
Abstract
The Organic-Walled Microfossils (OWM), especially acritarchs, are essential in understanding eukaryotes’ early evolution and diversification in the Proterozoic Era. The study reports the fine-scale morphological features of the complex acanthomorphic acritarchs recovered from the Owk Shale of the Kurnool Group, Cuddapah Supergroup, South India. The acritarchs include Weissiella grandistella, Galeasphaeridium bicorporis, Galeasphaeridium sp., Germinosphera bispinosa and Tanarium tuberosum, which are characteristic of the Ediacaran age. Morphological features of all the recovered fossil forms were studied under high-power binocular microscopes and a Confocal Laser Scanning Microscope (CLSM), a new, non-destructive, non-intrusive method for detailed cellular morphology and anatomy, providing insight into three-dimensional morphology at micron-scale resolution. Many features were left unrecognised under the Transmitted-Light Microscopy (TLM). The present acanthomorphic acritarch assemblage of the Owk Shale closely resembles the Ediacaran Complex Acanthomorphic Palynomorphs (ECAP), which are of Ediacaran age. The rise in the complexity and diversity of the morphological features has been documented in the complex acanthomorphic acritarchs from the late Neoproterozoic sedimentary successions worldwide. The new assemblage strengthens the age of the Kurnool Group in favour of the Ediacaran. It adds information about the presence of the ECAP in the southern part of the Indian subcontinent.
INTRODUCTION
Diversity among the Organic-Walled Microfossils (OWMs) ranges from smooth-walled sphaeromorphs to forms bearing processes with distinct morphological ornamentations, reflecting the adaptations that enable organisms to evolve and survive under different environmental conditions. Acanthomorphic acritarchs are an essential component for understanding the biostratigraphy and evolutionary dynamics of marine eukaryotes after the Marinoan global glaciation since they are amongst the most prevalent eukaryotic fossils in early Ediacaran strata (Golubkova, 2023; Liu et al., 2013; Ouyang et al., 2021). Acanthomorphic acritarchs with spines or processes are the most common eukaryotic fossils of the late Neoproterozoic realm (Anderson et al., 2017; Golubkova et al., 2010; Grey, 2005; Knoll, 1992; Sergeev et al., 2011; Tiwari & Knoll, 1994; Vidal, 1990; Zhang et al., 1998). The Ediacaran acanthomorphic biotas are distributed between the interval from 630 to 540 Ma (Golubkova, 2023; Golubkova et al., 2015; Grazhdankin et al., 2020; Liu et al., 2013; Ouyang et al., 2021; Shen et al., 2005; Xiao et al., 2022; Zhou et al., 2017). Ediacaran acanthomorphic acritarchs are significant indicators of the evolutionary pattern of marine eukaryotic forms. Such acritarchs have been reported from different parts of the world which, includes: South China, Australia, Eastern Europe, Siberia, India, Svalbard China, Australia and Norway (Burzin, 1994; Golubkova, 2023; Golubkova et al., 2015; Grey, 2005; Joshi & Tiwari, 2016; Knoll, 1992; Knoll & Ohta, 1988; Liu & Moczydlowska, 2019; Liu et al., 2013; Moczydlowska, 2016; Moczydlowska & Nagovitsin, 2012; Moczydlowska et al., 1993; Ouyang et al., 2021; Shukla & Tiwari, 2014; Sokolov, 1997; Tiwari & Knoll, 1994; Veis et al., 2006; Vidal, 1990; Volkova et al., 1979; Vorob’eva et al., 2009; Willman & Moczydlowska, 2008; Willman et al., 2006; Xiao, 2004; Yin, 1987, 1999; Yin & Li, 1978; Yin & Liu, 1988; Yuan & Hofmann, 1998; Zang & Walter, 1992; Zhang et al., 1998; Zhou, 2007; Zhou et al., 2001). All these acritarch assemblages were studied under the high-power binocular microscopes. They originated during the Ediacaran time and spread worldwide rapidly, pre-dating the appearance of Ediacara-type bilaterian metazoans, and then became extinct before the end of the Neoproterozoic Period. The present study demonstrates the first comprehensive documentation of the well-preserved, distinct, organic-walled acanthomorphic acritarchs from the Owk Shale of the Kurnool Group, south India. The newly recovered acanthomorphic assemblage includes: Weissiella grandistella, Galeasphaeridium bicorporis, Galeasphaeridium sp. Germinosphera bispinosa and Tanarium tuberosum. These acanthomorphic acritarchs are studied under the optical microscope to document their complex morphological features, and under the Confocal Laser Scanning Microscope (CLSM) to document additional morphological features, that is, fine-scale details that were unrecognised under the optical microscope. The CLSM study provides insights into cellular complexity at the micron level over evolutionary timescales. The documented entities are very significant in terms of understanding the evolution of multi-cellular life and the atmospheric-ocean system of the Ediacaran Period. This study further substantiates the Ediacaran age of the Owk Shale of the Kurnool Group. It supports the theory of the possible extension of the Kurnool Group of sedimentation up to the Ediacaran, or possibly up to the early Palaeozoic.
GENERAL GEOLOGY AND AGE
Stratigraphic framework
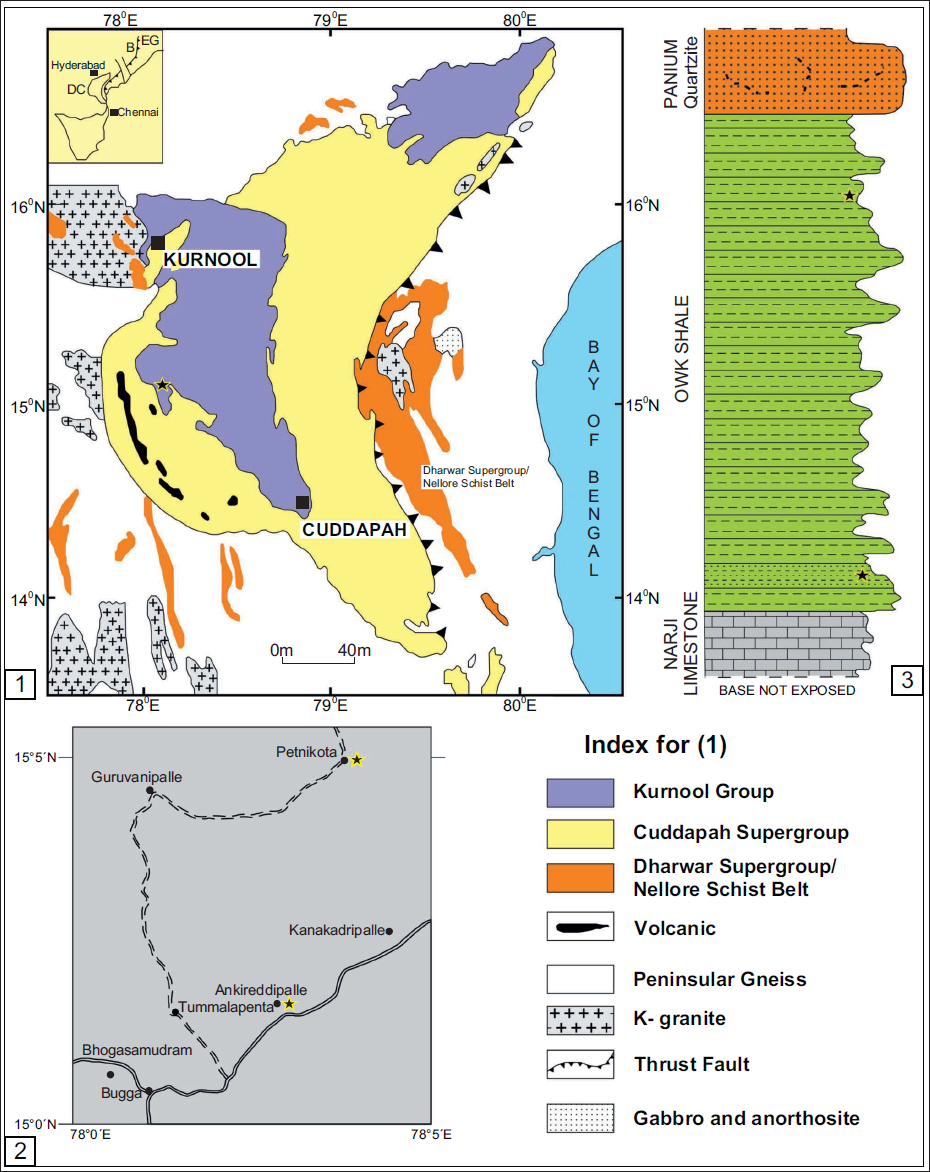
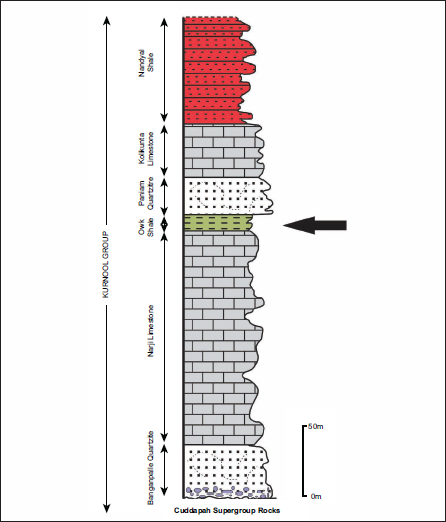
The Cuddapah Supergroup is a prominent Proterozoic sedimentary basin that exhibits a distinct N-S trending crescent-shaped geometry in the southern part of the Indian subcontinent (Figure 1). The Eparchaean unconformity marks the southern and western boundaries of the Cuddapah Basin and overlies the granite-greenstone terrain of the Eastern Dharwar Craton (EDC). The eastern part is concave and is marked by the prominent boundary thrust parallel to the Nellore Schist Belt (NSB), the Eastern Ghats Mobile Belt (EGMB), and the East Coast of India (Nagaraja et al., 1987). The first comprehensive documentation of the lithological record was provided by King (1872). Later, substantial revisions and additions were contributed by Meijerink et al. (1984) and Nagaraja et al. (1987). The Cuddapah Supergroup has been stratigraphically further subdivided into four groups: Papaghani, Chitravati, Nallamalai, and Kurnool. The Kurnool Group is the youngest group of the Cuddapah Supergroup, occupying a large area and deposited with an angular unconformity. The Kurnool Group has been further subdivided into six well-defined formations in stratigraphic order: Banganpalle Quartzite, Narji Limestone, Owk Shale, Paniam Quartzite, Koilkuntla Limestone, and Nandyal Shale. The thickness of these units is approximately 450 metres (Nagaraja et al., 1987; see Figure 2 and Table 1).
(1) Generalised geological map of the Kurnool Basin (after Mishra, 2011), inset: Bhandara Craton and DC – Dharwar Craton separated by Godavari Graben and EG – Eastern Ghat. (2) Geographical localities of the fossil-yielding areas in Ankireddipalle village, Kurnool district, Andhra Pradesh (marked by a star). (3) Lithostratigraphic succession was exposed at Ankireddipalle village, showing the Narji Limestone, Owk Shale, and Paniam Quartzite. Stars indicate the levels of carbonaceous fossils.
Generalised lithostratigraphic succession of the Kurnool Group, lying above the Cuddapah Supergroup rocks (after Nagaraja et al., 1987). The arrow shows the studied horizons.
Generalised lithostratigraphy of the Kurnool Group (after Nagaraja et al., 1987).

The deposition of the Kurnool Group took place in two cycles of quartzite-limestone-shale. The basement of the Kurnool Group consists of the Banganpalle Quartzite, which the massive, flaggy Narji Limestone overlies. The buff, purple and khaki-coloured Owk Shale unit follows it. These three litho-units constitute the first cycle of deposition; the second cycle of sedimentation is marked by the occasional conglomerate at the base of the Paniam Quartzite, overlain by siliceous to flaggy Koilkuntla Limestone, which gradually passes into the purple-coloured Nandyal Shale.
Age
The basal unit of the Kurnool Group, Banganpalle Quartzite, is a diamond-bearing unit. The source of these diamonds is considered to be the Vajrakarur Kimberlite, which has been dated at 1140 Ma and occurs nearby (Crawford & Compston, 1973). Based on this evidence, the Kurnool Group sedimentation is believed to be younger than 1140 Ma. However, Dongre et al. (2008) found the limestone xenoliths in the adjoining Siddanpalle Kimberlite. During the time of the Kimberlite emplacement, it is considered that there were only two limestone-bearing horizons present in the vicinity, that is, the Bhima Group and Kurnool Group (~50 km apart). It was suggested that the limestone xenolith may be remnants of either of the limestone sequences, and the Late Mesoproterozoic age (>1,090 Ma) was proposed for the Kurnool Group. Raman and Murthy (1997) studied both shale units of the Kurnool Group and proposed minimum and maximum ages of 500 Ma (K/Ar) and 980 Ma (Rb/Sr), respectively. Wabo et al. (2022) conducted Palaeomagnetic and radiometric studies on the Narji Limestone using the 40Ar/39Ar dating method, yielding an age of ~800 Ma. Gururaja et al. (2000) proposed the Pc-C boundary within the Kurnool Group based on certain fossils. Currently, the Kurnool Group is considered Neoproterozoic in age (Kumar et al., 2021, 2022; Sharma & Shukla, 1999, 2012, 2016; Shukla et al., 2020).
MATERIAL AND METHODS
The present study is based on the microfossils recovered from the Owk Shale of the Kurnool Group in southern India. The samples of the Owk Shale were collected from Ankireddipalle village (15°07’; 78°03’) in the Kurnool district of Andhra Pradesh, southern India. At this spot, the thickness of the section is 15 m, and it is exposed on the eastern part of the hillock ∇1445. The shale samples were macerated by refined palynological maceration techniques devised by Grey (1999). The organic residue obtained was mounted on a glass slide using PVC and Canada Balsam (R.I. = 1.5). The illustrated specimens are deposited in the repository of BSIP, Lucknow under the wide statement number 1668.
RESULTS
Optical microscope (OM)
About 60 palynological slides of the Owk Shale fossils were examined under a transmitted-light microscope (Olympus BX53) at 40× and 100× (oil immersion) to document the simple morphological details of the present microfossils. All the measurements of the microfossils described in this article were made using the Cell Cense Standard Software, and photographs were taken with the microscope-mounted software-supported Olympus DP 26 digital camera. England Finder coordinates and slide numbers are provided in parentheses in each figure caption for all the described microfossil specimens in this article. All the slides containing demonstrated specimens are deposited in the Birbal Sahni Institute of Palaeosciences, Lucknow, India, repository under Museum Statement Number BSIP-XXXX.
Confocal Laser Scanning Microscope (CLSM)
The CLSM facility at BSIP was used with standard protocols (Schopf et al., 2006) to obtain additional fine-scale morphological information at high spatial resolution, and 3D images of the microfossils were also generated. Three-dimensional Confocal fluorescence imaging was acquired on a Leica TCS SP8 Confocal Laser scanning biological microscope system equipped with two Melles Griot lasers, a 448 nm 20 mW-output argon ion laser and a 633 nm 10mW-output helium-neon laser (Melles Griot, Carlsbad, CA). The images were obtained using a 100× oil-immersion objective (Numerical Aperture = 1.4). The images acquired by this system were subsequently processed and examined on the LAS-X imaging software associated with the CLSM.
SYSTEMATIC PALAEONTOLOGY
The present study reports the presence of W. grandistella, G. bicorporis, Galeasphaeridium sp., T. tuberosum, and Germinosphaera bispinosa in the Owk Shale of the Kurnool Group, India. Morphological features are based on observations made under the microscope, that is, OM and CLSM.
Genus Galeasphaeridium Vorob’eva et al., 2009
Type Species Galeasphaeridium sp. Vorob’eva et al., 2009
Galeasphaeridium sp.
(Plate 1, Figures 1 and 2)
Material: One complete specimen has been documented in this study.
Stratigraphic Horizon: Owk Shale, Kurnool Group.
Dimensions: The recovered specimen’s vesicle is 124.25 µm long and 74.01 µm wide. The processes of this specimen range from 3.2 µm to 16.8 µm in length and 5.7 µm to 13.8 µm in width.
Photomicrographs of Galeasphaeridium sp. (1, 2); Tanarium tuberosum (3, 4), Germinosphaera bispinosa (5-8) from the Owk Shale, Kurnool Group under transmitted light Optical Microscope (OM) and Confocal Laser Scanning Microscope (CLSM). Optical photomicrographs (1, 3, 5, 7); Confocal Laser Scanning micrographs (2, 4, 6, 8). 2, 4, 6, 8 are recorded under grey CLSM imagery. (1, 2) Galeasphaeridium sp., BSIP slide no: 16585, K51/2; (3, 4) Tanarium tuberosum BSIP slide no: 16583, K32/4; (5-8) Germinosphaera bispinosa BSIP slide no: 17641, S41/3, 17642, E31/4. Scale Bar = 20 µm. Arrows show different processes and spines.

Photomicrograph of Weissiella grandistella (1-9) from Owk Shale, Kurnool Group under transmitted light Optical Microscope (OM) and Confocal Laser Scanning Microscope (CLSM). Optical photomicrographs (1, 3, 5), Confocal Laser Scanning micrographs (2, 4, 6-9). 2, 4, 6-9 are recorded under the grey colour CLSM imagery. (1-9) Weissiella grandistella, (1) BSIP slide no16583, S18/1; (3) BSIP slide no16583, N25/2; (5) BSIP slide no16585, M33/4; (8, 9) enlarged view of Weissiella grandistella showing the presence of small spines all over the body. Scale bar=20µm for (1-7); Scale bar = 10 µm (8, 9); Arrows showing the processes and spines.

Photomicrographs of Galeasphaeridium bicorporis from the Owk Shale, Kurnool Group under transmitted light Optical Microscope (OM) and Confocal Laser Scanning Microscope (CLSM). Optical photomicrographs (1, 3, 5, 7), Confocal Laser Scanning Micrographs (2, 4, 6, 8); 2, 4, 6, 8 are recorded under grey colour CLSM imagery. (1-8) Galeasphaeridium bicorporis; (1) BSIP slide no. 17643, S25/3; (2) BSIP slide no17644, O35/4; (5) BSIP slide no. 16585, H37/3; (7) BSIP slide no16580, K28/1; Scale bar = 20 µm.

Microscopic Observations: The specimen is a translucent, ellipsoidal vesicle with 3-4 processes arising from its wall. The processes are complete. The surface has some conical spine-like features distributed over the body (Plate 1, Figure 1).
CLSM Observations: The CLSM study shows that spines are present throughout the body. The processes appear incomplete with broken distal tips.
Remarks: The specimen reported in the present study closely resembles Galeasphaeridium oviscoris as described by Vorob’eva et al. (2009), but is smaller and has less-developed processes, as observed in the type specimen. Morphological features make it easy to compare with the G. oviscoris.
Occurrence and Stratigraphic range: Ediacaran in age. Kel’tminskaya-1 borehole, Northeastern margin of the East European Platform (EEP) (Vorob’eva et al., 2009).
Genus Tanarium Kolosova, 1991, emend. Moczydlowska et al., 1993. Moczydlowska, 2005
Type species T. tuberosum Moczydlowska, 2005
T. tuberosum
(Plate 1, Figures 3 and 4)
Material: One complete specimen has been observed.
Stratigraphic Horizon: Owk Shale, Kurnool Group.
Dimensions: The recovered specimen has a vesicle of 91.43 µm in length and 70.47 µm in width. The conical processes are present on the surface and range in length from 5.1 µm to 8.8 µm; the base width ranges from 7 µm to 9 µm, with blunt or broken tips.
Microscopic Observations: Spheroidal vesicles have a few small conical-shaped processes distributed unevenly on the surface. The processes are short, unbranched, and hollow-conical, with short, pointed tips and folds on the body.
CLSM observations: The CLSM study confirms the processes’ connection with the vesicle; the processes are hollow and conical in shape (Marked by an arrow in Plate 1, Figures 3 and 4).
Remarks: The specimen reported here has conical spines attached to the vesicle.
Occurrence and Stratigraphic range: Early Ediacaran. Yakutia, Siberian Platform (Moczydlowska, 2016); the Kursov Formation (Moczydlowska, 1993); Khamaka Formation (Moczydlowska, 2016); Lena-Anabar Depression (Moczydlowska, 1993, 2005, 2015); Officer Basin, Australia (Willman & Moczydlowska, 2008); lower Ungoolya Group (Willman et al., 2006); Tanana Formation, Australia (Willman & Moczydlowska, 2008).
Genus Germinosphaera Mikhailova (1986), Butterfield in Butterfield et al., 1994
Type species G. bispinosa Mikhailova, 1986
G. bispinosa
(Plate 1, Figures 5–8)
Material: Two complete specimens have been recovered.
Stratigraphic Horizon: Owk Shale, Kurnool Group.
Dimensions: The recovered specimens range from 87.88 µm to 120.37 µm in length and 72.43 µm to 113.64 µm in width. The length of the processes ranges from 6.7 µm to –16.48 µm, and the width ranges from 9.03 µm to 13.29 µm.
Microscopic Observations: Spheroidal vesicle bearing one to two unbranched tubular processes extending freely from the vesicle wall. The distal termination of the process is broken. The process is hollow and communicates directly with the vesicle (marked by an arrow in Plate 1, Figures 5 and 7).
CLSM observations: The CLSM study shows that the processes are freely connected to the inner wall and are hollow (marked by an arrow in Plate, Figures 6 and 8).
Remarks: Butterfield et al. (1994) amended the species description to include specimens with one to four processes, which are distributed on the equatorial plane of the vesicle, and synonymised G. bispinosa and G. unispinosa into G. bispinosa (according to name priority). Loron et al. (2021) also advocated this, and we, too, are in favour of appropriately merging the two species.
Occurrence and Stratigraphic range: Dashka Formation, East Siberian Platform, Siberia (Mendelson & Schopf, 1992; Mikhailova, 1986); Svanbergfjellet Formation, Akademikerbreen Group Spitsbergen; Visingsö Group, Sweden (Butterfield et al., 1994); Ganga Basin, Bahraich Group, Avadh Formation, Ujhani Formation, (Prasad & Asher, 2001); Northern part of the Eastern European Platform, Vychegda Formation, upper assemblage, (Vorob’eva et al., 2009); Escape Rapids Formation and Grassy Bay Formation, Shaler Supergroup, Canada (Loron et al., 2019); rare Mesoproterozoic occurrence is also reported from the Fort Confidence Formation, Dismal Lakes Group, Canada (Loron et al., 2021).
Genus Weissiella Vorob’eva et al., 2009
Type species W. grandistella Vorob’eva et al., 2009.
W. grandistella
(Plate 2, Figures 1–9)
Material: Four well-preserved specimens have been recovered.
Stratigraphic Horizon: Owk Shale, Kurnool Group.
Dimensions: The specimens recorded in the present study have a vesicle diameter (without processes) of around 100–101 µm in length and 73–85 µm in width. The processes range from 5 µm to 13 µm in width and 3.5 µm to 14 µm in length. Small spines are present all over the body, ranging from 4 µm to 6 µm in length and 2 µm to 5 µm in width (Plate 2).
Microscopic Observations: Single-walled spherical, hollow vesicle having heteromorphic robust processes, tubular (marked by an arrow in Plate 2, Figures 1, 3 and 5), having a broad base with rounded or blunt tips. Conical processes (marked by a white arrow on the vesicle in Plate 2, Figure 1) have also been observed over the body. Clear micro laminations are visible in some processes (marked by an arrow in Plate 2, Figure 1).
CLSM Observations: The recovered specimens have been observed under the CLSM, which shows some fine fibril-like features present all over the body (Plate 2, Figures 2, 4, 6–9).
Remarks: The specimens reported in the present study resemble the characteristic generic features of Weisiella, but are short in size compared to the type specimen. The micro-laminated pattern is also not very well visible. However, spines all over the body have been seen under the CLSM (Plate 2, Figures 8 and 9), which helps compare with the type specimen.
Occurrence and Stratigraphic range: Commonly found in the Ediacaran successions of the world. Vychegda Formation; EEP (Vorob’eva et al., 2009); Doushantuo Formation, South China (Liu et al., 2013; Ouyang et al., 2021); Outer Krol belt, Lesser Himalaya (Shukla & Tiwari, 2014; Xiao et al., 2022).
Genus Galeasphaeridium Vorob’eva et al., 2009
Type Species G. bicorporis Vorob’eva et al., 2009
G. bicorporis
(Plate 3, Figures 1–8)
Material: Four specimens, two complete and two incomplete, were recovered. All the specimens were examined using optical and CLSM.
Stratigraphic Horizon: The Owk Shale, Kurnool Group.
Dimensions: The recovered specimens’ vesicles range from 62.75 µm to 158.48 µm in length and from 41.82 µm to 112.39 µm in width. The processes of these specimens range from 2.35 µm to 19.25 µm in length and 2.1 to 20.96 µm in width.
Microscopic Observations: The specimens are opaque to translucent, spherical to ellipsoidal vesicles with two conical processes at opposite poles (Plate 3, Figures 1, 5, 7). The processes arise from the base and taper, giving it an ogee-like appearance.
CLSM Observations: The specimens under the CLSM show the complete connection between the processes and vesicle, and also show that some processes have a circular opening at the tip (marked by an arrow in Plate 3, Figure 1). The vesicle surface does not appear smooth under CLSM but lacks any convincing structure to be recognised.
Remarks: The specimens reported in the present study resemble the type specimen reported by Vorob’eva et al. (2009) but are smaller in size than the reported ones. The process size is also small compared to the type specimen, and the morphology of these fossils, especially the presence of processes at opposite ends, makes them comparable with the type specimen.
Occurrence and Stratigraphic range: Kel’tminskaya-1 borehole, Northeastern margin of the EEP (Vorob’eva et al., 2009).
DISCUSSION
The Owk Shale of the Kurnool Group is a fossiliferous horizon that yielded several OWMs, including the Ediacaran Complex Acanthomorphic Palynomorphs (ECAP) assemblage (Shukla et al., 2020). The ECAP acritarchs are an important component of the Ediacaran Period and provide significant information about the stratigraphic position of the fossil-bearing strata. They are also helpful in global correlation. The palaeobiological studies conducted on the Owk Shale of sediments document both the prokaryotic and eukaryotic types of biological remains (Arya & Rao, 1979; Das Sharma et al., 1992; King, 1872; Kumar et al., 2021, 2022; Rajurkar, 1963; Salujha et al., 1972; Sharma, 2008; Sharma & Shukla, 1999, 2012, 2016; Shukla et al., 2020; Vijayam, 1968). In addition to the reported palaeobiological remains, new acanthomorphic acritarchs are documented in the present study. W. grandistella, G. bicorporis, Galeasphaeridium sp. G. bispinosa and T. tuberosum are typical ECAP, reported only from different Ediacaran successions worldwide. The unique aspect of the present study is that the recorded microfossils have been studied three-dimensionally using a CLSM to reveal additional morphological features of the recorded entities (Plate 2, Figure 8). The present study generates both greyscale and coloured imagery of all the recorded microfossils (Plates 1–3). These photographs also reveal how the biological remains were preserved under the sediments and depict their fine-scale morphological features at very high resolution. The presence of such advanced morphological entities in the Owk Shale supports an Ediacaran age for the shale.
BIOSTRATIGRAPHIC SIGNIFICANCE OF RECORDED ACANTHOMORPHS
The diverse acanthomorphic acritarch assemblage has been reported from different Ediacaran successions worldwide. The assemblage reported in the present study closely resembles that of Vorob’eva et al. (2009). The presence of acanthomorphic acritarchs in the Owk Shale of the Kurnool Group demonstrates that, along with other microfossils reported by Sharma and Shukla (2012, 2016), Shukla et al. (2020), and Kumar et al. (2021), a rich complex acanthomorphic assemblage is also present. A few acanthomorphs, namely Cavaspina aff. C. acuminate and Variomargosphaeridium aff. V. litoschum have already been reported (Shukla et al., 2020). The taxa reported in the present study vary in the shape and size of vesicles, as well as in their processes and spines. Vorob’eva et al. (2009) reported acanthomorphic acritarchs from the EEP, which is typically Ediacaran in age, and the present assemblage resembles them very closely. Presence of W. grandistella, G. bicorporis, Galeasphaeridium sp.? G. bispinosa and T. tuberosum indicates that the reported assemblage is Ediacaran in age. W. grandistella has been reported worldwide from different Ediacaran successions. Galeasphaeridium sp. reported in this article very closely resembles the G. oviscoris (Vorob’eva et al., 2009), although it lacks convincing spines and contains other generic morphological features of the Galeasphaeridium.
G. bispinosa is a long-ranging acanthomorph reported from the Early Mesoproterozoic to the Cambrian. However, the abundance of Obruchevella and other acanthomorphic acritarchs supports the idea that the Owk Shale is Ediacaran in age. Given that the worm burrows occur in the underlying Narji Limestone and the Ediacaran discs in the overlying Paniam Quartzite, the acanthomorphic acritarchs could also serve as a viable tool for stratigraphic correlation. This also solves the age conundrum of the Kurnool Group and supports an Ediacaran age for the Owk Shale of the Kurnool Group. The radiometric ages of the Kurnool Group are very meagre, and the palaeobiological data from the Owk Shale of the Kurnool Group show significant contrast with the radiometric ages and indicate that the sedimentation of the Kurnool Group might have extended up to the terminal part of the Neoproterozoic and may be extended up to the early Palaeozoic as well.
CONCLUSIONS
The present acanthomorphic assemblage from the Owk Shale of the Kurnool Group is the first comprehensive record of the Complex acanthomorphic Palynomorphs, which are of Ediacaran age. The three-dimensional microphotographs of the recorded microfossils were also generated using a CLSM, revealing fine-scale morphological features of the recovered microfossils that were previously left unrecognised under the OM. The recorded assemblage is also highly significant for its global correlation, as it closely resembles the EEP Assemblage and other acanthomorphic microbiota worldwide. New Data further strengthens the earlier inferences about the age of the Owk Shale of the Kurnool Group as Ediacaran.
Footnotes
Acknowledgements
YK thanks the Vice Chancellor and Head, Department of Geology, Central University of Tamil Nadu and Director, Birbal Sahni Institute of Palaeosciences, Lucknow, for providing the facilities and necessary permissions to publish this article (RDCC/2025-26/42). We sincerely acknowledge the support of Precambrian palaeobiology lab mates at BSIP. Ms Shivalee Srivastava and Ms Archana Sonker provided technical assistance for Confocal Laser Scanning Microscopy.
Contribution of Authors
YK and MS for sample collection; YK, MS, VKS and SG for writing of the manuscript. YK and VKS conducted the CLSM studies.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: We also thankfully acknowledge the financial support received under the DST-RFBR Grant (INT/RUS/RFBR/P-278).