
Abstract
Pigment-dispersing factor–immunoreactive neurons anterior to the accessory medulla (aPDFMes) in the optic lobes of insects are circadian pacemaker neurons in cockroaches and fruit flies. The authors examined whether any of the aPDFMes of the cockroach Leucophaea maderae are sensitive to changes in period and photoperiod of light/dark (LD) cycles as a prerequisite to adapt to changes in external rhythms. Cockroaches were raised in LD cycles of 11:11, 13:13, 12:12, 6:18, or 18:6 h, and the brains of the adults were examined with immunocytochemistry employing antisera against PDF and orcokinin. Indeed, in 11:11 LD cycles, only the number of medium-sized aPDFMes specifically decreased, while it increased in 13:13. In addition, 18:6 LD cycles increased the number of large- and medium-sized aPDFMes, as well as the posterior pPDFMes, while 6:18 LD cycles only decreased the number of medium-sized aPDFMes. Furthermore, PDF-immunoreactive fibers in the anterior optic commissure and orcokinin-immunoreactive fibers in both the anterior and posterior optic commissures were affected by different lengths of light cycles. Thus, apparently different groups of the PDFMes, most of all the medium-sized aPDFMes, which colocalize orcokinin, respond to changes in period and photoperiod and could possibly allow for the adjustment to different photoperiods.
For equator-distant organisms, the length of the light/dark (LD) cycles varies regularly during the course of the year because of the inclination of the Earth’s axis. In addition, because of the gravitational pull of the moon on the Earth’s oceans, the rotation of the Earth around its axis is gradually slowed, causing the length of a day to change since the first organisms inhabited the Earth. Thus, a main task of endogenous clocks such as circadian clocks and photoperiodic clocks is to allow for entrainment to changing external geophysical rhythms. Accordingly, photoperiodic clocks not only entrain to but also appear to measure the duration of the photophase (“photoperiodic timer”) and appear to count the number of consecutive long or short days (“photoperiodic counter”) before they initiate photoperiodic adaptations such as pupal diapause (Shiga et al., 2003; Emerson et al., 2009). It is under debate whether photoperiodic clocks and circadian clocks are constituted by the same cellular circuits. In mammals and insects, a dual circadian oscillator model was proposed previously to explain adjustment of locomotor rhythms to different photoperiods (Pittendrigh and Daan, 1976). Recently, in the fruit fly Drosophila melanogaster, it was suggested that a zeitgeber-dependently environmentally modifiable network of circadian clock-gene-expressing neuronal groups in the brain is responsible for either controlling the morning (M-cells) or the evening (E-cells) peak of locomotor activity (Helfrich-Förster, 2009). While in the fruit fly, the pigment-dispersing factor–immunoreactive (PDF-ir) small ventral lateral neurons (sLNv) are assumed to be M-cells, which are phase advanced by light at the late night, in the cockroach Leucophaea maderae, PDF-ir neurons are also known while the nature of M- and E-cells remains to be discerned.
In the cockroach L. maderae (= Rhyparobia maderae), lesion and transplantation experiments located the circadian pacemaker, which controls locomotor activity rhythms to the optic lobes’ accessory medulla (aMe) with associated PDF-ir neurons (Stengl and Homberg, 1994; Reischig and Stengl, 2003a). The about 12 PDF-ir neurons anterior to the aMe (aPDFMes) can be subgrouped according to soma size into 4 small, 4 medium-sized, and 4 large aPDFMes. Posterior to the aMe are the less well-described 2 small and 2 large posterior PDF-ir neurons (pPDFMes). The respective functions of the different PDF-ir neurons in different neuronal circuits are not well understood. However, backfill studies combined with immunocytochemistry suggested that the 4 medium-sized aPDFMes colocalize orcokinin and FMRFamide-immunoreactivity and project via the anterior optic commissure (AOC) to ipsi- and contralateral midbrain targets. Three of them reach as far as into the contralateral optic lobe (Reischig and Stengl, 2004; Soehler et al., 2008). The 4 large aPDFMes do not colocalize orcokinin or FMRFamide. They project via the lobula valley tract to the AOC and posterior optic commissure (POC) and branch in different ipsilateral and/or contralateral midbrain targets, and only one of them (the conspicuously largest) reaches into the contralateral optic lobe (Soehler et al., 2008). The 4 pPDFMes are assumed to restrict their arborizations to optic lobe neuropils while not projecting to the midbrain. Their respective branching pattern within the aMe neuropil is not known. The small PDFMes appear to be local neurons of the aMe and are not well characterized due to their low level of PDF-immunoreactivity (Reischig and Stengl, 2003b). Therefore, they are not taken into account in this study.
In the Madeira cockroach, PDF is an input and output signal of the circadian clock (Petri and Stengl, 1997), while in the German cockroach, PDF is necessary for circadian rhythmicity (Lee et al., 2009), and in the cricket Gryllus bimaculatus, PDF plays a role for photic entrainment next to additional functions in the clock (Abdelsalam et al., 2008; Hassaneen et al., 2011). In the Madeira cockroach, orcokinin and not PDF is assumed to play a role in photic entrainment because orcokinin injections produce a light-like phase response curve with delays at the early night and advances at the late night (Hofer and Homberg, 2006a). Whether light or darkness is an input signal to PDF-dependent neuronal circuits in the brain of the Madeira cockroach is not known. Furthermore, while PDF-ir circadian pacemaker neurons are assumed to play a role in photoperiod-controlled behaviors in flies (Hamanaka et al., 2007; Shiga and Numata, 2009), this is not known for the Madeira cockroach.
Here we examined in L. maderae whether PDF-ir neurons are sensitive to changes in photoperiod (the duration of the photophase) and period (the number of hours per day) of LD cycles. On the basis of previous work (Barrett and Page, 1989; Page and Barrett, 1989), we tested whether the PDF-ir branching pattern or the number of their respective PDF-ir somata was changed via different LD regimes during development: 11:11 (T22), 12:12 (T24), 13:13 (T26), 18:6 (long days), and 6:18 (short days). Indeed, changes in LD regimes during development affected primarily the number of PDF-ir medium-sized aPDFMes, which colocalize orcokinin and project via the AOC to the contralateral aMe. Thus, light regimes either affect the survival of PDFMes or the expression of PDF in PDFMes during development. This indicates that even the circadian system of a tropical species without immediate need to adjust to changes in photoperiod has the plasticity to allow for radiation into different climatic zones.
Materials and Methods
Animals
Adult male cockroaches (L. maderae) were reared at constant conditions: temperature 26 °C and humidity 60%, with 5 different light/dark cycles—that is, 11 h light–11 h dark (LD 11:11), 12 h light–12 h dark (LD 12:12), 13 h light–13 h dark (LD 13:13), 6 h light–18 h dark (LD 6:18), and 18 h light–6 h dark (LD 18:6). All brains were dissected at about the same zeitgeber time (ZT) 1:00 to avoid ZT-dependent effects.
Primary Antisera
The anti-β-PDH antiserum (#3B3), an affinity-purified rabbit antiserum against synthetic Uca pugilator β-PDH, was generated by Dr. H. Dircksen (University of Stockholm, Sweden) (Dircksen et al., 1987). Its specificity was shown by immunodot-blotting assays of high-pressure liquid chromatography (HPLC)–separated crude sinus gland extracts (Dircksen et al., 1987; Mangerich et al., 1987). The specificity of the antiserum for cockroach PDF has been demonstrated through preadsorption controls (Stengl and Homberg, 1994; Petri et al., 1995; Reischig and Stengl, 1996). The monoclonal anti-Drosophila-PDF (#C7) antibody raised in mouse was provided by the Developmental Studies Hybridoma Bank. To examine its specificity, the PDF-ir network in the cockroach wholemount brain was labeled by using anti-β-PDH antiserum diluted at 1:20,000 and anti-Drosophila-PDF diluted at 1:5 simultaneously. Both antisera labeled the same neuronal structures (data not shown). The anti-Asn13-orcokinin antiserum (provided by Dr. H. Dircksen) was raised in rabbits. Its specificity on cockroach brain sections was determined using liquid-phase preadsorption by Hofer and Homberg (2006b).
Immunocytochemistry on Paraffin Sections
The brains of cockroaches raised in LD 11:11 (T22; n = 29) and LD 13:13 (T26; n = 34) hours were processed for immunocytochemistry on paraffin sections. The immunocytochemical staining protocol was performed as described before (Reischig and Stengl, 2003b). The brains were dissected from the head capsule and fixed overnight in 4% paraformaldehyde (PFA) at 4 °C. Subsequently, they were washed (3 times for 10 min) with 0.1 M phosphate-buffered saline (0.1 M PBS, pH 7.4) and dehydrated in an ethanol series and embedded in paraffin (Paraplast plus; Sigma-Aldrich, Deisenhofen, Germany). Serial frontal 10-µm sections were cut with a rotary microtome and mounted on microscope slides. Afterwards, the samples were deparaffinized with xylene and rehydrated using graded ethanol concentrations. The brain sections were stained with anti-β-PDH for 18 to 24 h diluted at 1:3000 in 0.1 M PBS containing 0.5% Triton X-100 (0.1 M PBT) and 1% normal goat serum (NGS; Jackson ImmunoResearch, West Grove, PA). Immunoreactive cells were detected by using a 3-step peroxidase technique (Sternberger, 1979; Reischig and Stengl, 1996). After a thorough rinsing with PBT (3 times for 10 min), the sections were incubated for 1 h in goat-anti-rabbit IgG diluted at 1:100 and for 1 h in rabbit PAP (DAKO, Hamburg, Germany) at a dilution of 1:300. After washing again, the brains were stained with 0.03% 3,3′-diaminobenzidine tetrahydrochloride (DAB; Sigma-Aldrich) and 0.015% H2O2 in 0.1 M PBS for 10 min. To visualize the neuropils of the brain, the sections were counterstained with 1% methylene blue.
Double-Staining on Sections
Double-staining experiments with the anti-Asn13-orcokinin and the monoclonal anti-Drosophila-PDF were performed with cockroaches, reared in short day lengths (LD 6:18; n = 20), normal day lengths (LD 12:12; n = 20), and long day lengths (LD 18:6; n = 20). The brains were dissected and fixed as described above. After fixation, the brains were embedded in a gelatine/albumin mixture (4.8% gelatine and 12% ovalbumin in demineralized water), postfixed in 8% formalin, and sectioned using a vibratome (Leica, Nussloch, Germany) in a frontal plane of 200 µm thickness. The free-floating sections were washed for 10 min in 0.1 M PBT for 6 times and preincubated for 3 h with 5% NGS and 0.02% sodium azide in 0.1 M PBT at room temperature. The primary antisera anti-Asn13-orcokinin (from rabbit) and anti-Drosophila-PDF (from mouse) were diluted at 1:4000 and 1:5 in 0.1 M PBT containing 1% NGS and 0.02% sodium azide and were incubated on sections for 3 days. After 6 consecutive washes in 0.1 M PBT for 10 min each, the sections were stained with a Cy2-conjugated goat anti-mouse secondary antibody (Cy2-GAM; Jackson ImmunoResearch) and a Cy3-conjugated goat anti-rabbit secondary antibody (Cy3-GAR; Jackson ImmunoResearch) (diluted in 1:300) in 0.1 M PBT containing 1% NGS and 0.02% sodium azide for 2 days. The sections were washed before dehydration in an increasing ethanol series (0%, 50%, 70%, 90%, 95%, 100%, 5 min each). They were precleared in a mixture of 50% methyl salicylate (MS; Merck, Darmstadt, Germany) and 50% ethanol for 5 min and cleared with MS for 15 min. Finally, to prevent compressions, the sections were mounted in Permount (Fisher Scientific, Pittsburgh, PA) in the center of 3 spacers between 2 coverslips (Zweckform, Oberlaindern, Germany).
Immunocytochemistry for Wholemount Brains
To determine the developmental plasticity of the PDF-ir network regarding the different day and night lengths, the wholemount brains of the cockroaches reared in LD 6:18 (n = 16), LD 12:12 (n = 18), and LD 18:6 (n = 18) were stained with anti-β-PDH antiserum. The procedure was described previously in detail (Wei et al., 2010). Briefly, dissected brains were fixed as described above. The brains were washed in 0.1 M PBS 5 times for 15 min each before treating with 1 mg/mL collagenase for 1 h. After washing 6 times with 0.1 M PBT for 20 min, the brains were preincubated overnight at 4 °C with 5% NGS and 0.02% sodium azide in 0.1 M PBT. The brains were incubated in anti-β-PDH antiserum diluted at 1:20,000 in 0.1 M PBT containing 1% NGS and 0.02% sodium azide for 5 days at 4 °C. After another washing step with 0.1 M PBT over 2 h, the primary antibody was detected by Cy3-GAR diluted at 1:300 in 0.1 M PBT containing 1% NGS and 0.02% sodium azide for 4 days at 4 °C. After washing again, the brains were dehydrated in an increasing series of ethanol, precleared in a mixture of 50% methyl salicylate and 50% ethanol, and cleared with methyl salicylate for 40 min. Finally, the wholemounts were mounted in Permount.
Visualization and Evaluation
The PDF-ir structures in paraffin sections were observed using a Zeiss microscope (Zeiss Axio Scope A1). A confocal laser-scanning microscope (Leica TCS SP5) was employed to image the 200 µm sections and wholemounts. The fluorescence signals of Cy3 were excited with a helium/neon laser at 543 nm and detected between 560 nm and 580 nm. Cy2 fluorescence was excited with a 488 nm argon laser and detected between 500 nm and 520 nm. The orcokinin-ir and PDF-ir structures were consecutively photographed using a 20x objective at a resolution of 1024 × 1024 pixels in the x-y plane and an interslice interval of 0.5 µm (axial distance). Due to the thickness and width of the wholemounts, they were scanned in 8 image stacks that were combined using Amira 3.1 and 4.1 (Visage Imaging, Fürth, Germany). For better structural analysis, the orcokinin-ir fibers in one optic lobe were three-dimensionally reconstructed with the “skeleton reconstruction” tool in Amira 3.1 (Wei et al., 2010).
To determine whether different LD cycles cause environmentally induced plasticity of the PDF-ir network, the somata of large- and medium-sized anterior PDF-ir somata next to the medulla (aPDFMes) and of the posterior PDF-ir neurons next to the medulla (pPDFMes) were counted in all experimental groups. In addition, the number of PDF-ir fibers in the AOC and POC was counted (Fig. 1). Furthermore, the length of the AOC and POC and the distances between all PDF-ir arborization sites in animals with LD 6:18, 12:12, and 18:6 were measured (n = 10). Finally, the number of orcokinin-ir fibers in the orcokinin-ir AOC and POC was counted in all 3 experimental groups. The data were compared and analyzed using SPSS 13.0 and Origin 6.0.
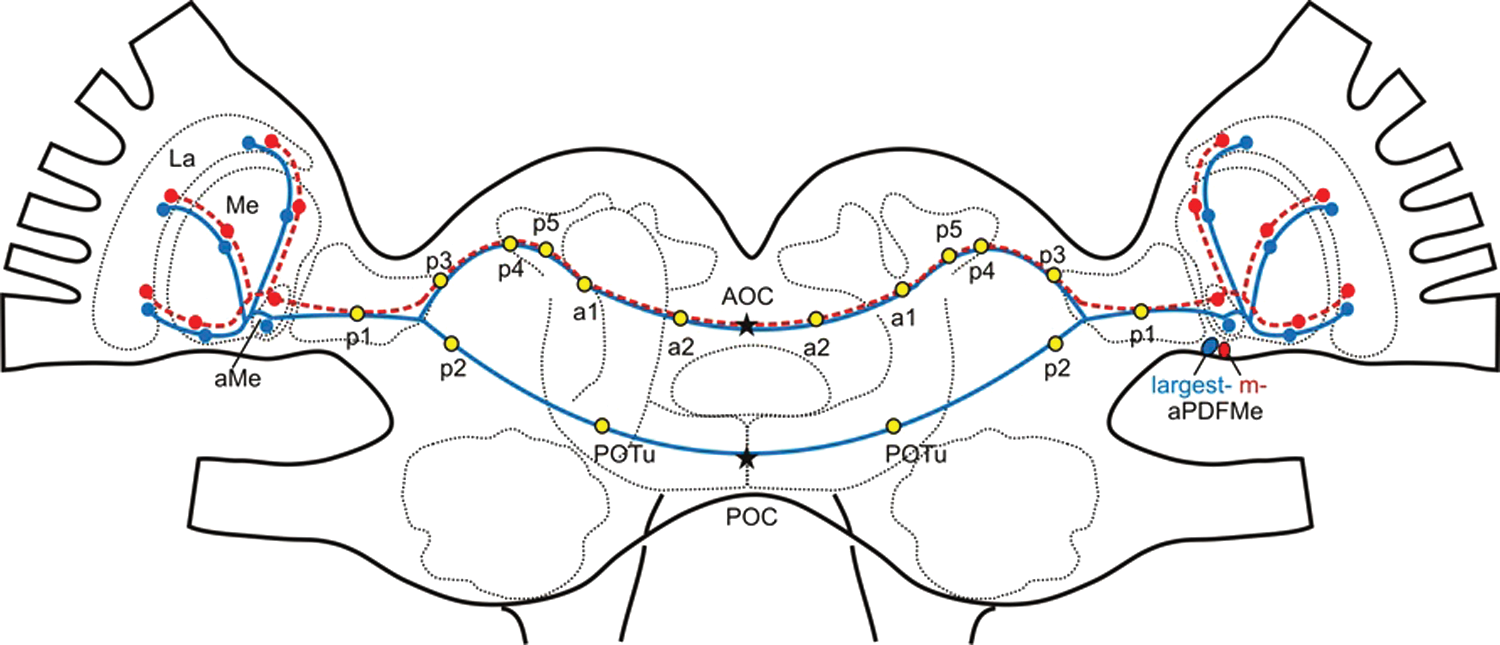
Arborization pattern of the pigment-dispersing factor–immunoreactive (PDF-ir) neurons connecting the ipsi- and contralateral optic lobe is shown in a brain model of the cockroach L. maderae. One particularly large PDF-ir neuron (blue) anterior to the medulla (aPDFMe) connected both optic lobes via the anterior and posterior optic commissures (AOC, POC). Three medium-sized PDF-ir neurons (red) cross the brain midline via the AOC. Along their way, they gave rise to several branching sites: 5 plexi (p1-5), 2 areas (a1-2), and the posterior optic tubercle (POTu). Stars mark the sites of fiber counts taken. La = lamina; aMe = accessory medulla; Me = medulla.
Results
To determine whether different photoperiods and different T cycles affect PDFMes, we examined whether rearing in different light regimes changes PDF-ir circadian pacemaker neurons associated with the accessory medulla (aMe) in L. maderae. We hypothesized that circadian pacemaker neurons with long endogenous periods are more activated via long-period T cycles compared to short-period T cycles and that light-sensitive cells are more activated via long photoperiods compared to short photoperiods. Since activation of neurons during development affects survival, we expected to see changes in the number of respectively affected cells. Furthermore, we wanted to distinguish pacemaker neurons that are affected by light and darkness equally or not equally. Therefore, cockroaches were raised in non-24-h LD (T) cycles 11:11 (T22; n = 29) and 13:13 (T26; n = 34) as well as in different photoperiods with long days (18:6; n = 18) or short days (6:18; n = 16). Then, the brains of the adult cockroaches were processed for PDF-immuno-cytochemistry. Double-labeling with anti-orcokinin together with anti-PDF antibodies was only performed in animals raised under different photoperiods (n = 20). The number of different groups of PDF-ir neuronal cells anterior or posterior to the aMe (aPDFMes, pPDFMes), as well as the number and length of PDF- or orcokinin-ir processes in optic lobe commissures, was evaluated. Then, the different experimental groups were compared to cockroaches raised in 12:12 (T24 = control; n = 18) LD cycles.
Different T Cycles Only Changed Medium-Sized aPDFMes
The number of immunoreactive somata of large aPDFMes and pPDFMes in cockroaches reared under the 3 different T cycles showed no significant difference (analysis of variance [ANOVA] least significant difference [LSD] p > 0.05). However, the number of somata of medium-sized aPDFMes in T22 animals (n = 29) was significantly smaller, and in T26 animals (n = 34), it was significantly higher as compared to T24 animals (n = 18) (ANOVA LSD p < 0.05) (Fig. 2A). The number of the PDF-ir fibers in the AOC in T22 and T24 animals showed no significant difference (ANOVA LSD p > 0.05). In contrast, the number of the PDF-ir fibers in T26 animals was higher in the AOC than in T22 or T24 animals (ANOVA LSD p < 0.05). The number of the PDF-ir fibers in the POC was not affected by changes in zeitgeber period (ANOVA LSD p > 0.05) (Fig. 2B).
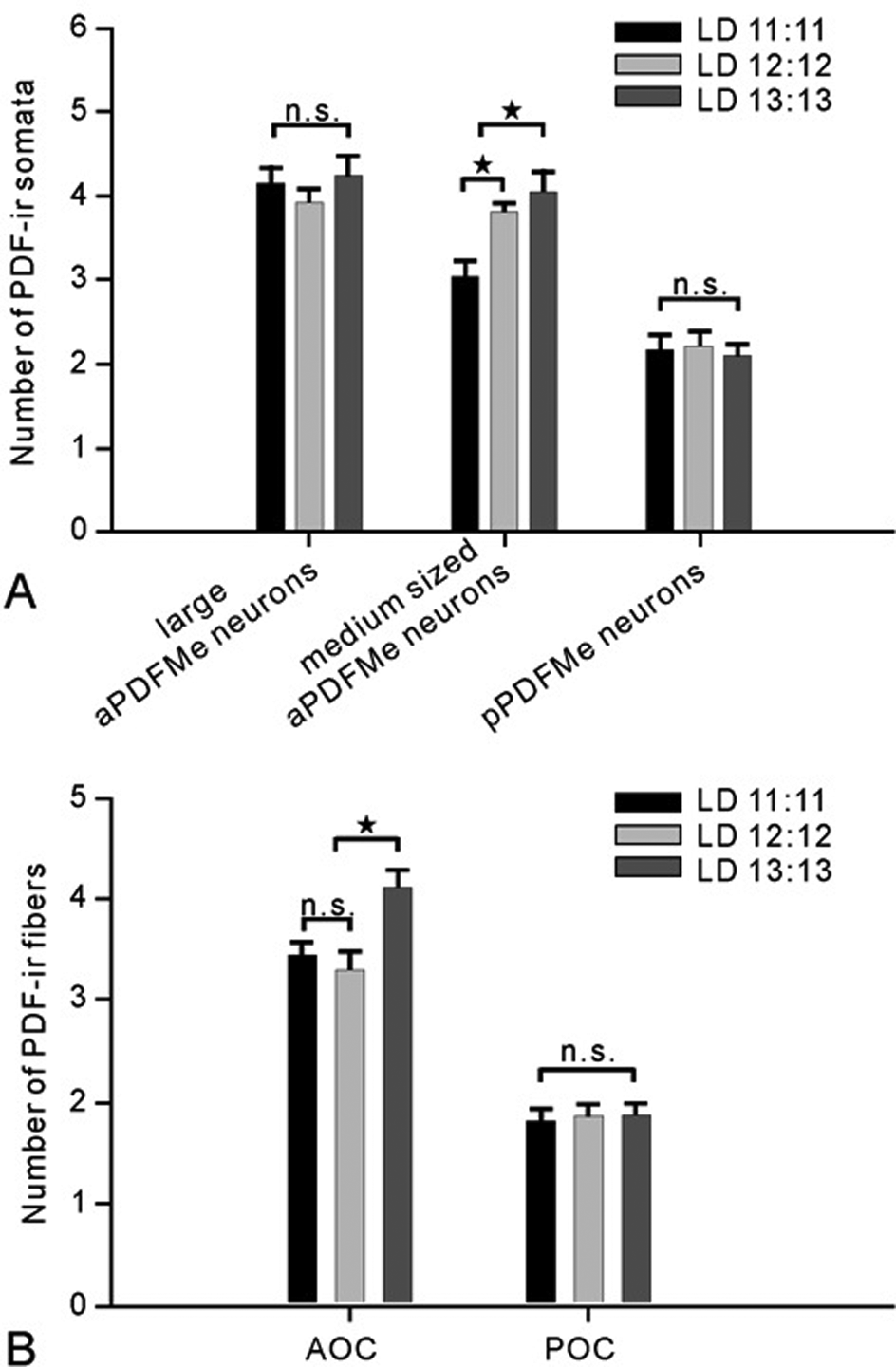
(A, B) Cockroaches raised in different light/dark (LD) time (T) cycles: 11:11 (T22; n = 29), 12:12 (T24; n = 18), and 13:13 (T26; n = 34) showed differences in the number of pigment-dispersing factor–immunoreactive (PDF-ir) somata and commissural processes (bars, standard error). The different light and dark regimes did not affect the soma numbers of large anterior PDF-ir neurons (aPDFMes) or of posterior PDF-ir neurons (pPDFMes), nor did they affect the number of PDF-ir fibers in the posterior optic commissures (POC) (ANOVA LSD p > 0.05; n.s. = not significant). However, the number of medium-sized aPDFMes in T24 animals was significantly higher than in T22 and lower than in cockroaches raised in T26 (ANOVA LSD p < 0.05, star). T26 animals displayed a significantly higher number of PDF-ir fibers in the anterior optic commissures (AOC) than T22 and T24 animals (ANOVA LSD p < 0.05, star).
Different Photoperiods Changed All Groups of PDFMes Differentially
The number of immunoreactive somata of the large aPDFMes and of the pPDFMes did not differ significantly in cockroaches raised in LD 6:18 (n = 16) and 12:12 (n = 18) (ANOVA LSD p > 0.05), while fewer medium-sized aPDFMes were counted in short day-rearing conditions (ANOVA LSD p < 0.05) (Fig. 3A). The cockroaches raised in LD 18:6 (n = 18) maintained significantly more somata of large- and medium-sized aPDFMes and of pPDFMes than those raised under LD 6:18 or 12:12 (ANOVA LSD p < 0.05) (Fig. 3A). The number of PDF-ir fibers in the AOC and POC showed no significant difference among the 3 experimental groups (ANOVA LSD p > 0.05) (Fig. 3B). The length of the PDF-ir fibers in the AOC and POC did not show a significant difference between the cockroaches raised in LD 6:18 and 12:12 (ANOVA LSD p > 0.05; n = 10). However, the cockroaches raised in LD 18:6 exhibited significantly longer fibers in the AOC and POC compared to those raised in LD 6:18 or 12:12 (ANOVA LSD p < 0.05; n = 10) (Fig. 3C). Linear regression analysis (n = 10) showed that the relative distances of the different PDF-ir arborization sites in the aMe, the 5 plexi (p1-p5), the posterior optic tubercle (POTu), and the 2 arborization areas (a1, a2) were regulated by different photoperiods (Fig. 4) (Wei et al., 2010).
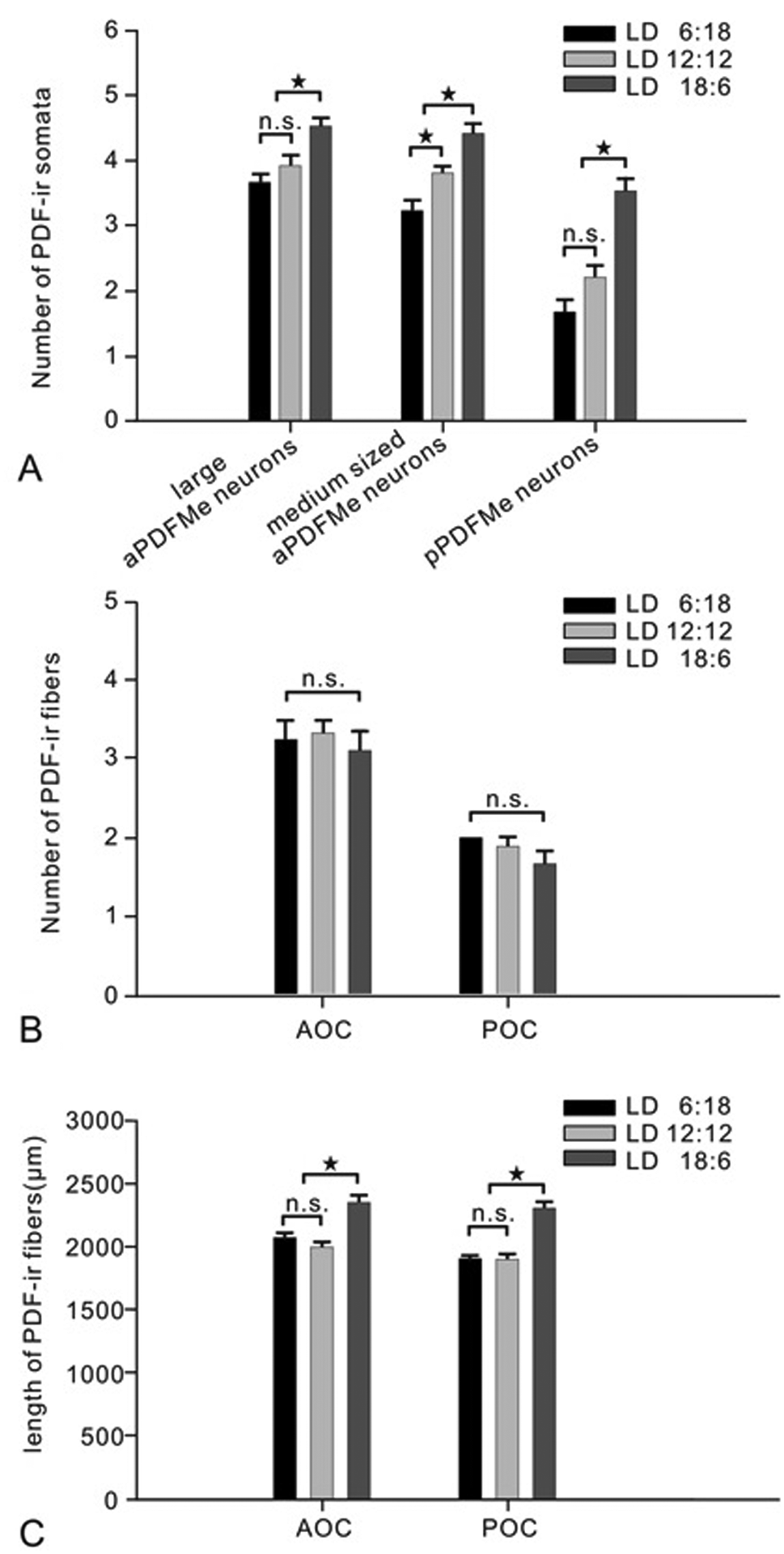
(A-C) Cockroaches reared in light/dark (LD) cycles 6:18 (n = 16), 12:12 (n = 18), and 18:6 (n = 18) showed significant differences in the number of the pigment-dispersing factor–immunoreactive (PDF-ir) somata as well as in the length of commissural processes (bars, standard error). More large- and medium-sized anterior PDF-ir neurons (aPDFMes) as well as more posterior PDF-ir neurons (pPDFMes) occurred in cockroaches with LD 18:6 compared to animals raised in LD 6:18 and 12:12. In the 6:18 light regime, only medium-sized aPDFMes decreased in number as compared to the other experimental groups (ANOVA LSD p < 0.05, A star). The number of PDF-ir fibers in the anterior and posterior commissures (AOC, POC) showed no significant difference among the 3 animal groups (ANOVA LSD p > 0.05 B, n.s. = not significant). The longer PDF-ir fibers in the AOC and POC occurred in animals under LD 18:6 and were absent in those animals under LD 6:18 and 12:12 (ANOVA LSD p < 0.05, C star, n = 10).
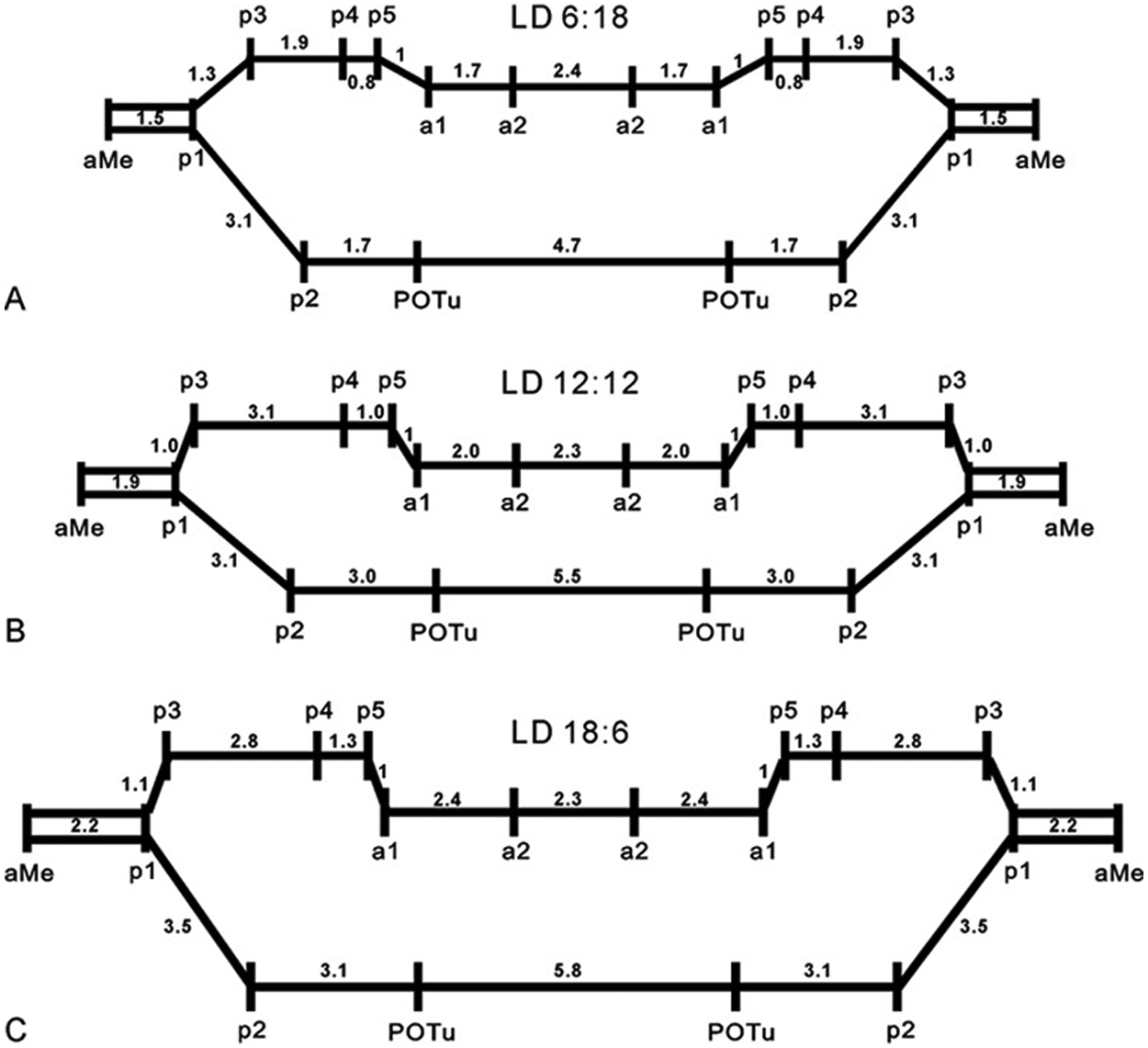
(A-C) The light/dark regimes (LD 6:18 in A, 12:12 in B, and 18:6 in C) changed the distances between pigment-dispersing factor–immunoreactive (PDF-ir) arborization sites (n = 10): 5 plexi (p1-5), 2 areas (a1-2), and posterior optic tubercle (POTu). The distance between p5 and area a1 was defined as “1.” Linear regression analysis calculated the respective multiples of this distance for the other connections. These were changed under different LD conditions.
Different Photoperiods Changed the Orcokinin-ir Network
Next to the network of PDFMes, orcokinin-ir neurons were further examined (n = 20) because all medium-sized aPDFMes colocalize orcokinin and PDF immunoreactivity (Soehler et al., 2011). In addition, orcokinin appears to act in the light-entrainment pathway of the circadian clock (Hofer and Homberg, 2006b; Soehler et al., 2011). Our results confirmed previous findings of orcokinin-ir branching patterns in the cockroach brain (n = 20). However, at least 3 new observations were reported in this study, based on 20 preparations examined.
First, at least 3 of the 4 medium-sized aPDFMes differentially colocalized PDF with orcokinin immunoreactivity (n = 20). Always, 1 soma contained relatively stronger PDF immunoreactivity as compared to weaker orcokinin immunoreactivity. In contrast, always the other 2 somata expressed strong orcokinin and weaker PDF immunostaining (Fig. 5A).
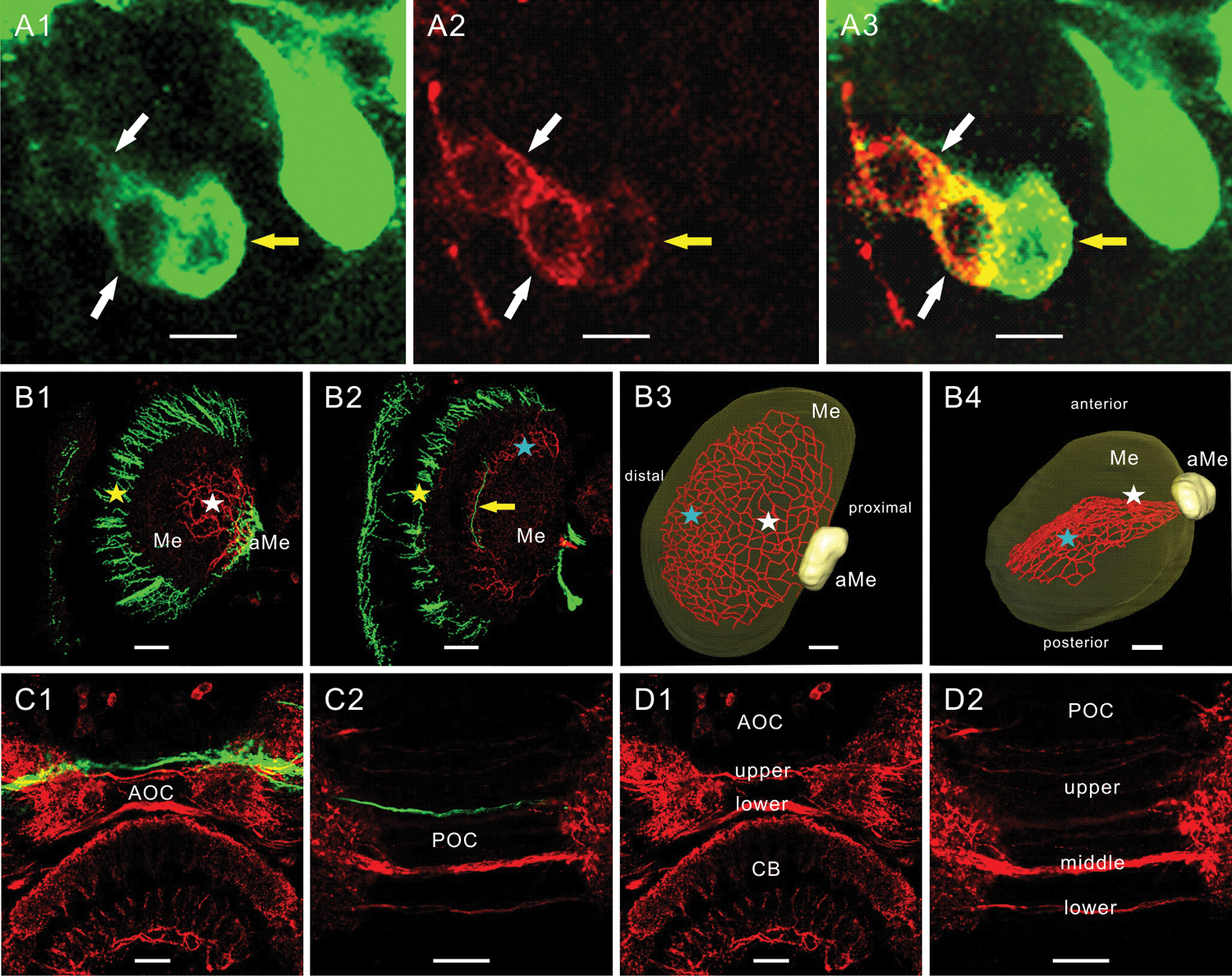
(A-D) Orcokinin-immunoreactive (orcokinin-ir) and pigment-dispersing factor–immunoreactive (PDF-ir) fibers in the optic lobes (A, B) and at the level of the anterior and posterior optic commissures (AOC, POC) (C, D). PDF immunoreactivity is shown in green and orcokinin immunoreactivity is shown in red. (A1-A3) Three medium-sized anterior PDF-ir neurons (aPDFMes) expressed colocalization and always differential intensity of staining with both peptide antisera (n = 20). Always 1 soma showed stronger PDF and weaker orcokinin immunoreactivity (yellow arrow), while the other 2 somata expressed more orcokinin and less PDF immunoreactivity (white arrows). The overlay image in A3 proved the colocalization of both peptides. (B1-B4) Orcokinin- and PDF-ir fibers in the optic lobes do not express colocalization of both peptides. The PDF-ir neurons connected the accessory medulla (aMe), the medulla (Me), and the lamina via the anterior fiber fan (B1, B2, yellow star) and via the median layer fiber system (B2, yellow arrow). The orcokinin-ir fibers ran from the posterior aMe, fasciculated in a proximal-median layer of the medulla (B1, B3, B4, white stars), and projected distal-posteriorly in a layer of the medulla toward the first optic chiasma (B2, B3, B4, blue stars). They formed a fiber network covering at least one complete layer in the medulla. The median layer fiber system of the PDF-ir neurons (B2, yellow arrow) and the fiber meshwork of the orcokinin-ir cells (B2, blue star) arborized in the same layer of the medulla. (C1-D2) No colocalization occurred in the upper and lower fiber bundles in the AOC or in the upper, middle, and lower fiber bundles in the POC. CB = central body. Scale bar = 10 m in A, 50 m in B-D
Second, the PDF-ir fibers from the anterior aMe projected toward the medulla and lamina via 2 distinct pathways: the anterior fiber fan covering the anterior and distal surface of the medulla (Fig. 5B1-B2, yellow star) and the median layer fiber system in the medulla (Fig. 5B2, yellow arrow). In contrast, the orcokinin-ir fibers from the posterior aMe extended into the medulla and ramified extensively in a proximal-median layer of the medulla (Fig. 5B1, B3-B4, white star). Dense orcokinin-ir fibers ran in a distal-posterior direction and gave rise to a fiber meshwork parallel to the distal surface of the medulla (Fig. 5B2, B3-B4, blue star). Despite the colocalization of both peptides in apparently all medium-sized aPDFMe somata of the ventral neurons next to the aMe, the median layer fiber system of the PDF-ir neurons and the fiber meshwork of the orcokinin-ir cells arborized in the same layer of the medulla without expressing any colocalization (Fig. 5B2). Clearly, colocalization of PDF and orcokinin immunoreactivity could not be observed in any of the described PDF-ir branching sites (n = 20) (Wei et al., 2010; Soehler et al., 2011).
Third, two distinguishable orcokinin-ir fiber fascicles crossed the brain midline via the AOC (Fig. 5C1, D1). Animals raised in LD 6:18 or LD 18:6 exhibited either fewer or more fibers in both bundles as compared to cockroaches raised under LD 12:12 (ANOVA LSD p < 0.05) (Fig. 6). The PDF-ir fibers in the AOC ran near the upper orcokinin-ir fiber bundle (Fig. 5C1), and 2 fiber bundles (middle and lower POC in Fig. 5D2) with dense orcokinin immunoreactivity connected both aMae via the POC. Several faint orcokinin-ir fibers were found superior to these 2 fiber bundles in the POC (upper POC in Fig. 5D2). The number of fibers in the middle bundle showed significant changes in response to LD environmental changes. Lengthening the light phase and shortening the dark phase (LD 18:6) induced a significant increase of the orcokinin-ir fibers in the middle bundle, while shortening the light phase and increasing the dark phase (LD 6:18) decreased the number of orcokinin-ir fibers in the middle bundle (ANOVA LSD p < 0.05) (Fig. 6). The number of fibers in the upper bundle and lower bundle showed no significant difference among the 3 experimental groups. Generally, only 2 orcokinin-ir fibers projected via the lower bundle. The orcokinin-ir fibers in the POC did not show colocalization with PDF immunoreactivity (Fig. 5C2).
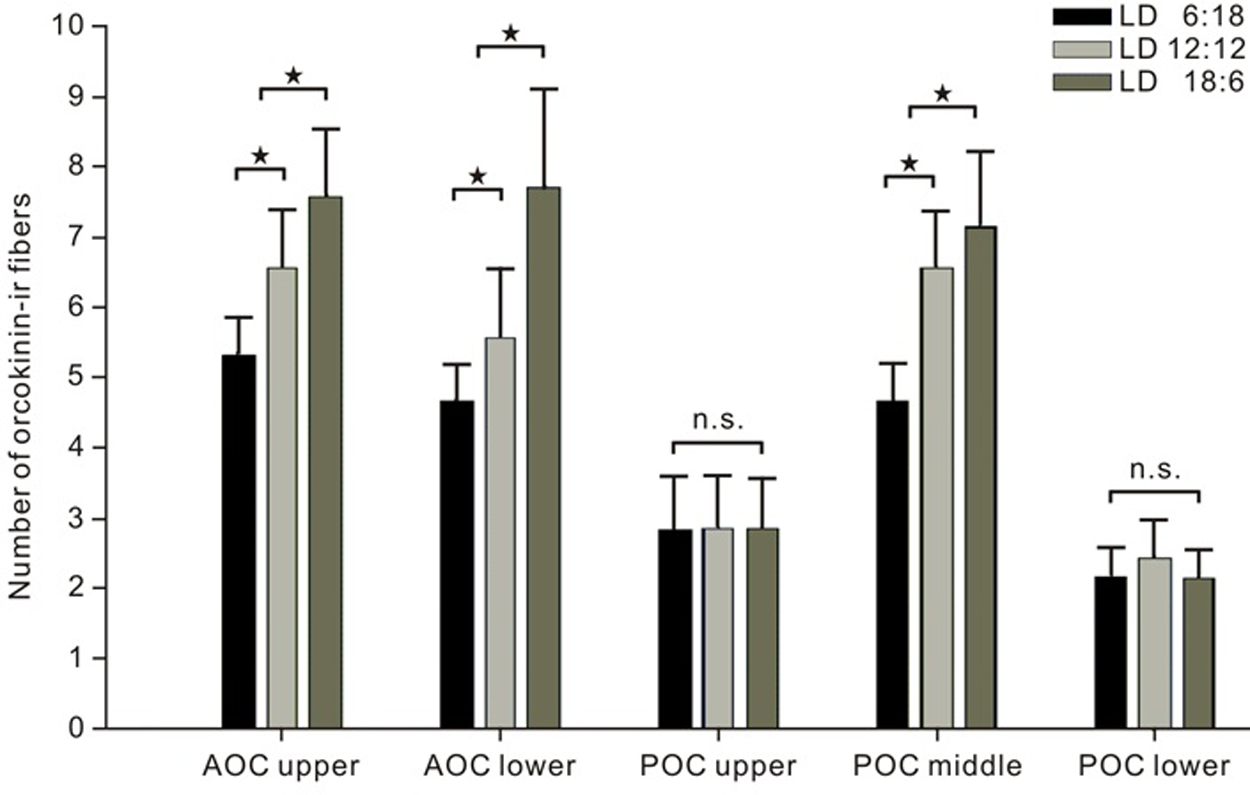
Cockroaches raised in light/dark (LD) 6:18, 12:12, and 18:6 showed differences in the amount of orcokinin-immunoreactive (orcokinin-ir) commissural fibers (n = 20; bars, standard error). The number of the orcokinin-ir fibers in the upper and lower fiber bundles in the anterior optic commissure (AOC) as well as in the middle fiber bundle in the posterior optic commissure (POC) increased significantly with lengthening of the light phase while shortening the dark phase (ANOVA LSD p < 0.05, star, n = 20). The number of fibers in the upper and lower bundles in the POC showed no significant correlation with the different LD conditions (ANOVA LSD p > 0.05, n.s. = not significant, n = 20).
Discussion
To determine whether PDF-ir circadian pacemaker circuits are sensitive to different photoperiods and T cycles, rearing in different light cycles was combined with immunocytochemistry employing antisera against PDF and orcokinin. After raising cockroaches L. maderae in different light regimes, T22, T24, T26, 6:18, and 18:6 LD, it was found that non-24-h periods as well as different length of the light phase in T24 animals changed the soma number and branching pattern of immunoreactive PDFMes specifically, with the medium-sized aPDFMes, which project via the AOC, being the most LD-regime responsive among the PDF-ir neurons. In all cases, we were not able to distinguish whether the number of immunoreactive cells changed in the different light regimes due to regulated survival of neurons or due to regulated PDF or orcokinin expression in the respective cells. This, however, would not affect the main conclusions concerning light responsiveness of the circadian pacemaker neurons.
Plasticity of the Optic Lobes
In accordance with our findings, it was known from previous studies in vertebrates as well as in invertebrates that light-dependent neuronal activity shapes neuronal circuits in the visual system (reviews: Hirsch et al., 1995; Heisenberg et al., 1995; Aamodt and Constantine-Paton, 1999). In D. melanogaster, changes in light levels changed the volume of light-dependent neuropils in the brain (Barth and Heisenberg, 1997). The lamina and medulla were largest in flies raised in constant light (LL) and smallest in flies reared in constant darkness (DD), mainly due to changes in the volume of photoreceptor cell terminals (Barth et al., 1997). The activity-dependent morphological changes in the optic lobes also were accompanied by changes in the physiology, such as altered contrast sensitivity (Deimel and Kral, 1992; Kral and Meinertzhagen, 1989). The experience-dependent plasticity in the optic lobes was interpreted as a form of long-term memory that allows adaptation to varying environmental conditions. Thus, it is very likely that in the cockroach, the light-dependent change in the length of the PDF-ir processes reflects respective light-dependent changes in the volume of PDF-ir neuropils. It is not completely resolved how light affects the PDF-ir l-LNvs to control PDF release at dawn in the fruit fly to elicit arousal (Parisky et al., 2008; Park and Griffith, 2006). Even less is known about the pre- and postsynaptic cells of the PDF-ir neurons in the cockroach.
Light Affects PDF-ir Neurons Differentially in the Cockroach L. maderae
Our studies show that the medium-sized aPDFMes, which project via the AOC, are affected via different T cycles and different length of the light phase during different photoperiods. One possible explanation would be that the medium-sized aPDFMes have longer endogenous periods and couple better to long T cycles as compared to short T cycles. Alternatively or additionally, the about 4 medium-sized aPDFMes are a diverse group of cells. Most of them are activated more by light than inhibited by darkness, and at least one is inhibited more by darkness than activated by light. Alternatively, only light or darkness of a minimum duration activates or inhibits the medium-sized aPDFMes ZT-independently. As the most likely hypothesis, we suggest that the medium-sized aPDFMes are only activated by light and inhibited by darkness during ZT 11-13 but are not affected via photic stimuli at other zeitgeber times. In contrast, the large aPDFMes and the pPDFMes appear to receive balanced activity by light and inhibition by darkness since the different T cycles did not affect them. However, apparently a longer light duration compared to the dark phase appears to activate these cells. Alternatively, we find it most likely that these cells are only light-responsive during a time window between ZT 13 and 18. Because the number of PDF-ir fibers in the AOC increased in T26 but did not increase in 18:6 LD cycles, possibly some of the contralaterally projecting medium-sized aPDFMes are activated by light while others are inhibited by decreases in the duration of darkness.
How light reaches the PDF-ir circuits in the cockroach is not completely clear. Lesion studies revealed that photoreceptors in or near the compound eyes provide light entrainment of locomotor activity rhythms (Nishiitsutsuji-Uwo and Pittendrigh, 1968; Roberts, 1965, 1974). Anti-histamine immunocyto-chemistry revealed that there is no direct photoreceptor input to the aMe, and the photoreceptors terminate in the lamina and a distal layer of the medulla (Loesel and Homberg, 2001). Light input to the circadian clock appears to be relayed via the GABAergic distal tract, which projects over the distal face of the medulla to the noduli of the aMe (Petri et al., 2002). In addition, intracellular recordings identified a light-excited GABAergic neuron next to the aMe with tangential projections to a median layer of the medulla, arborizations in the noduli of the aMe, and projections to accessory neuropils of the lamina via the fiber fan over the anterior face of the medulla (Loesel and Homberg, 2001). Contralateral light input appears to reach the noduli of the aMe via 3 orcokinin-immunoreactive ventromedian neurons, which are excited by light and are polarization sensitive (Loesel and Homberg, 2001; Hofer and Homberg, 2006a, 2006b). They connect both aMae with a median layer of both medullae via the POC (Loesel and Homberg, 2001). Possibly, the processes of these light-sensitive orcokinin-ir ventromedian neurons project via the middle bundle in the POC, which changed in fiber number light-regime-dependently. Furthermore, injection studies combined with locomotor activity assays support a role of orcokinin, next to allatotropin and GABA in the processing of photic inputs via the noduli of the aMe (Petri et al., 2002; Hofer and Homberg, 2006a). Orcokinin, allatotropin, and GABA injections into the vicinity of the aMe shifted locomotor activity daytime-dependently. Because their respective phase response curves resembled light-dependent biphasic phase response curves, in contrast to PDF (Petri and Stengl, 1997), it was assumed that the respective orcokinin-, allatotropin-, and GABA-ir cells are part of the light entrainment pathway. Finally, centrifugal neurons with processes in the median protocerebrum, which are inhibited by light, send axonal arborizations to the aMe (Loesel and Homberg, 2001). Whether there is also light input via extraocular photoreceptors from the cryptochrome-positive lamina and lobula organs remains to be examined (Fleissner et al., 2001).
Previous studies examined how light affects circadian pacemaker development in L. maderae raised in non-24-h light rhythms (Barrett and Page, 1989; Page and Barrett, 1989). It was shown that raising cockroaches in 11:11 LD (T22) cycles throughout their development caused a significant reduction in the delay portion of the light-dependent phase response curve in addition to a shortening of their endogenous period (τ). In contrast, in cockroaches raised in 13:13 LD (T26), the advance portion of the light-dependent phase response curve was significantly reduced and τ lengthened. The cellular basis of the experience-dependent plasticity of the circadian system remained unknown. In addition, it was unknown whether clock genes affect circadian system plasticity as described in the fruit fly by Fernández et al. (2008). Our data suggest that the medium-sized aPDFMes might at least partly relay light-dependent phase delays and might possibly also inhibit light-dependent phase advances. Therefore, medium-sized aPDFMes might be part of the neuronal circuit, which encodes light-dependent phase delays of locomotor activity at the end of the day and the beginning of the night, possibly via long endogenous pacemaker periods. Thus, the medium-sized aPDFMes are candidates for an E oscillator circuit in the Madeira cockroach, different from the PDF-ir sLNvs in D. melanogaster. Since PDF injections cause phase delays of locomotor activity at the late day and because the number of medium-sized aPDFMes decreased in T22 cycles, this assumption is in accordance with previous evidence (Barrett and Page, 1989; Page and Barrett, 1989; Petri and Stengl, 1997; Schneider and Stengl, 2005). Whether the decrease in the light-dependent advances of locomotor activity in T26 cycles at least partly depends on the increase in the number of immunoreactive medium-sized and large posterior PDFMes remains to be examined further. Thus, our data add further evidence to the assumption that even in a tropical species, the pacemaker center in the aMe not only regulates 24-h rhythms. It also has the potential to control ultradian rhythms of action potentials in the gamma frequency range (Schneider and Stengl, 2007) and might even allow for adaptation to photoperiodic cycles during the course of the year.
Footnotes
Acknowledgements
We are grateful to Dr. H. Dircksen for the generous supply with antibodies and to Johannes Grosshans for preparation of the antiserum-labeled sections. Dr. Thomas Reischig and Rita Zintl provided some of the preparations in different day lengths. In addition, we thank our referees, which considerably improved our manuscript. Supported by DFG grants STE 531/15, STE 531/18, and STE 531/21.
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.