
Abstract
The present study deals with the reconstruction of Early Permian floral diversity, palaeoenvironment, and depositional setting using a multiproxy approach involving morphotaxonomy, palynology and palynofacies analyses of a coal-bearing sequence of Kurasia Colliery, Chirimiri Coalfield, Son Basin, India. The megafloral assemblage is of moderate diversity and is characterised by the presence of three groups, that is, Cordaitales, Glossopteridales and Equisetales. Macrofloristically, Cordaitales are represented by a single leaf fossil genus, that is, Noeggerathiopsis (N. elongata, N. hislopi, N. minor and Noeggerathiopsis sp.) and two seed genera, namely Cordaicarpus karharbariensis and Samaropsis ganjrensis. Glossopteridales are represented by three genera- Gangamopteris, Glossopteris and a seed genus Alatocarpus. Gangamopteris is represented by five species, namely, G. angustifolia, G. cyclopteroides, G. major, G. rajaensis and Gangamopteris sp.; Glossopteris is represented by seven species- G. communis, G. decipiens, G. major, G. nautiyalii, G. raniganjensis, G. spatulata, and Glossopteris sp. Equisetales are represented by two genera- Paracalamites (Paracalamites sp.) and Raniganjia (R. bengalensis). The palynological analysis has revealed only one assemblage, dominated by the monosaccate genus Parasaccites, followed by Plicatipollenites and the bisaccate Scheuringipollenites, along with other marker taxa such as Crucisaccites and Callumispora. This palynoassemblage is well correlated with the Parasaccites-Scheuringipollenites palynoassemblage of the Upper Karharbari Formation of the Godavari Basin. The palynocomposition has affinities with Cordaitales, Glossopteridales, Coniferales and Filicales. The megafloral and palynofloral composition suggests that the studied section represents Upper Karharbari-Lower Barakar transitional vegetation. The presence of large leaves in these sediments supports the existence of warm, temperate, and humid climatic conditions conducive to coal formation. The palynofacies study shows the dominance of structured phytoclasts, suggesting that the studied sediments were deposited in low-energy, suboxic-dysoxic forest swamp settings.
Keywords
INTRODUCTION
The sedimentary strata that were deposited across various regions of peninsular India during the Permo-Carboniferous upto Cretaceous periods are collectively known as the Gondwana deposits (Veevers & Tewari, 1995). These deposits represent a significant phase in the geological history of the region, marking the reinitiation of sedimentation during the? late Carboniferous/earliest Permian, after a prolonged depositional hiatus since the Proterozoic Era, in several isolated basins– Son-Mahanadi, Damodar, Rajmahal, Satpura, and Wardha-Godavari. These basins have significant economic value due to the presence of huge coal reserves and are aligned along three major linear belts that follow the present-day river valleys of the Narmada, Son, Damodar, Pranhita, Godavari and Mahanadi; and two hill ranges, namely Satpura and Rajmahal. The coal reserves in these basins, part of the Lower Gondwana Group, are primarily confined to specific formations: the Karharbari, Barakar, and Raniganj formations, and are absent in the Talchir and Barren Measures formations. Among these basins, the Son-Mahanadi Basin, located in the central and eastern parts of India, is one of the most significant. It has thick and extensive sedimentary sequences rich in coal-bearing deposits, making it an important coal-producing basin of the country. The Son-Mahanadi Basin encompasses around 18 major coalfields, Athgarh outliers, Bisrampur, Chirimiri, Hasdo-Arand, Ib River, Jhilimili, Johilla, Korba, Korar, Lakhanpur, Mand-Raigarh, Sendurgarh, Singrauli, Sohagpur, Sonhat, Talcher, Tatapani-Ramkola and Umaria, along with their adjoining regions, which together form the primary sedimentary deposits of the Son-Mahanadi Basin (Singh & Saxena, 2015).
The coal-bearing sedimentary sequences of these coalfields hold significant importance in understanding the evolution and proliferation of the ‘Glossopteris Flora’ across the Gondwanan continents. This flora had appeared in the southern continents during the earliest Permian, after deglaciation following the Late Palaeozoic Ice Age. Its primary macrofloral occurrence has been well documented across all Gondwanan landmasses beginning from the Asselian (Chandra, 1992; McLoughlin, 2001; McLoughlin et al., 2005 and references therein). The subsequent diversification of this flora throughout the Permian Period has been extensively studied by numerous researchers (both macrofloral and palynological aspects), particularly in various coalfields and adjoining regions of the Son Basin. These include Korba (Saxena et al., 2013; Singh et al., 2011a, 2012a; Vijaya & Sinha, 2005), Tatapani-Ramkola (Kar & Srivastava, 2003; Saxena, Singh, Cleal, et al., 2019; Singh et al., 2011b, 2012b; Tripathi et al., 2012), Singrauli (Pillai et al., 2025; Saxena, Singh, Murthy, et al.; Saxena, Singh, Shabbar, et al., 2016; Saxena, Singh, Murthy, et al., 2019; Saxena et al., 2021, 2022; Singh & Saxena, 2015; Singh et al., 2016, 2017; Vijaya et al., 2012), Sohagpur (Agnihotri et al., 2016; Chandra & Srivastava, 1982; Feistmantel, 1882), Mand-Raigarh and Johilla (Pillai et al., 2018). However, as compared to the comprehensive palaeobotanical studies conducted in the aforementioned coalfields, the Kurasia Colliery of the Chirimiri Coalfield has received no attention so far. More recently, Gupta et al. (2025) have reported and discussed in detail the upper Talchir macroflora from the outcrop exposures in and around the Chirimiri Coalfield.
The present study aims to reconstruct the Permian palaeoenvironment and palaeovegetation of the Kurasia Colliery in the Chirimiri Coalfield through a multiproxy analysis comprising macroflora, palynology, and palynofacies. By analysing coal-bearing sediments, the study also seeks to assess and understand palaeofloral diversity and its dynamics, and to establish a biostratigraphic age of the succession. The study is significant in understanding and reconstructing vegetational dynamics across the Sakmarian and Artinskian times and correlating these findings with contemporaneous strata from other Gondwana basins in peninsular India.
GEOLOGICAL BACKGROUND
The Son Basin, a part of the larger Son-Mahanadi Master Basin, is one of India’s most significant Gondwana basins due to its rich coal reserves. It is located in the central and eastern part of India, and is recognised as one of the largest intracratonic rift basins on the Indian Peninsula. Some significant coalfields of the Son Basin are: Korba, Tatapani-Ramkola, Singrauli, Sohagpur, Johilla, Mand-Raigarh and Chirimiri. The Chirimiri Coalfield, located in the Eastern Bundelkhand Plateau region, is one of the most developed and productive coalfields in Chhattisgarh State. Chirimiri Coalfield spans between latitudes 23°8′ N and 23°15′ N, and longitudes 82°17′ E and 82°25′ E. This coalfield includes seven active mining collieries: Kurasia, Chirimiri, New Chirimiri Pondi Hill (NCPH), West Chirimiri, Duman Hill, North Chirimiri, and Koriya. Among these, Kurasia Colliery, situated in the south-central part of the coalfield, is considered one of the most significant coal mines of the region.
In this region, the Permian-age rock sequences belong to the Talchir and Barakar formations. The Barakar sediments appear as steep escarpments over the Talchir outcrops. Notably, the Barren Measures and Raniganj formations are absent, probably owing to either non-deposition or subsequent erosion (Raja Rao, 1983). Overlying the Barakar Formation are the Deccan Traps, suggesting that Mesozoic sedimentary layers are also missing from the stratigraphic sequence of the area.
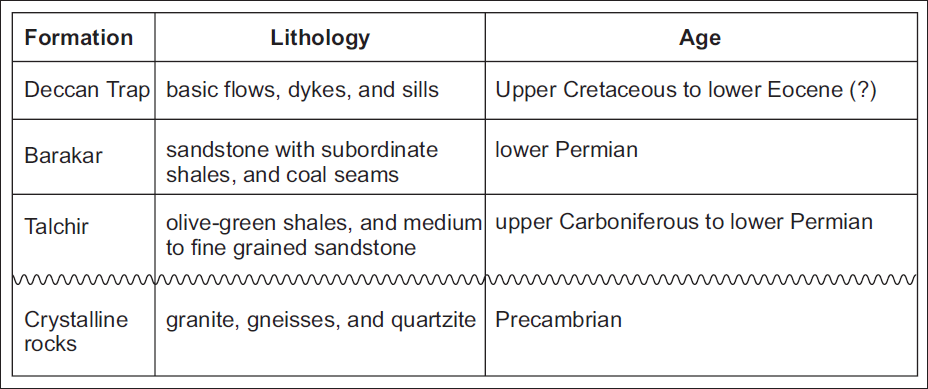
The Barakar Formation is predominantly composed of shale, coal, and light grey coarse-grained sandstone (Raja Rao, 1983). The shale, typically grey in colour, is relatively minor within the overall sedimentary sequence. In the present study, the focus is on the lithologically designated Barakar Formation exposed in the Kurasia Colliery. The general stratigraphical framework of the Chirimiri area is summarised in Figure 1.
Stratigraphic succession of Chirimiri area, Son Basin, Chhattisgarh (after Raja Rao, 1983).
MATERIALS AND METHODS
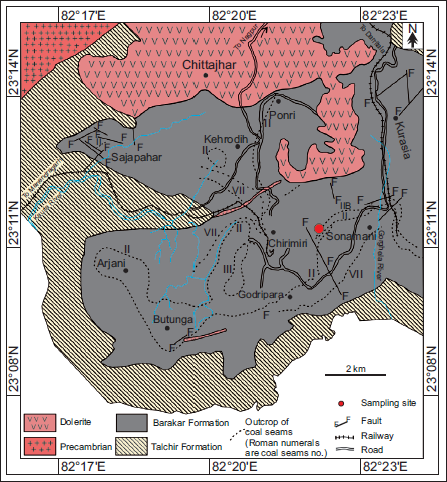
The megafossil assemblage described in this study was procured from the exposed sequence of the Karharbari/ Barakar Formation, Kurasia Colliery (23º11′11.8′′N and 82º21′ 24.0′′E) within the Chirimiri Coalfield, Chhattisgarh (Figure 2). Around 196 macrofossil specimens were procured from the fine carbonaceous shale units intercalated between the coal seams and shale/siltstone partings from a ~13 m thick exposure (Figure 3A and 3B). The samples were collected by one of the authors (KJS). The fossil-bearing horizons belong to the lowermost sequence of the Barakar Formation (lithologically designated) exposed in the Colliery. The complete sequence in the Colliery is about 86 m thick, comprising five coal seams, namely Seam 5 (Sonawari Seam), Seam 4, Seam 3, Seam 2 and Seam 1, with intermittent partings of sandstone, siltstone, grey shales and carbonaceous shale. The samples are procured from the shale partings above the Sonawari seam.
Location and geological map of Chirimiri area, Son Basin, Chhattisgarh, India (after Raja Rao, 1983).
(A) Field photograph of coal-bearing sequence, Kurasia Colliery, Chirimiri Coalfield, Chhattisgarh, Son Basin. (B) Litholog of the studied section, Kurasia Colliery, Chirimiri Coalfield, Chhattisgarh, Son Basin.

The macrofloral remains were found preserved as impressions and compressions on carbonaceous shale. These specimens were examined by analysing morphological details for their taxonomic evaluation (Chandra & Singh, 1992; Chandra & Surange, 1979; Gupta et al., 2025 and references therein).
Five palynological samples were collected at various intervals from the grey and carbonaceous shale units between the Sonawari Seam (Seam 5) and Seam 4. The samples were first cleaned and then manually crushed using a mortar and pestle. Approximately 40–50 g of the crushed material from each sample was subjected to chemical processing to extract pollen and spores using standard maceration techniques (see Gupta et al., 2023 for detailed description). However, a standard non-oxidative maceration technique was used for the palynofacies sample preparation (Aggarwal et al., 2019). For the palynofloral analysis, at least 100 palynomorphs were counted per sample. In contrast, at least 500 organic particles were counted in each sample for palynofacies analysis to obtain a sizeable amount of organic matter. Prepared slides were analysed under an Olympus BX-61 microscope equipped with a DP-20 camera, using Cell-A software for detailed observation and image documentation.
Repository: All the macrofloral specimens and palynological and palynofacies slides pertaining to this study are deposited in the museum repository of the Birbal Sahni Institute of Palaeosciences (BSIP), Lucknow, vide Statement No. 1672 and macrofloral specimen numbers 42524-42539 and slide numbers 17645-17663.
RESULTS
Megafloral observations
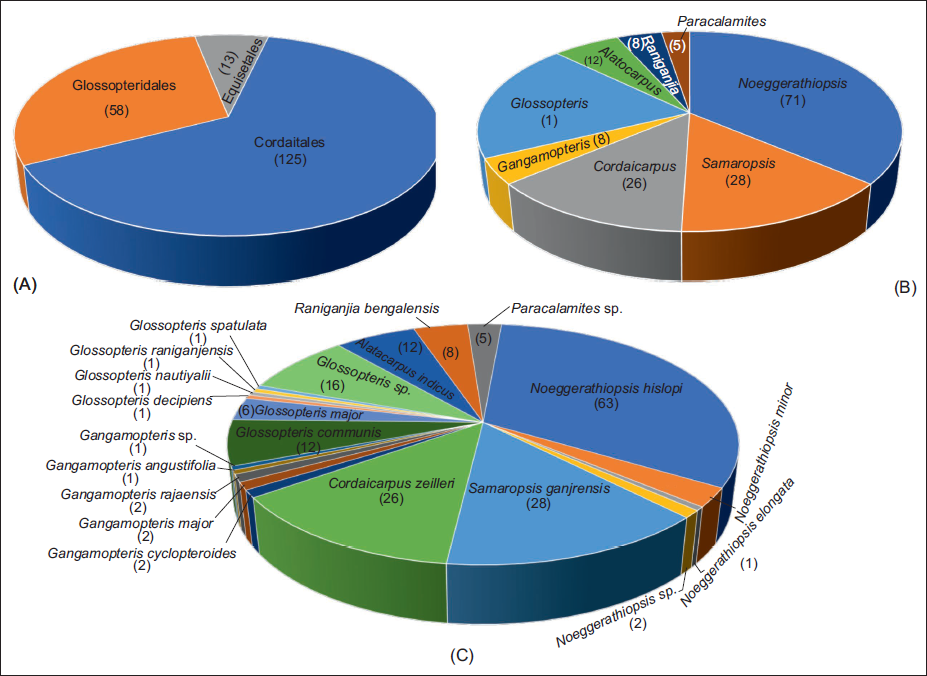
The present macrofloral assemblage exhibits a rich diversity of midrib-less leaf morphotypes and is characterised by the Noeggerathiopsis-Glossopteris-Gangamopteris assemblage. Cordaitales dominate the assemblage and are represented by three genera. Leaf genus Noeggerathiopsis is the most abundant, with 71 specimens identified, belonging to four species (N. elongata, N. hislopi, N. minor and Noeggerathiopsis sp.) and two seed genera comprising Cordaicarpus (C. karharbariensis) and Samaropsis (S. ganjrensis). Glossopteridales are represented by three genera- leaf genus Glossopteris comprises 38 specimens, belonging to seven species (G. communis, G. decipiens, G. major, G. nautiyalii, G. raniganjensis, G. spatulata and Glossopteris sp.). The genus Gangamopteris includes seven specimens, belonging to five species (G. angustifolia, G. cyclopteroides, G. major, G. rajaensis and Gangamopteris sp.). One seed taxon of this order, Alatocarpus is represented by Alatocarpus indicus. Equisetalean are represented by two genera- Paracalamites, represented by five specimens, and Raniganjia with one species, Raniganjia bengalensis (eight specimens). A few selected plant fossil specimens from this assemblage are illustrated in Plate 1, and their quantitative distribution is shown in Figure 4. As the assemblage comprises already known taxa, the detailed descriptions of each taxon, along with their systematic palaeobotany and photographic illustrations, will be published elsewhere (MS in prep).
Photographs of megafossil assemblage recovered from an Early Permian coal-bearing sequence of Kurasia Colliery, Chirimiri Coalfield, Chhattisgarh. (Scale bar is 5 mm for all specimens). 1). Gangamopteris angustifolia, BSIP Museum Specimen No. 42528; 2). Gangamopteris rajaensis, BSIP Museum Specimen No. 42532; 3). Gangamopteris major, BSIP Museum Specimen No. 42536; 4). Noeggerathiopsis hislopi, BSIP Museum Specimen No. 42529; 5). Glossopteris communis, BSIP Museum Specimen No. 42533; 6). Glossopteris nautiyalii, BSIP Museum Specimen No. 42525; 7). Raniganjia bengalensis, BSIP Museum Specimen No. 42528; 8). Alatocarpus indicus, BSIP Museum Specimen No. 42536; 9). Cordaicarpus zeilleri, BSIP Museum Specimen No. 42536; 10). Samaropsis ganjrensis, BSIP Museum Specimen No. 42538; 11). Noeggerathiopsis elongata, BSIP Museum Specimen No. 42526.

Quantitative distribution of macrofloral remains recovered from an Early Permian coal-bearing sequence of Kurasia Colliery, Chirimiri Coalfield, Chhattisgarh, Son Basin.
Palynological observations
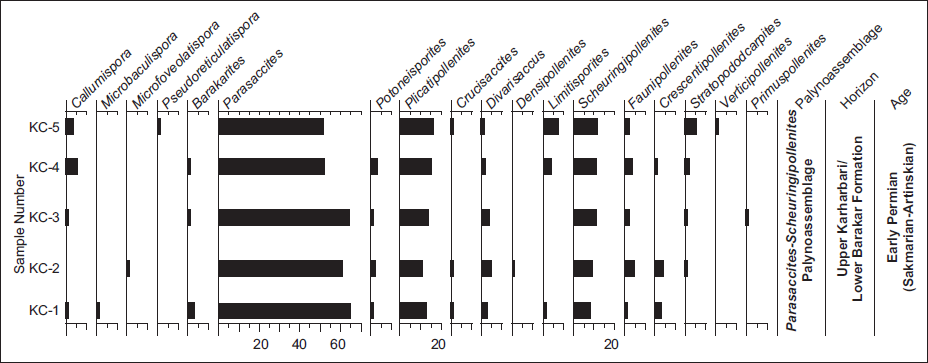
All five samples yielded abundant pollen and spores along with organic matter. The recovered palynoassemblage is diverse and exhibits good preservation of both spores and pollens. Pollens are abundant than spores, together comprising 18 genera and 33 species, along with organic matter. The recovered palynocomposition is dominated by monosaccate pollen grains, that is, Parasaccites (52%–65%) (P. korbaensis, P. obscurus, P. ovatus, Parasaccites sp.), and subdominant taxa include Plicatipollenites (11%–16%) (P. indicus, P. gondwanensis, P. triangonalis, Plicatipollenites sp.) followed by occurrences of Potonieisporites (0%–3%) (P. magnus, P. barrelis), Crucisaccites (0%–1%) (C. latisulcatus), Divarisaccus (1%–5%) (D. lelei), Barakarites (0%–3%) (B. indicus), Densipollenites (0%–1%) (Densipollenites sp.) non-striate bisaccate pollen grains Scheuringipollenites (8%–12%) (S. barakarensis, S. maximus), striate bisaccate pollen grains represented by Faunipollenites (=Protohaploxypinus) (1%–5%) (F. singrauliensis, F. varius), Limitisporites (0%–7%) (L. diversus, L. elongatus), Striatopodocarpites (0%–5%) (S. magnificus, Striatopodocarpites sp.), Verticipollenites (0%–1%) (V. oblongus), Crescentipollenites (0%–4%) (C. fuscus, Crescentipollenites sp.), Primuspollenites (0%–1%) (P. levis) and good number of trilete spores are represented by Callumispora (0%–5%) (C. barakarensis, C. adensa, C. magnifica, Callumispora sp.), Microbaculispora (0%–1%) (Microbaculispora sp.), Pseudoreticulatispora (0%–1%) (P. barakarensis) and Microfoveolatispora (0%–1%) (Microfoveolatispora sp.). The quantitative distribution of recovered palynomorphs is shown in Figure 5. The distribution of palynomorphs shows that the recovered palynoassemblage has dominance of Parasaccites, followed by Plicatipollenites and Scheuringipollenites, along with other significant marker palynotaxa, Crucisaccites and Callumispora. This palynoassemblage is well correlated with the Parasaccites-Scheuringipollenites palynoassemblage of the Upper Karharbari Formation of the Godavari Basin (Jha & Aggarwal, 2011). Moreover, the occurrence of striate bisaccates palynomorphs in the assemblage indicates that this may be the Upper Karharbari-Lower Barakar transition. However, in the Chirimiri Coalfield, the Karharbari Formation has not been lithologically demarcated by any researcher so far. Therefore, this coal-bearing sequence of Kurasia Colliery, Chirimiri Coalfield, is envisaged to belong to the Lower Barakar Formation. Based on the identified palynoassemblage, the age of the studied section is assigned as Early Permian (Artinskian). Stratigraphically significant pollen and spore taxa are illustrated in Plate 2.
Frequency distribution of different pollen and spores recovered from an Early Permian coal-bearing sequence of Kurasia Colliery, Chirimiri Coalfield, Chhattisgarh, Son Basin.
Photomicrographs of Palynomorphs recovered from an Early Permian coal-bearing sequence of Kurasia Colliery, Chirimiri Coalfield, Chhattisgarh. 1). Callumispora barakarensis, BSIP Museum Slide No. 17657, O30/3; 2). Callumispora adensa, BSIP Museum Slide No. 17654, J46/2; 3). Callumispora magnifica, BSIP Museum Slide No. 17654, T55/2; 4). Pseudoreticulatispora barakarensis, BSIP Museum Slide No. 17659, E20/1; 5). Microfoveolatispora sp., BSIP Museum Slide No. 17650, P22/2; 6). Microbaculispora sp., BSIP Museum Slide No. 17645, V36/4; 7). Barakarites indicus, BSIP Museum Slide No. 17655, J27/1; 8). Parasaccites korbaensis, BSIP Museum Slide No. 17645, N12/3; 9). Parasaccites obscurus, BSIP Museum Slide No. 17651, S19/1; 10). Parasaccites ovatus, BSIP Museum Slide No. 17645, V33/3; 11). Plicatipollenites triangonalis, BSIP Museum Slide No. 17650, P39/1; 12). Plicatipollenites gondwanensis, BSIP Museum Slide No. 17646, N14/2; 13). Potonieisporites magnus, BSIP Museum Slide No. 17650, J24/4; 14). Plicatipollenites indicus, BSIP Museum Slide No. 17647, Q30/3; 15). Densipollenites sp., BSIP Museum Slide No. 17650, N15/1; 16). Crucisaccites latisulcatus, BSIP Museum Slide No. 17657, N46; 17). Potonieisporites barrelis, BSIP Museum Slide No. 17651, T43/4; 18). Divarisaccus lelei, BSIP Museum Slide No. 17657, W44/4; 19). Limitisporites diversus, BSIP Museum Slide No. 17658, Q34/4; 20). Scheuringipollenites maximus, BSIP Museum Slide No. 17649, E52/3; 21). Scheuringipollenites barakarensis, BSIP Museum Slide No. 17650, O41/3; 22). Primuspollenites levis, BSIP Museum Slide No. 17653, R25/2; 23). Limitisporites elongatus, BSIP Museum Slide No. 17645, J33/1; 24). Faunipollenites varius, BSIP Museum Slide No. 17649, W33; 25). Striatopodocarpites magnificus., BSIP Museum Slide No. 17655, M49/3; 26). Crescentipollenites fuscus, BSIP Museum Slide No. 17656, L25/2; 27). Verticipollenites oblongus, BSIP Museum Slide No. 17657, Q44/3; 28). Faunipollenites singrauliensis, BSIP Museum Slide No. 17654, F19.

The fossil affinities of the dominant palynotaxa in the present assemblage suggest a strong representation of the Cordaitales, as indicated by genera Parasaccites, Plicatipollenites, and Barakarites. Scheuringipollenites, Faunipollenites, Striatopodocarpites, Limitisporites and Verticipollenites represent Glossopteridales. The taxa such as Potonieisporites, Divarisaccus, Crucisaccites, Densipollenites, Crescentipollenites and Primuspollenites are attributed to Coniferales. In addition, the presence of spores such as Callumispora, Microfoveolatispora, Pseudoreticulatispora and Microbaculispora indicates Filicales (ferns) (Murthy et al., 2024).
Palynofacies analysis
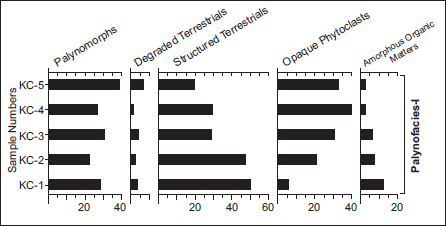
Five samples were investigated for the palynofacies analysis. In this study, organic residues have been widely used to interpret the depositional environments of many sediments, including coal, lignite and associated rocks (Cazzulo-Klepzig et al., 2009; Tyson, 1995). In this study, palynofacies is used to reconstruct the depositional palaeoenvironmental conditions during sediment deposition, following the classification schemes proposed by Tyson (1995) and Batten (1996). The palynofacies is characterised by the dominance of structured organic matters (20%–50%), followed by the sub-dominance of palynomorphs (22%–38%), opaque phytoclasts (6%–33%), amorphous organic matters (2%–12%), and degraded terrestrials (1%–7%). These different organic matter types and their relative frequencies, along with their corresponding sampling depths, are shown in Figure 6 and Plate 3. Based on recovered organic matter, a single palynofacies assemblage has been identified.
Frequency distribution of different organic matter recovered from an Early Permian coal-bearing sequence of Kurasia Colliery, Chirimiri Coalfield, Chhattisgarh, Son Basin.
Photomicrographs of organic matter recovered from an Early Permian coal-bearing sequence of Kurasia Colliery, Chirimiri Coalfield, Chhattisgarh. (SP- Pollen and Spores; DM- Degraded Terrestrials; ST- Structured Terrestrials; OP- Opaque Phytoclasts; AOM- Amorphous Organic Material). 1). BSIP Museum Slide No. 17660, P53/3; 2). BSIP Museum Slide No. 17661, K55/4; 3). BSIP Museum Slide No. 17662, H42/4; 4). BSIP Museum Slide No. 17663, M46/3.

DISCUSSION
A coal-bearing sequence of the Kurasia Colliery, Chirimiri Coalfield, Son Basin, Chhattisgarh State, Central India, has been explored for palaeobotanical and palynological remains. The study aims to assess the biostratigraphic status of the sequence and to understand the palaeovegetation dynamics and depositional environment. The data presented herein is the first record from this colliery of the Son Basin. Fossil floral assemblages are recovered from the carbonaceous shales associated with grey shale, sandstones, and coal-bearing horizons. A rich and diverse megafloral assemblage has been found preserved in these facies and holds a good number of adpressed fossils of Cordaitales, Glossopteridales and Equisetales group of plants. The complete megafloral assemblage comprises eight genera and 21 species. The Cordaitalean genus Noeggerathiopsis dominates the assemblage, followed by Glossopteridalean genera Glossopteris and Gangamopteris, along with the equisetalean genera Paracalamites and Raniganjia. An excellent assemblage of Cordaitalean and Glossopteridales seeds is also recorded. The occurrence of the Noeggerathiopsis and Gangamopteris-dominated floral composition in the assemblage played a significant role in the terrestrial ecosystems of the Talchir and Karharbari formations, whereas the Glossopteris-dominated floral composition characterises the terrestrial ecosystems of the Barakar Formation and the overlying stages of the Lower Gondwana. Therefore, the retrieved macrofloral assemblage indicates a transitional phase between the upper Talchir and overlying Karharbari or lower Barakar times, signifying a gradual change in vegetation from the dominance of forms without midribs to those with definite midribs, characteristic of the overlying formations.
The characterisation of Karharbari deposits as a distinct formational unit has been contentious since the time of their discovery in the late nineteenth century. Although a formal designation of it as a ‘Formation’ was given in the Damodar Basin only, in the other peninsular Gondwana basins, it was identified only based on macrofloral content. It has been argued that the contact between Talchir and Karharbari is erosional and demarcated by a conglomeratic pebble bed (polymictic), derived from the underlying Talchir Formation (Ghosh & Basu, 1967). However, this horizon is not traceable across all the basins, and many oppose it. The lithological characters are generally similar to those of Barakar, but in Karharbari, coal seams are unbanded and not commercially exploitable. In fact, it is the enigma caused by the preference of researchers over the past two centuries, whether based on palaeobotanical, lithological, or depositional evidence. Recently, Das et al. (2025) have discussed in detail its stratigraphic status and history of nomenclature (pp. 17) and opined on its presence in all the Gondwana basins of peninsular India.
It is interesting to note that coal measures of the Kurasia Colliery lithologically belong to the Barakar Formation; and the Karharbari Formation is not lithologically demarcated; however, the complex megaflora recovered from the Colliery shows typical Karharbari floral elements such as Noeggerathiopsis and Gangamopteris in association with Glossopteris, albeit the absence of Buriadia and Botrychiopsis. This Karharbari flora is distinctive from typical Barakar flora and is relatively similar to Talchir flora. In contrast, the lithology of the beds with Karharbari flora is similar to that of the Barakar Formation and distinct from the Talchir Formation. This clearly indicates that Karharbari climatic conditions must have prevailed during the deposition of the lower part of the Barakar Formation and continued into the onward flora. Das et al. (2025) have re-evaluated the status of the Karharbari Formation as a distinct palaeoecological and stratigraphic interval ranging from late Sakmarian to early Artinskian age. They argued this formation as an ecologically transitional unit within the Lower Gondwana.
The presence of five genera, that is, Buriadia, Botrychiopsis, Noeggerathiopsis, Gangamopteris and Glossopteris, forms the basis of the Karharbari Formation. The last three genera can also be found in the Talchir or Barakar Formation. However, Buriadia and Botrychiopsis are restricted only to the Karharbari Formation. In the present assemblage of the Kurasia Colliery, most relevant groups, that is, Cordaitales and Glossopteridales, are represented by both megafossils and palynomorphs. However, the Equisetales are represented only by megafossils, not by palynomorphs. Similarly, the groups Coniferales and Filicales are represented only palynologically, and their most relevant megafossils, that is, Buriadia and Botrychiopsis, are absent in the macroplant assemblage. It is possible that both these genera (Buriadia and Botrychiopsis) perished during sediment transportation due to their weak vegetative morphology. If we combine the above observations, we can say for sure that all five marker plants of Karharbari Formation, that is, Buriadia, Botrychiopsis, Noeggerathiopsis, Gangamopteris and Glossopteris, were growing in the Kurasia Colliery area.
The above-mentioned floral entities, barring Buriadia and Botrychiopsis, have also been reported from the flora of the Lower Barakar Formation (Singh et al., 2006a, 2006b; Srivastava, 1992). It is inferred that the Karharbari flora had continued to exist in the lower Barakar (Artinskian) during Early Permian, but during the upper Barakar times (after Artinskian), most of the earlier forms (midrib-less) disappeared, leading to the dominance of Glossopteris speciation in the assemblage. This evidence clearly suggests that floral similarities between these two geological units do not correspond to lithological similarities, and vice versa. The presence of the above-mentioned plant groups also suggests their importance in the formation of coal in the area.
The recovered megafloral assemblage shows close resemblance to the Karharbari megafloral assemblage described by Maithy (1965) from Giridih Coalfield, Maithy (1969) from Daltonganj Coalfield, Kulkarni (1971) from the South Karanpura Coalfield, Srivastava (1974) from the Auranga Coalfield, Chandra and Srivastava (1982, 1991) from South Rewa Gondwana Basin, and Singh et al. (2006a) from Talcher Coalfield. Furthermore, the recovered megafloral assemblage may also be compared with the megafloral records from the Lower Barakar Formation, such as those described by Maheshwari and Prakash (1965) from Bansloi Valley, Rajmahal Hills. Similar assemblages were also noted by Bajpai (1990) from the Deogarh Coalfield; Srivastava (1992) from the Raniganj Coalfield; Srivastava (1977) and Srivastava and Tewari (1996) from the Auranga Coalfield; Singh et al. (2006b, 2007) from the Ib-River Coalfield. The assemblage also shows close similarity in having Noeggerathiopsis dominant assemblage along with Gangamopteris, as discussed by Gupta et al. (2025) from the adjoining Chirimiri Coalfield.
The recovered floral assemblage also exhibits similarities with those from coeval Early to Middle Permian sequences of the southern Gondwanan continents. These include Antarctica: The Milorgfjella region, Dronning Maud Land (Bose et al., 1990; Plumstead, 1975); Australia: megafloral assemblage from the Irwin Coal Measures Formation within the Perth Basin, Western Australia (Rigby, 1966); South America: Floral stage of Rio Bonito Formation, Paraná Basin from Brazil (Bernardes-de-Oliveira, 1980; Bernardes-de-Oliveira et al., 2016; Iannuzzi & Souza, 2005); South Africa: megafloral assemblage of the lower Ecca Group (Anderson, 1977; MacRae, 1988), Vryheid Formation, Vereeniging in the Karoo Basin (Anderson & Anderson, 1985).
Furthermore, palynological analysis has revealed the dominance of monosaccate pollens, represented by Parasaccites and sub-dominance of Plicatipollenites, followed by occurrences of Potonieisporites, Crucisaccites, Divarisaccus, Barakarites and non-striate bisaccate pollen grain- Scheuringipollenites, striate bisaccate pollen grains Faunipollenites, Limitisporites, Striatopodocarpites, Crescentipollenites, Verticipollenites, Primuspollenites, Densipollenites and trilete spores Callumispora, Microbaculispora, Pseudoreticulatispora and Microfoveolatispora.
In the recovered palynoassemblage, the dominant occurrence of Parasaccites in association with Scheuringipollenites and the presence of Crucisaccites help to corroborate the assemblage with Parasaccites-Scheuringipollenites palynoassemblage of the Upper Karharbari palynoflora of Godavari Basin, as well as other basins in India. The recovered palynoassemblage shows similarity with palynoassemblages described from different coalfields of different Gondwana basins in having common palynotaxa, namely Parasaccites, Plicatipollenites, Potonieisporites, Divarisaccus, Crucisaccites, Barakarites, Densipollenites, Scheuringipollenites, Faunipollenites, Striatopodocarpites, Crescentipollenites, Primuspollenites, Verticipollenites, Callumispora, Pseudoreticulatispora and Microbaculispora. Similar palynoassemblages observed in Damodar Basin are: Palynozone-2 of Raniganj Coalfield (Tiwari, 1973); Son-Mahanadi Basin: Palynozone-2 of Korba Coalfield (Bharadwaj & Srivastava, 1971); Palynozone-2 of Umaria Coalfield (Srivastava & Anand-Prakash, 1984); Assemblage-1 of the Talcher Coalfield (Tripathi & Bhattacharyya, 2001); Satpura Basin: Palynoassemblage-I of Shobhapur and Pathakhera Coal mine from Pathakhera Coalfield (Srivastava & Sarate, 1989). Wardha-Godavari Basin: Biozone-II and III of Umrer Quarry, Nagpur (Bharadwaj & Anand-Prakash, 1974); Palynozone-3 of borecore GGK-20 (Srivastava & Jha, 1989); Upper Karharbari Palynozone of Chintalapudi sub-basin (Jha et al., 2018); Palynozone-1 of Koyagudem area (Aggarwal et al., 2018); Palynoassemblage-A of Umrer Coalfield (Jha et al., 2007); Palynoassemblage-I of Somavaram area (Aggarwal et al., 2019). Moreover, the occurrence of striate bisaccates in the present assemblage indicates that this may be the Upper Karharbari-Lower Barakar transition. This coal-bearing sequence of Kurasia Colliery, Chirimiri Coalfield, is envisaged to belong to the Lower Barakar Formation.
The recovered palynoassemblage has also exhibited strong similarities with palynoassemblages and biozones described from several Gondwanan regions, including: South America: Hamiapollenites karroensis Subzone of the Rio Bonito Formation, Paraná Basin from Brazil (Souza & Marques-Toigo, 2005); Antarctica: Palynoassemblage of the Beacon Supergroup, Dronning Maud Land (Lindström, 1995); Australia: Stage 3 palynoassemblage of the Eastern Province (Kemp et al., 1977); palynoassemblage of Unit IV, Canning Basin (Balme, 1980); and Collie Basin (Backhouse, 1991) in Western Australia; South Africa: Palynoassemblage-1 of the Ketewakai Coalfield (Manum & Tien, 1973) and Palynoassemblage of the Lisimba Member, Idusi Formation (Semkiwa et al., 2003) from Tanzania; Assemblage Zone 2 of the Mid-Zambezi Basin from Zimbabwe (Falcon, 1973); Palynoassemblage-2 of the Luwumbu Coal Formation, North Luangwa Valley (Utting, 1976); palynoassemblage of the main coal seam of the Siankondobo Sandstone Formation (Stephenson, 2008) from Zambia; Palynoassemblage-2 of the lower Ecca Group (Modie & Le Hérissé, 2009); Witbank Coalfield, Karoo Basin (Wheeler & Götz, 2017).
Both mega and miofloral assemblages of the Kurasia Colliery, Chirmiri Coalfield area indicate a dense forest type vegetation dominated by arborescent trees bearing Cordaitales foliage along with other plants belonging to Glossopteridales, Coniferales, Filicales and Equisetales.
Palynofacies analysis reveals that organic debris is predominantly composed of structured phytoclasts (mainly woody elements, tracheids and cuticles) along with palynomorphs. This composition firmly suggests a terrestrial depositional environment. The high percentage of translucent phytoclasts along with pollen grains emphasises a low-energy depositional setting, which is also evident in the lithological framework (Aggarwal et al., 2019; Tyson, 1995). A moderate proportion of opaque phytoclast and low amorphous material in the sediments indicates that their deposition likely occurred in suboxic-dysoxic environments. Therefore, deposition of the studied sediments is envisaged to occur in low-energy, suboxic-dysoxic forest swamp settings.
Depositional Environment
The macrofloral composite reveals the dominance of Cordaitales, followed by Glossopteridales, indicating the presence of a terrestrial marshy ecosystem and prevalence of cooler climatic conditions. These plant groups have been envisaged to flourish in swampy and marshy ecosystems under the influence of cooler, humid climates (Goswami et al., 2006, 2018). This climatic scenario is also evident in the dominant occurrences of these plant groups in the Early Permian sequences and their subsequent reduced occurrences in the late Permian sequences of Indian Gondwana. It is inferred that Cordaitalean-dominated forest was emergent in the proximity of a lake/swamp in the studied area, as also manifested by the prolific occurrences of Noeggerathiopsis in the nearby areas of Chirimiri Coalfield (Gupta et al., 2025). Cordaitales are envisaged to have preferred wetter conditions for growth on broad flood plains of extensive swamps or lake areas. The specimens procured herein are well preserved, suggesting that the deposition site for the vegetal matter was close to the original growing site. It also suggests that the deposition occurred in a lacustrine environment/stagnant water bodies, with vegetal matter derived from trees growing around water bodies. The fine-grained, compact carbonaceous and grey shale with well-preserved plant remains was probably deposited in a low-energy setting of an ox-bow lake, as revealed by the unchanged nature of the biomass. The complete and undamaged leaves of Noeggerathiopsis, Gangamopteris, and Glossopteris clearly indicate a reducing depositional environment, with minimal transportation prior to burial.
The occurrence of large leaves in these sediments supports the existence of conducive warm, temperate and humid climatic conditions during the coal formation (Chandra, 1992; Chandra & Chandra, 1987). The favourable climatic conditions were further substantiated by the recovery of palynoflora, especially Parasaccites-Plicatipollenites-Scheuringipollenites complex, with a higher degree of variation in the diversity of other palynomorphs. The variations in the morphology and abundance of the recovered pollen and spores offer valuable insights into their climatic sensitivity (Tiwari & Tripathi, 1987). The current palynoassemblage is predominantly composed of radial monosaccate pollens (Parasaccites, Plicatipollenites, Potoneisporites, Divarisaccus, Crucisaccites and Densipollenites), typically associated with cool and arid climatic conditions (Stephenson et al., 2007; Tiwari & Tripathi, 1987). Additionally, the assemblage exhibits a notable presence of both non-striate (Scheuringipollenites and Limitisporites) and striate bisaccate pollen grains (Faunipollenites, Crescentipollenites, Striatopodocarpites and Verticipollenites), along with a minor occurrence of pteridophytic trilete spores (Callumispora, Microfoveolatispora, Microbaculispora, Pseudoreticulatispora and Barakarites). This composition suggests a generally cool climate with increased humidity, which is favourable for the development of thin coal seams. Based on floral records, it is envisaged that during the deposition, the climate was moderately cool and humid, which led to the thriving of a mixed forest of Cordaitales together with Glossopterids and Equisetales. The envisaged vegetation scenario is presented in Figure 7.
Vegetational reconstruction of Kurasia Colliery, Chirimiri Coalfield, Son Basin during the Early Permian, based on recovered megafloral assemblage (modified after Pillai et al., 2024). (A) Noeggerathiopsis, (B) Glossopteris, (C) Gangamopteris, (D) Raniganjia, (E) Paracalamites.

Palaeoenvironmental interpretation
All five samples are predominantly composed of structured phytoclasts (mainly woody elements, tracheids and cuticles) along with palynomorphs. This composition clearly indicates deposition in a terrestrial environment. The high percentage of translucent phytoclasts and pollens emphasises a low-energy depositional setting, which is also corroborated by the lithological framework (Aggarwal et al., 2019; Tyson, 1995). Moderate percentages of opaque phytoclasts and low amorphous material suggest that sediment deposition probably occurred in suboxic-dysoxic settings. Therefore, a low-energy, suboxic-dysoxic forest swamp setting is interpreted.
CONCLUSIONS
The present macrofloral and palynological study of a coal-bearing sequence of Kurasia Colliery, Son Basin, has been carried out to infer palaeovegetation dynamics and composition, palaeoclimate and associated depositional environment during the Early Permian. Recovered megafloral and palynological assemblage, along with palynofacies studies, considerably add to our current understanding of the late Karharbari/Lower Barakar flora. Salient findings of the present work are: Retrieved macrofloral assemblage reveals dominance of Cordaitales (Noeggerathiopsis-4 species), followed by Glossopteridales (Glossopteris-7 species, Gangamopteris-5 species), Equisetales (Paracalamites, Raniganjia) and seeds (Cordaicarpus, Samaropsis and Alatocarpus). Noeggerathiopsis-Glossopteris-Gangamopteris macrofloral assemblage represents a transitional phase between the Karharbari (Sakmarian) and the overlying Lower Barakar Formation (Artinskian). Recovered miofloral assemblage has an abundance of monosaccate pollen, mainly Parasaccites, Plicatipollenites and bisaccate pollen, Scheuringipollenites, along with other palynotaxa. This recovered palynocomposition is corroborated by the Parasaccites-Scheuringipollenites palynoassemblage of the Godavari Basin, specifically from the Upper Karharbari Formation. However, the occurrence of striate bisaccates palynomorphs in the assemblage suggests that this may represent the Upper Karharbari-Lower Barakar transition zone, which is envisaged to belong to the Lower Barakar Formation. The recovered palynocomposition shows dominance of Cordaitales over Glossopteridales. Based on macrofloral and miofloral assemblage, the age of the studied sequence is assigned to the Early Permian (Artinskian) age. The dominant occurrence of Cordaitales suggests that this particular plant group was also flourishing on the raised grounds along with glossopterids during the Early Permian. Palynofacies study shows the dominance of structured phytolcasts, suggesting that the studied sediments were deposited in low-energy, suboxic-dysoxic forest swamp settings. The study reveals flourishing of mixed forest comprising Cordaitales, Glossopteridales and Equisetales growing in a terrestrial marshy ecosystem with cooler climatic conditions during the Early Permian.
Footnotes
Acknowledgements
The authors are grateful to the Director, Birbal Sahni Institute of Palaeosciences, Lucknow, for providing the necessary facilities and permission to publish the paper (RDCC/2025-2026/50). Authors are also thankful to anonymous reviewers for their constructive comments and suggestions.
Authors’ Contribution
Anju Saxena: Supervision, conceptualisation, data Interpretation, writing original draft, review and editing. Suyash Gupta: Practical work and analyses, data representation, writing original draft. S. Suresh K Pillai: Palaeovegetation reconstruction and draft review. Srikanta Murthy: Palynological Interpretation, draft review and editing. Husain Shabbar: Data representation and interpretation. Muskan Maurya: Practical work and analyses, data representation. Kamal Jeet Singh: Field sample collection.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Ethical Approval and Informed Consent Statements
This research is in accordance with ethical guidelines that prioritise participant consent, confidentiality, and the integrity of the data.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This study is funded by the Birbal Sahni Institute of Palaeosciences, Lucknow.