summer days is just not triggered by the photoperiod but by temperature. This makes perfect sense, given the fact that day length does not vary to such an extent throughout spring and summer in the South, but temperatures can be rather high in the summer.
This behavior was clearly different in the Northern HU strain (and the laboratory strain CantonS) that started E activity later under long photoperiods at 20 °C but surprisingly not at 30 °C. Instead, both strains considerably advanced their M peaks into the night at 30 °C, a behavior that seems to make no sense in nature. This suggests that the clocks of HU and CantonS flies have difficulties in dealing with constant high temperatures. We may argue that both strains are not used to temperatures of 30 °C, HU not accustomed because of its Northern origin and CantonS not accustomed because we kept this strain more than 30 years constantly at 20 °C. Perhaps temperature is a more stringent environmental factor that changes life history traits faster than light (compare above). Our results with the natural strains suggest that photoperiod may be the most important cue for timing activity in the temperate North, whereas temperature may be the dominant factor in the South. Nevertheless, our data are just case studies. Many more natural strains need to be tested to infer real adaptations to local environments.
Furthermore, we have to admit that several of our conditions are not natural: Our twilight simulations did lack the systematic spectral changes of light. In addition, it was completely dark during the night, and we kept irradiance during the day constantly at 100 lux and temperature constantly at 20 °C (or 30 °C), although in nature, irradiance may rise to much higher levels (~100,000 lux at maximum) and temperature oscillates with average amplitudes of approximately 10 °C, depending on latitude, altitude, and the actual weather conditions (Bhutani, 2009). We limited irradiance to 100 lux because a previous study had shown that flies avoided exposing themselves to high irradiances (Rieger et al., 2007). Thus, it seems more natural to not expose them to strong light without offering a possibility to hide. Nevertheless, our conditions with completely dark nights, no change in irradiance during the day, and no additional temperature cycle may create another artifact: the flies’ activity gets stimulated by the rather rapid increase/decrease of irradiance during twilight (Kempinger et al., 2009). Therefore, the endogenous M and E peaks may partly be masked. In nature, nights are not completely dark, and temperature is at its minimum during dawn and then continuously increases until it reaches its maximum approximately 2 hours after midday. Similarly to light-dark cycles, temperature cycles are potent zeitgebers for CantonS flies (Tomioka et al., 1998; Glaser and Stanewsky, 2005; Yoshii et al., 2002, 2005; Busza et al., 2007). When light and temperature cycles are combined during a 12-hour:12-hour day, they contribute synergistically to entrainment of the activity rhythms without obvious masking effects (Yoshii et al., 2009, 2010). Future experiments will reveal whether the natural-like combination of light and temperature cycles in simulated long days will influence behavior differently in the different wild-type strains and whether this will lead to less advanced M peaks and more delayed E peaks on hot days in CantonS and HU flies.
In spite of the discussed limitations, our study underlines the idea that differences in clock properties have evolved in adaptation to the local environmental fluctuations (Kyriacou et al., 2008). It remains exciting to uncover the underlying genes and the neuronal basis of these differences.
Footnotes
Acknowledgements
The authors thank R. Costa and C.P.K. Kyriacou for the wild strains “Houten” and “Alto Adige.” They are also most grateful to R. Costa, J. Hirsh, P. Menegazzi, and T. Yoshii for fruitful discussions of the present study and J. Hirsh for language editing. This study was supported by the European Community (6th Framework Project EUCLOCK, no. 018741).
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.