
Abstract
In most temperate zone vertebrates, photoperiodic change plays the major role in the timing of seasonal breeding. However, direct environmental stimuli such as temperature, rainfall, or availability of food are thought to be important for fine-tuning breeding activities. Building on evidence from wild Island canaries (Serinus canaria), the authors had shown advancing effects of green vegetation on breeding under captive, short-day conditions. So far, the precise, sensory modalities of this stimulatory cue are unknown. Here the authors present new data that confirm advanced breeding activities in the presence of green vegetation and narrow its stimulatory components. They found that direct exposure of the birds to fresh green vegetation represented the strongest stimulus and advanced breeding by up to 2 months compared to controls. In contrast, access to artificial green vegetation, extracts from green vegetation, or olfactory components alone had no such effects. This is, to the best of the authors’ knowledge, the first experiment that examines sensory components of an effective, supplementary, non-photoperiodic cue in a temperate zone species. The data suggest that in order to use non-photoperiodic information to time breeding, birds must be able to integrate and process multisensory stimuli. Single non-photoperiodic sensory cues are insufficient to affect the timing of seasonal breeding.
Animals have evolved annual rhythms in response to changing environments. Outside the tropics, photoperiod constitutes the most reliable predictor of seasonal change and, therefore, plays an important role in the timing of animals’ life cycle events. In particular, synchronizing reproduction with favorable environmental conditions is crucial for an individual’s fitness (Farner, 1985; Wingfield et al., 1992). For birds of the temperate zone, such optimal breeding conditions are tightly linked to the vernal increase in day length. Hence, photoperiod acts as initial predictive cue to regulate the onset of gonadal development, thus dictating the time window for reproduction. In order to fine-tune the exact laying date, photic information is integrated with supplementary cues such as temperature, rainfall, or food availability (Wingfield and Farner, 1993; Hahn et al., 1997; Wingfield et al., 2000). In populations of song sparrows (Melospiza melodia), for example, that live at similar latitudes but at different altitudes, the degree of gonadal recrudescence in males differs by up to 2 months, which is correlated to differences in the daily temperature range and to differences in the appearance of new green shoots and flowers on the vegetation (Perfito et al., 2004).
Animals are faced with the challenge of effectively integrating environmental information encoded by multiple sensory modalities to be able to respond appropriately. Moreover, the relative importance of such different sensory stimuli is likely to vary over time depending on the changing environment. In interindividual communication, the processing of multisensory signals clearly has advantages for the receiver, as it provides insurance against misinterpreting the message (Partan and Marler, 2005). Multisensory enhancement (i.e., the combination of sensory modalities) that increases the impact of the signal has been shown experimentally for avian courtship behavior (Uetz and Roberts, 2002; Partan et al., 2005). Female pigeons, presented with courtship displays of males, responded more often with courtship behavior when both auditory and visual information was available than when either component was presented alone (Partan et al., 2005).
While in most species of mid and high latitudes, gonadal maturation occurs in response to a stimulatory long-day photoperiod (for review, see Dawson, 2008), we obtained evidence that the wild ancestors of the domesticated canary, the Island canary (Serinus canaria) within the Madeiran archipelago (32°35′ N, 16°32′ W), can initiate reproduction under a short-day photoperiod, even before the winter solstice, which is about 6 weeks earlier than the long-term average of breeding activities in this species (Schmitz, 1899; Voigt and Leitner, 1998; Leitner et al., 2003). We found this observation to correlate with the early growth of fresh green vegetation, resulting from unusually large amounts of rainfall in the preceding months. In a subsequent experimental study using a captive group of Island canaries, we verified green vegetation but not rainfall as the stimulatory cue (Voigt et al., 2007).
Relying on other than the main predictive cue (photoperiod) increases the risk of a failed breeding attempt; therefore, such cues must convey a strong message. While supplementary cues such as temperature and rainfall give more information about immediate conditions, the vegetation integrates to some extent those cues and thus provides reliable information about environmental conditions over the previous months. Green vegetation per se constitutes a complex cue, encoding multiple sensory stimuli such as visual, olfactory, and tactile information, and the importance of each of those components for stimulating reproductive activity is likely to differ. Moreover, there is evidence that ingestion of fresh green plant material triggers reproduction in a small herbivorous mammal (Negus and Berger, 1977) and accelerates photostimulated ovarian growth in white-crowned sparrows (Zonotrichia leucophrys gambelii; Ettinger and King, 1981). Therefore, we aimed to identify the actual stimulatory component in the complex cue provided by green vegetation. We partitioned the cue into the three components—olfaction, vision, and ingestion—and presented those stimuli alone or combined and analyzed advancement of reproductive activity in comparison to birds breeding under control conditions.
Materials and Methods
Animals and Housing
Subjects were a captive population (n = 28) of Island canaries (S. canaria), also referred to as wild canaries, kept at the Max Planck Institute for Ornithology in Seewiesen, Germany (47°58′ N, 11°14′ E). The birds are descendents of individuals that were wild-caught on the main island of Madeira (32°49′ N, 17°16′ W) in 1995, and those of the same wild strain obtained from local breeders in 2004 in Germany. Birds were kept under the natural photoperiod (civil twilight) of the Madeiran archipelago (32°50′ N): maximum 15.2/8.8 L/D (June 18-25), minimum 10.9/13.1 L/D (December 19-26), and temperature ranged from 16.2 to 23.9 °C, according to the variations in their natural habitat (Fig. 1). Three months before the start of the experiment in each year, all individuals were randomly assigned to two experimental groups and one control group. Each group was kept in one indoor aviary (250 × 200 × 215 cm) throughout the study period. Mean sex ratio (M:F) was 1.5:1. Aviaries were equipped with wooden perches, nest bowls, and nesting material (coconut fibers, cotton wool). Canary seed mixture, weaning food, and water were provided ad libitum. The floors were equipped with a layer of wooden chips.
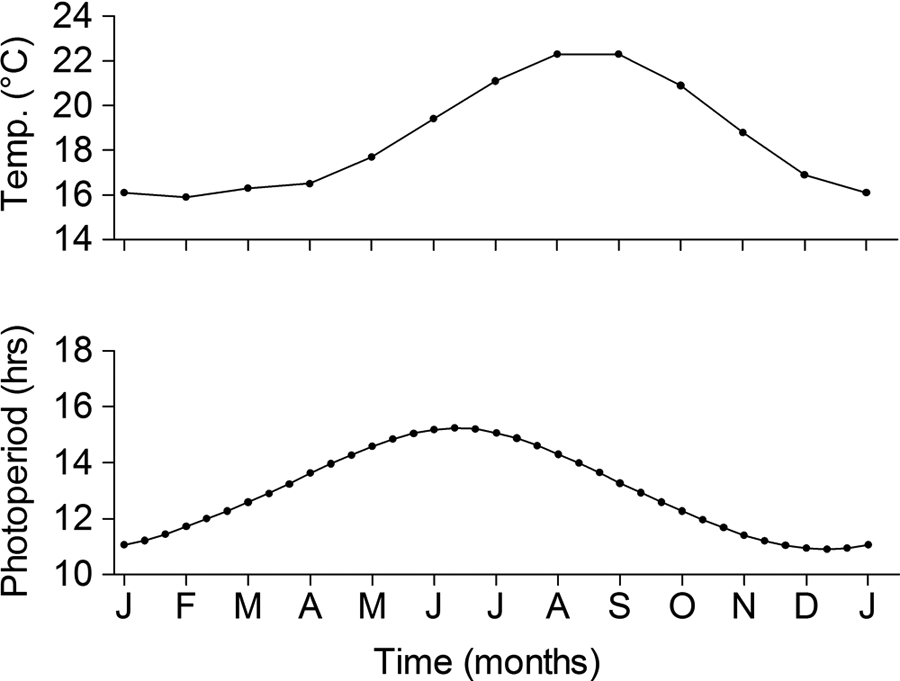
Photoperiod (civil twilight) and mean monthly temperature (average of 30 years, after Sträßer, 1998) within the Madeiran archipelago (32°50′ N).
Experimental Design
Experiments were performed during 3 consecutive years (2007 to 2009), respectively from the end of November until the end of August. The start of the experiment corresponded to “Madeiran short-day” (10.9/13.1 L/D). Treatments were assigned randomly to the 3 groups in each year and are outlined in Table 1. Birds remained in their respective aviary for the entire study period. Manipulations were performed during the last week of November and persisted the entire breeding season (except for plant extracts). Behavioral observations were carried out 3 times a week throughout the experimental period, and breeding activities were recorded. The breeding season was defined as the time period from the onset of first nest-building activities until hatching of the last clutch.
Overview of the Different Treatments throughout the Experimental Period
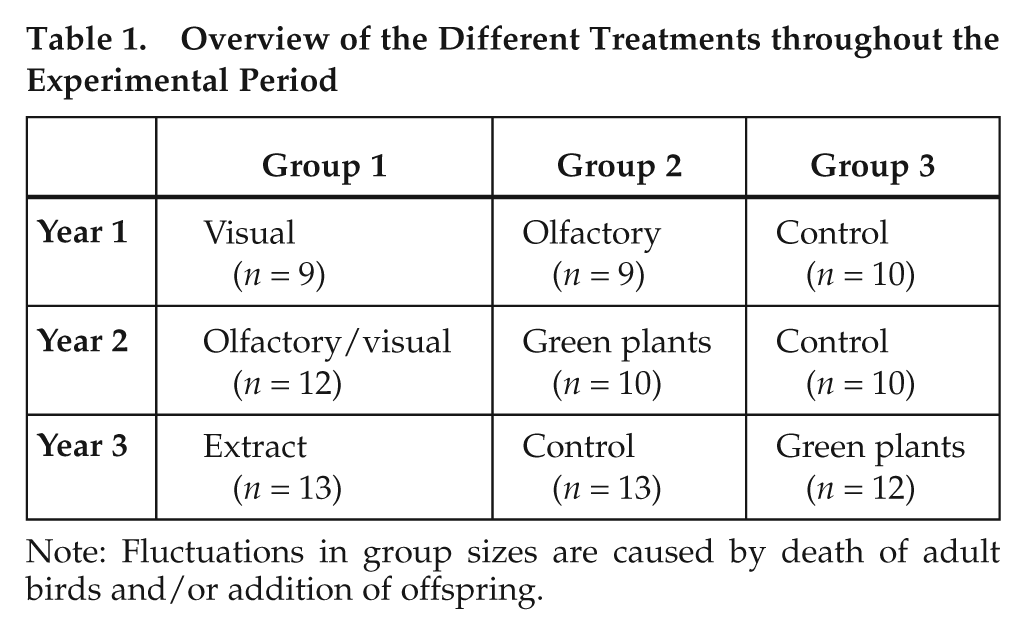
Note: Fluctuations in group sizes are caused by death of adult birds and/or addition of offspring.
Vegetation
Plant material used in the experiment consisted of a 3-m2 layer of common meadow grass (Poa pratensis); 2 bamboo (Phyllostachys bissetii), 2 m high, and 2 coniferous tree species; and Dwarf Alberta spruce (Picea glauca conica) and European Black Pine (Pinus nigra) 1.5 m high, respectively. Those plant species can be found in the natural habitat of Island canaries. Further, they proved to be successful in advancing breeding activities in a previous study (Voigt et al., 2007).
Treatment
Olfactory stimulus
Birds were exposed to fresh green vegetation placed outside the aviary behind an opaque cover. The plants’ aroma could penetrate the top of the aviary that was covered with wire mesh.
Visual stimulus
Birds were exposed within the aviary to the plant material as plastic replica, including artificial turf.
Olfactory and visual stimulus
Birds were exposed to fresh green vegetation placed behind the wire mesh inside the aviary.
Multisensory stimulus
Birds were exposed to fresh green vegetation within the aviary. This setup allowed for olfactory, visual, and physical interaction as well as ingestion.
Ingestive stimulus (plant extracts)
Birds received a daily dose of plant extract provided with weaning food (method described below).
Control
Birds were not exposed to any manipulations, and aviaries contained only the standard equipment as described above.
Plant Extraction
In total, 300 mL of fresh leaves of grass (ca. 45 g), pine (ca. 50 g), and bamboo (ca. 35 g) was extracted at room temperature by immersion for 2 h consecutively in the following solvents: 250 mL of methanol, dichloromethane, and hexane. The extracts were filtered through filter paper and concentrated to approximately 5 mL on a rotation evaporator (40°C, 150 mbar). The different solvent extracts for each plant species were combined and taken up in 25 mL methanol/dichloromethane in total. Each day, 3 mL of extract (5.5 g fresh-weight grass leaves, 6 g pine needles, 4 g bamboo leaves) was applied on 30 g of weaning food (Quiko, Bocholt, Germany). For solvents to evaporate, the food was left for 15 to 20 min before administering to the birds. The 3 ingredients were applied one type per day in a random order. The daily food consumption did not differ between these 3 components (F2, 88 = 0.05, p = 0.96). Administration of plant extract was discontinued about 1 month after the first nest had been built.
Statistical Analysis
Statistical analyses were performed with R 2.10.1 (2009; R development core team). We compared the onset of nesting and the laying date of the first egg separately using one-way analysis of variance (ANOVA) with experimental treatment as factor. Because across years, control birds did not differ significantly in the onset of breeding, year was dropped as a factor from the model (p > 0.1). Post hoc comparisons were performed with the Dunnett test by comparing all experimental groups against the control group. We used Pearson correlation to investigate the relationship between onset of breeding and reproductive success.
Results
Breeding Activity
Experimental manipulations significantly influenced the onset of nest building (F5, 23 = 5.39, p = 0.002) and the laying date of the first egg (F5, 22 = 4.82, p = 0.004). Post hoc comparisons showed that the group that received the full stimulus of green vegetation (“multisensory”) advanced the beginning of nest building by almost 8 weeks compared to the control group (mean difference ± SEM: 54.9 ± 13.4 d; Dunnett post hoc test: t = 4.09, p = 0.0011; Fig. 2A). Similarly, the laying date of the first egg in this group was significantly earlier than in the control birds (mean difference ± SEM: 46.3 ± 13.7 d; Dunnett post hoc test: t = 3.37, p = 0.0065; Fig. 2B). The period between start of nest building and start of egg laying was consistent among females (p > 0.65 for all comparisons). Those groups that received the olfactory or the visual stimulus also advanced breeding activities by several weeks compared to controls, but these differences were not significant (date of first egg, visual: mean difference ± SEM: 37.4 ± 18.8 d; Dunnett post hoc test: t = 1.99, p = 0.119; olfactory: mean difference ± SEM: 19.1 ± 18.8 d; Dunnett post hoc test: t = 1.01, p = 0.486). No effect on the onset of breeding was found in the groups that received the olfactory-visual stimulus (t = 0.15, p = 0.929) and the ingestive stimulus (t = 1.27, p = 0.998).
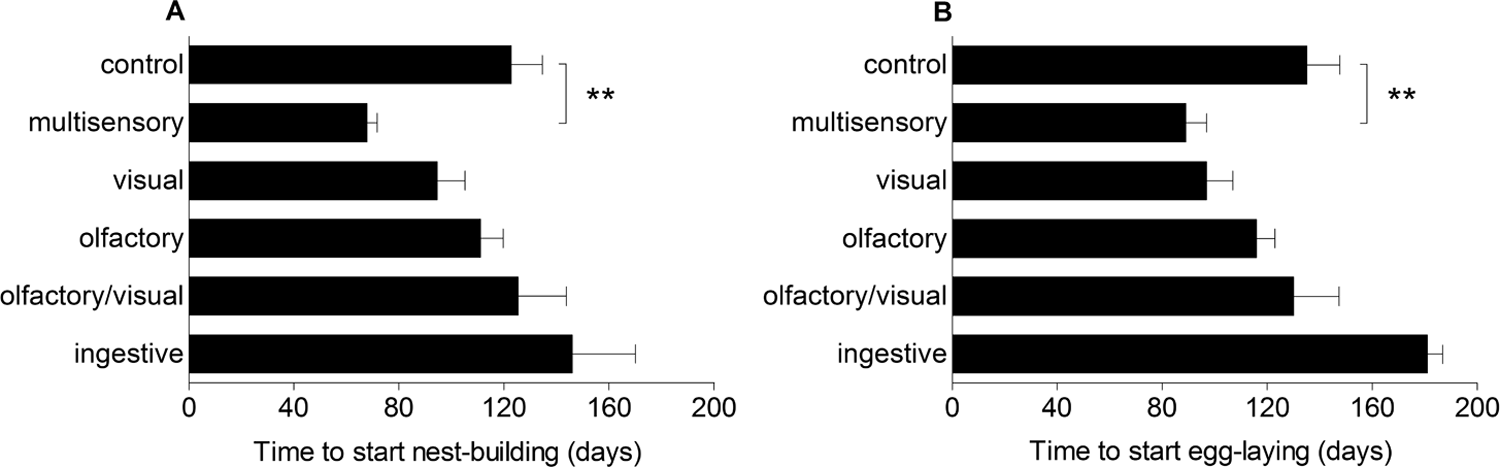
Influence of the different cues on breeding activity. The multisensory cue of fresh green vegetation significantly advanced nest building (A) and egg laying (B) compared to controls. Other stimuli had no significant effect (**p < 0.01).
Breeding activities lasted longest in the group that received the multisensory stimulus (204 days) compared to all other groups (range, 152-175 days). In all groups, breeding ceased at the end of July/beginning of August.
Number of Offspring
Across treatments, there was a significant correlation between the onset of nest building and the number of hatchlings (r = −0.47, p = 0.010, R2 = 0.221, N = 29) but not the number of fledglings (r = −0.357, p = 0.057, R2 = 0.127, N = 29). Thus, the earlier the birds started nesting, the more offspring they produced (Fig. 3).
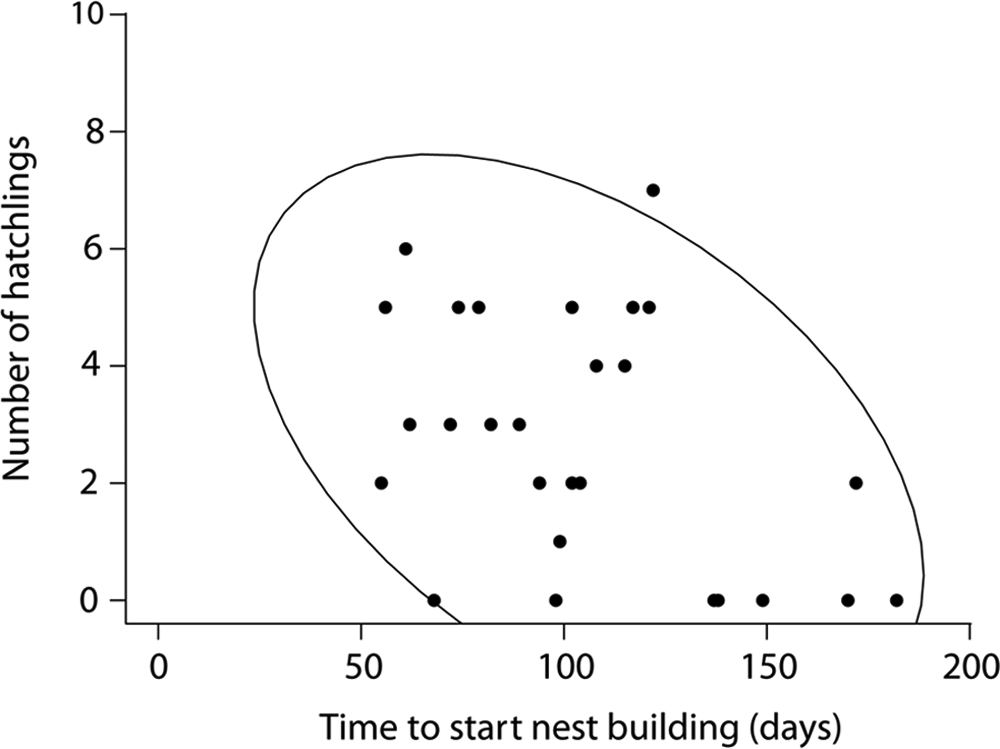
Correlation of offspring number and onset of nest building showing 90% confidence ellipse. The latency to start nest building was significantly negatively correlated with the number of hatchlings (r = −0.47, N = 29, p = 0.010).
Discussion
The present experimental study is, to the best of our knowledge, the first to examine the role of the different sensory components of an effective non-photic cue in stimulating advanced breeding activities in a photoperiodic bird. Our data show that birds must integrate and process multiple sensory stimuli represented by green vegetation in order to obtain reliable information about environmental conditions.
We found that when birds had direct access to fresh green vegetation, all females started building their first nest within a few days. In contrast, the advanced breeding activities in those groups that received either the visual or the olfactory stimulus were not significantly different from controls and resulted from the response of single females. Because the experimental setup was designed to keep the birds in groups rather than in pairs, it can be argued that social interactions might have contributed to the advanced breeding activities. However, this should have affected all groups equally, which was obviously not the case. Moreover, it can be assumed that in the natural environment, interindividual interactions play an important role in initiating early breeding events. Birds live in close proximity to each other, and neighboring nests are only a few meters apart. Also, it seems unlikely that the birds’ response to fresh green vegetation resulted from a carryover effect because experimental treatments differed between successive years (Table 1), and laying dates of the first egg were not influenced by year. Further, the environmental conditions in the aviaries outside the breeding season were similar for all groups.
Green plant material is known to contain compounds that, when injected or ingested, induce reproductive activity in rodents (Friedman and Friedman, 1939; Negus and Pinter, 1966; Negus and Berger, 1977). In a study with montane meadow voles, for example, the substance 6-methoxybenzoxazolinone (6-MBOA) has been identified as an active ingredient of monocotyledone plants (Sanders et al., 1981). While feeding on fresh green wheat leaves was shown to have an effect of advancing photostimulated ovarian growth in white-crowned sparrows (Ettinger and King, 1981), no single compound is yet known to trigger breeding activities in birds. In our experiment, birds that received the ingestive stimulus (plant extracts) did not initiate breeding earlier than control birds. Several technical reasons could have contributed to the failure of the birds to respond because the uptake of active compounds by the birds might have been simply too low. Also, the daily alternation of the type of plant extract could have played a role. Nevertheless, overall, it appears that while none of the sensory stimuli presented on its own elicited a significant response in females, the simultaneous processing of the different modalities enhanced their behavioral performance.
In most temperate zone species, increasing day lengths usually have a marked effect on the hypothalamo-pituitary-gonadal (HPG) axis, leading to an increased rate of gonadal recrudescence (Sharp, 2005). The rate of gonadal maturation in relation to photoperiod varies between species because of different thresholds for photostimulation. While in Japanese quail, the transition from nonstimulatory to stimulatory photoperiods is rapid and a critical day length can easily be defined, this process is more complicated in the European starling because gonadal growth occurs over a wider range of photoperiods (reviewed in Dawson, 2008). Gonadal growth rates are known to differ between males and females. Long day lengths are obviously stimulatory for both sexes but, whereas males under these conditions always reach full gonadal development, it is thought that the final stage of ovarian maturation in many species strongly depends on supplementary cues (Ball and Ketterson, 2008; Caro et al., 2009). In female great tits, maximum follicular development does not occur until females have started nest building (Silverin et al., 1989).
In the present study, first nest-building activities by females were observed 1 week after the winter solstice, and the first egg was laid 4 weeks later, corresponding to 17 min of increasing photoperiod. In previous years, breeding activities were even more advanced, and egg laying occurred before or coincided with winter solstice (Leitner et al., 2003; Voigt et al., 2007), although those eggs failed to develop (Voigt et al., 2007). These data strongly suggest that the initial stages of gonadal maturation, at least in female canaries, must have commenced well before winter solstice, when days were still short. One possibility is that after regaining photosensitivity in autumn, females reactivated a slow but constant gonadal growth, which was accelerated by introducing the fresh green vegetation by the end of November, and final ovarian development might have occurred when nest building started in early January. Related to this, it was found in an experiment with domesticated canaries (S. canaria) that photostimulation alone was not sufficient to induce full gonadal maturation in females, which indicates their sensitivity to environmental supplementary cues (Bentley et al., 2003). A slight but progressive gonadal growth during fall and winter has also been reported for female but not male white-crowned sparrows (Farner et al., 1966; Kern, 1972). Alternatively, we can assume that Island canaries possess a circannual rhythmicity of reproductive timing, on which both photic and non-photic cues act as synchronizers or modifiers (Gwinner, 1986; Visser et al., 2010). For free-living animals, photoperiod is thought to be the most important zeitgeber. However, it cannot be excluded that such a circannual rhythm is equally well entrained to environmental cues other than light.
Although the neural processing of non-photic cues is not yet understood, these cues must ultimately exert their effects on the gonadotropin-releasing hormone (GnRH) system in order to stimulate gonadal growth. It was recently shown in Japanese quail that photostimulation results in increased levels of thyrotropin in the pars tuberalis of the pituitary, which in turn triggers the release of GnRH from the hypothalamus (Nakao et al., 2008). Supplementary cues activate a number of different sensory systems such as the visual, olfactory, and somatosensory systems, which are thought to consist of independent pathways. In order to simultaneously process such diverse signals, information must be integrated at some point. It was recently found in barn owls that the main visual pathway (tectofugal pathway) processes not only visual but also auditory information, suggesting a multisensory function (Reches and Gutfreund, 2009). Therefore, integration areas could be located at a lower level such as the thalamus or higher up in the telencephalon. Both brain regions are targets of somatic projections (Delius and Bennetto, 1972), and the latter is also involved in olfaction (Rieke and Wenzel, 1978). However, the connection between those pathways and the HPG axis still remains elusive.
Footnotes
Acknowledgements
We are grateful to Roswitha Brighton for help with plant extract administration and animal care and Sonja Bauer and Andrea Peter for bird care. We further thank Wolfgang Goymann for discussion of the experimental design and Barbara Helm and Manfred Gahr for comments on the manuscript. The research was funded by the Max Planck Society, Germany.
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.