
Abstract
The Ediacaran Period was a time when life got big with confirmed fossils of megascopic algae, lichens, sponges, and worms. But which of these kinds of organisms are the most likely biological affinities for enigmatic vendobionts such as Dickinsonia? These iconic quilted organisms have no clear modern relatives, but interpretation as animals has been based in part on putative evidence that they could move of their own accord. These supposed trace fossils of vendobionts, including ‘footprints’ and ‘feeding traces’, are reexamined here for Dickinsonia, Quaestio, and Kimberella. Supposed footprints are more likely tool marks of specimens displaced by wind or water on melting ice, and feeding traces are more likely bundles of ice crystals formed from supercooled water within cavities. Funisia was a sessile creature thought to be able to wave about within the water column, but confinement of the fossils to thin layers is evidence that it was prostrate on the ground. Dickinsonia, Quaestio, Kimberella, and Funisia are found in rocks with sedimentological, petrographic, and geochemical evidence of soil formation, and may have been sessile lichens of Ediacaran terrestrial communities.
Keywords
INTRODUCTION
The Ediacaran Period was a turning point in the evolution of life on Earth, as the time when life got big, meters long rather than millimetres (Payne et al., 2009). Most of the classical Ediacaran fossils from South Australia were originally (Glaessner & Wade, 1966) interpreted as highly mobile animals (Figure 1). Later reassessment objected that they could not have been animals for a variety of reasons, such as asymmetric segmentation (Seilacher, 1992). The discovery of palaeosols beneath the fossils (Retallack, 2012, Seilacher, 1992). The discovery of palaeosols beneath the fossils (2013a) was evidence for an alternative interpretation as some kind of terrestrial vegetation (Figure 2). Only a few Ediacaran fossils were undisputed animals that could move of their own accord, as demonstrated by trace fossils, such as Helminthoidichnites (Plate 1.1) and Archaeonassa (Buatois & Mángano, 2016). These are regarded as burrows and trails of worm-like creatures, perhaps represented by the poorly preserved and tiny (2 × 4 mm) body fossil Ikaria warioota (Evans et al., 2020). Diversity of motile animals in Ediacaran rocks remained low, but a marked increase in trace fossil diversity marks the transition into the Cambrian (Mángano & Buatois, 2017). Non-motile Ediacaran animals such as sponges are also recorded (Wang, Liu, et al., 2024). But what of the other organisms of the Ediacaran age? Could they also move of their own accord? The iconic quilted vendobionts in particular (Plate 1.11–15) have proven problematic for biological affinities, which range from xenophyophore foraminifera (Seilacher et al., 2005), to lichens (Retallack, 1994), cnidarians (Harrington & Moore, 1956), sea pens (Glaessner & Wade, 1966), annelid worms (Glaessner & Wade, 1966), and extinct clades (Pflug, 1973; Seilacher, 1992). Most vendobionts were sessile with holdfasts and even root-like attachments (Mitchell et al., 2019, 2020; Tarhan et al., 2010). This review challenges published evidence that they could move of their own accord (Evans et al., 2024; Ivantsov & Malakhovskaya, 2002). Mobility is a key attribute of animals and thus relevant for assessing alternative hypotheses for biological affinities and habitats of vendobionts.
Classical reconstruction of Ediacaran fossil assemblages of the Ediacara Member of the Rawnsley Quartzite of South Australia at the Museum of the Earth, Ithaca, New York. The museum label in 2015 described this as a ‘legacy diorama’, showing outdated ideas of highly mobile animals promoted by Glaessner and Wade (1966): l-r, Dickinsonia costata, Arborea arborea, Medusinites asteroides, Praecambridium sigillum, Parvancorina minchami, Kimberella quadrata and Cyclomedusa davidi. All but Arborea were then considered highly mobile ocean swimmers.

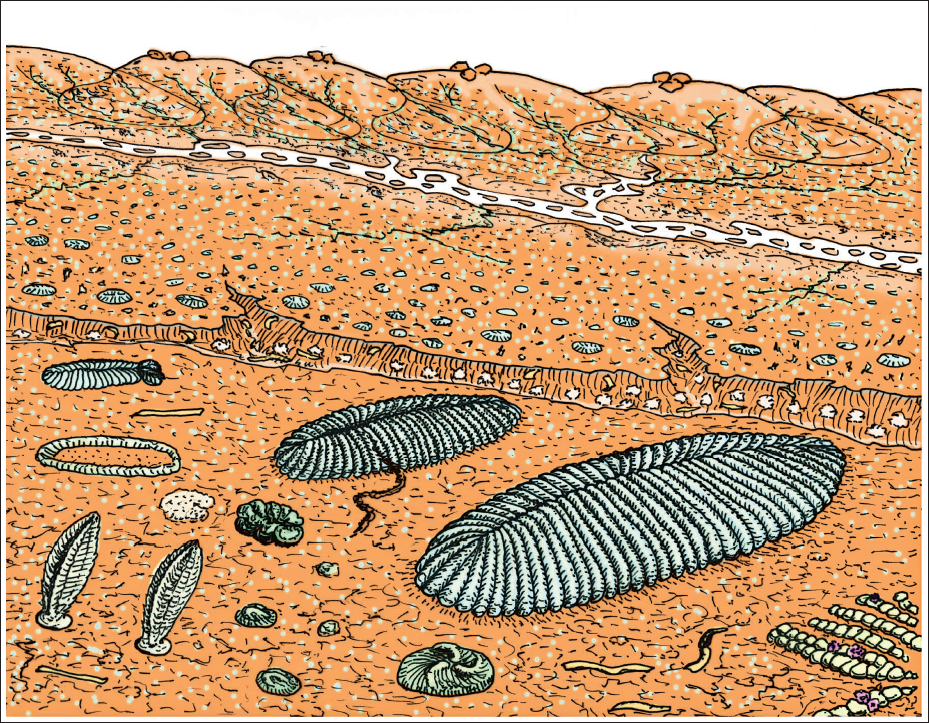
Revised reconstruction of Ediacaran fossil assemblages of the Ediacara Member of the Rawnsley Quartzite based on occurrence in Gypsid paleosols in South Australia, showing l-r, Spriggina floundersi, fragments of Aulozoon soliorum, Phyllozoon hanseni, Dickinsonia costata, Quaestio simpsonorum displaced from former location by wind, Parvancorina minchami, Obamus coronatus, trail of Helminthoidichnites tenuis feeding on Dickinsonia costata, Tribrachidium heraldicum, and Funisia dorothea.
ALTERNATIVE BIOLOGICAL INTERPRETATIONS OF EDIACARAN FOSSILS
While the biological affinities and mobility of Ediacaran vendobionts remain controversial, a variety of biological affinities are plausible because they are represented by other Ediacaran fossils of undisputed taxonomic position. Not all Ediacaran burrows and trails are necessarily from animals because flaring trails (Meyer et al., 2014), such as Lamonte trevallis (Plate 1.3), have been shown to be the work of slime mould (Amoebozoa, Dictyostelida) slugs (grex) formed by aggregation of migrating amoebae (Retallack, 2013b). Other microbial groups also had megascopic expression in the form of stromatolites (Tungussia julia: Walter et al., 1979), lake balls (Nemiana simplex: Retallack, 2016a), and microbial colonies (Cyclomedusa davidi: Grazhdankin & Gerdes, 2007). These sessile microbial communities include fossils previously interpreted as sea jellies, such as Medusinites and Cylomedusa of Glaessner and Wade (1966). There are Ediacaran fossils of a variety of marine algae, including brown, red, and green algae (Plate 1.4; Xiao, Zhang, et al., 1998; Xiao, Knoll, et al., 1998; Xiao et al., 2002, 2020). Fungi of glomeromycotan or mucoromycotinan affinities (Retallack, 2015) are evident from spores with hyphae (Plate 1.9; Grey, 2005) and spores that have fractured in the characteristic brittle manner of fungal chitin (Plate 1.10). Lichenized fungi are best represented by permineralized remains (Plate 1.6) showing thallus differentiation (Plate 1.7) and haustorial connection between hyphae and spherical photobionts (Plate 1.8) of Ganarake scalaris (Retallack, 2022a). Other impressions, such as Nilpenia rossi (Plate 1.5; Droser et al., 2014), have also been considered Ediacaran lichens (Lücking & Spribille, 2024). There are also tubular fossils skeletonised in chitin or calcite, such as Corumbella (Plate 1.2; Osés et al., 2022), Cloudina, and Namacalathus that may represent marine phoronid lophophorates (Shore et al., 2021), or worms (Babcock et al., 2005; Schiffbauer et al., 2020). Trace fossils such as Helminthoidichnites (Plate 1.1) and Archaeonassa may be the work of worm-like creatures (Buatois & Mángano, 2016), such as Ikaria (Evans et al., 2020).
ALTERNATIVE PALAEOENVIRONMENTAL INTERPRETATIONS OF EDIACARAN FOSSILS
On first discovery, Ediacaran megafossils were identified as shallow marine invertebrates (Glaessner & Wade, 1966). Even though the biological affinities of these fossils remain highly problematic (Seilacher, 1992), marine habitats are still urged without corroborating evidence (Runnegar, 2022; Weyland & Droser, 2025). Within the past decade, the distinction between marine and non-marine rocks of Ediacaran age has been tested by various techniques. It began with recognition of palaeosols within red beds with Ediacaran fossils (Retallack, 2012, 2013a), some of them with well-developed calcareous nodules (calcic or Bk horizons) and desert rose pseudomorphs (gypsic or by horizons). The profiles also showed weathering of feldspar to clay, geochemical tracers of that reaction (molar ratio of alumina over bases), and oxidation on land (molar ratio of ferric to ferrous iron). Ediacaran palaeosols are more deeply weathered in nutrient bases and phosphorus than Mesoproterozoic palaeosols, and comparable with early Palaeozoic palaeosols in calculated rate of weathering of nutrient bases (Ca, Mg, Na, and K) and of phosphorus (Retallack, 2022b). Ediacaran isotopic depletion of carbon and oxygen in pedogenic versus marine carbonate is also greater than in Mesoproterozoic palaeosols, as additional evidence of enhanced biogenic weathering on land (Retallack & Bindeman, 2024). Other geochemical discriminators of marine versus non-marine conditions successfully applied to Ediacaran rocks are C/S ratios (Retallack, 2014), Ge/Si ratios (Retallack, 2017), B/K ratios (Retallack, 2020), and light rare earth to heavy rare earth ratios (Retallack, 2024a). Other indications of non-marine conditions are drab-haloed filament traces (Retallack, 2012, 2013a) and the microbial fabric widely identified as ‘old elephant skin’ (Rivularites repertus: Retallack, 2024b). Eolian interbeds with adhesion warts are also evidence of exposure of South Australian Ediacaran sediment to wind (McMahon et al., 2020; Retallack, 2019).
Marine beds of Ediacaran age are also known, and are grey coloured with reduced (ferrous) iron like Phanerozoic marine rocks, rather than red with oxidised (ferric) iron (Retallack, 2014). Ediacaran marine beds are proven by the same molecular ratios and enrichments of nutrient bases and phosphorus mentioned above (Retallack, 2014, 2020, 2024a). Marine Ediacaran rocks contain stromatolites (Walter et al., 1979) and the tubular fossils attributed to animals such as Corumbella and Cloudina (Babcock et al., 2005; Schiffbauer et al., 2020; Shore et al., 2021). The Ediacaran Period is no different from all post-Archean periods in having grey marine or lacustrine, versus red nonmarine and palaeosol facies (Retallack, 2022c). Also grey are Ediacaran intertidal sediments with diagnostic flaser (clay crescent fill) and linsel (lenticular sand) beds (Retallack, 2012, 2013a). Like modern soils of intertidal trees (mangal) and salt marsh, Ediacaran intertidal beds have highly tapering and sinuous paleochannels, abundant pyrite nodules, and distinctive fossil assemblages (Retallack, 2016b, 2016c).
Ediacaran palaeosols have been especially useful for evaluating Ediacaran paleoclimate, which, with one exception, was temperate to periglacial rather than tropical. The exception is the Baltic paleosol, a deeply weathered tropical soil (Oxisol) on flood basalt at the unconformable base of Vendian shales dated to 560 Ma around the Baltic and White Seas, once considered anomalous for its high latitude (Driese et al., 2018; Liivamägi et al., 2014). However, the paleolatitude of Baltica has been reassessed at 557 Ma as 17.6° ± 5.7° (Domeier et al., 2023), so the Baltic palaeosol is no longer a climatic anomaly. Sand wedges, ice wedges, and freeze-thaw banding have been documented from both Cryogenian (Retallack, 2023; Retallack et al., 2015) and Ediacaran palaeosols (Retallack, 2022d). Ediacaran periglacial palaeosols are associated with a series of glacial advances evident from tillites and solifluction flows, labelled Gaskiers (580 Ma), Fauquier (672 Ma), Bou Azzer (565 Ma), and Hankalchough (550 Ma; Retallack, 2022d). These glacial advances are associated with glacioeustatic sea level drawdowns creating paleovalleys as deep as 600 m (Giles et al., 2024; Retallack et al., 2014). Between these stratigraphic levels of glacial facies and periglacial palaeosols are many other palaeosols with weathering ratios and salinisation as evidence of temperate paleoclimatic conditions. The global distribution of Ediacaran palaeosols included gypsic and calcic palaeosols of temperate aridlands (Retallack, 2012, 2013a, 2022e), as well as deeply leached palaeosols of temperate humid paleoclimate (Retallack, 2014, 2016c). The transition from Cryogenian Snowball Earth to the Ediacaran was not an abrupt paleoclimatic change from frigid to tropical, because the Ediacaran Period remained cool with glacial advances (Retallack, 2022d).
MOBILITY OF DICKINSONIA?
Movement of Dickinsonia has been inferred from chains or circles of sometimes overlapping convex impressions in the overlying slab (Plate 2.1–2) interpreted as ‘footprints’ next to concave impressions of a body Evans, Gehling, et al., 2019). These ‘footprints’ have been given a trace fossil name ‘Epibaion’ (Ivantsov & Malakhovskaya, 2002), but that genus is not considered valid and should not be italicised (Retallack, 2021). In other examples, narrow tubes of Aulozoon soliorum (Plate 2.3–5) protruding from below Dickinsonia have been proposed as its trail (Ivantsov et al., 2019). Aulozoon, however, is only a tenth the width of Dickinsonia, and a shallow, hollow structure in thin section (Plate 2.5) below Dickinsonia’s lower surfaces. Dickinsonia is preserved on bed soles, so that Aulozoon superimposed on Dickinsonia (Plate 2.3) means that it was within sediment and below it in life (Retallack, 2007, 2022e; Seilacher et al., 2005; Surprenant et al., 2023). Interpreting Aulozoon as a streamer up into overlying water (Surprenant et al., 2024) is thus impossible.
The mechanism of movement of Dickinsonia has been proposed as muscular expansion and contraction (Gehling et al., 2005) or ciliary crawling (Ivantsov, 2011). Slight concentric contraction of Dickinsonia has also been explained as tentacles (Harrington & Moore, 1956) or hypothallus (Retallack, 2007), and is not the kind of peristalsis needed for forward motion. Ciliary crawling may work for millimetric creatures such as placozoans (Srivastava et al., 2008), but is unlikely for Dickinsonia as long as 1.5 m (Retallack, 2007).
Evidence against the movement of Dickinsonia includes indentation of adjacent specimens (Plate 1.12) similar to competitive standoff of sedentary creatures, such as encrusting corals (Jackson & Buss, 1975), bryozoans (Taylor, 2016), or lichens (Brodo et al., 2010). In thin section, the lower surface of Dickinsonia has rhizinelike projections and attached Aulozoon reaching down into the substrate, unlike the smooth upper surface (Plate 2.4). Other specimens of Dickinsonia are split or folded by cracking and folding of their substrate (Evans, Huang, et al., 2019). Especially telling is lack of reaction or avoidance of a burrowing worm (Plate 2.1). This may be a case of herbivory, rather than scavenging as argued by Gehling and Droser (2018), because Dickinsonia retained high relief and definition, unlike known and measured decayed specimens, which are measurably deflated (Retallack, 2007). Furthermore, that Helminthoidichnites trail (Plate 2.1) has sharp levees, like those of subaerial, rather than subaqueous, trails (Wang, Davies, et al., 2024).
Alternative hypotheses for Dickinsonia movement are as algal wracks or ‘snow mice’. McIlroy et al. (2009) considered ‘Epibaion’ comparable with impressions left in beach sands by wracks of dead algae heaped on beaches by storms. Beach shorefaces have parallel lamination, unlike the sedimentology and geochemistry of Dickinsonia beds (Retallack, 2013a, 2019, 2020, 2024b). ‘Snow mice’ are polsters of still-living moss or lichens driven by wind on melting ground ice common in periglacial alpine and polar regions (Hotaling et al., 2020; Pérez, 1997). This explanation satisfies observed directions or curve of movement (Evans, Gehling, et al., 2019) better than imagined animal intention, which, in some cases, was aimless. Comparable modern vagrant polsters are the moss Grimmia (Plate 2.6) and lichens Xanthoparmelia (Plate 2.7) and Dimelaena (Plate 2.8). The growth of ground ice severs both rhizinous and cord attachments to the substrate, and strong katabatic winds or their eddies from uplands displace them short distances. There is independent evidence of permafrost from periglacial involutions and ground ice at stratigraphic levels with Dickinsonia in South Australia and Russia (Retallack, 2021). Dickinsonia could not move of its own accord.
Dickinsonia (Plate 1.1, 1.12) has long been regarded as a marine worm (Glaessner & Wade, 1966), but fundamental problems preclude that interpretation. It cannot have been an annelid because the segments alternate along a midline, instead of true metameric segmentation (Seilacher, 1992). This distinctive arrangement is similar to the geologically older vendobiont Fractofusus, which has a fractal pattern of branching tubes (Taylor et al., 2023). Damaged specimens show adventitious regrowth by a mechanism of a terminal meristem, a mode of growth found in plants and lichens, but not in worms or arthropods (Retallack, 2022f). Also, unlike soft-bodied marine worms, Dickinsonia resisted compaction by overlying sedimentary rock to an extent requiring stiff biopolymers, such as chitin, unknown in worms (Retallack, 1994).
Crustose lichen is an alternative interpretation of Dickinsonia as a sessile organism with compaction-resistant chitin, chambered internal structure, fractal organisation, filamentous histology, terminal meristem, and basal rhizines and cord. A lichen interpretation is also compatible with non-marine occurrence atop palaeosols (Retallack, 2012, 2013a), within eolian interflag sandstone laminae (Retallack, 2019), and on matrices with low B/K ratios (Retallack, 2020), high Ge/Si ratios (Retallack, 2017), and high light rare earth to heavy rare earth ratios (Retallack, 2024a).
MOBILITY OF QUAESTIO?
Quaestio simpsonorum is another enigmatic Ediacaran fossil with imprints as convex epireliefs on the sole of the cover bed beside concave epireliefs of the fossil itself (Plate 3.1). These have been interpreted as evidence that it moved of its own accord, as if the imprints were a ‘footprint’ of animal motion (Evans et al., 2024). Like ‘Epibaion’ already discussed for Dickinsonia (Plate 2.1), these are not trace fossils of deliberate animal activity (Retallack, 2021), but tool marks from the lateral physical displacement of individuals (Plate 2.2). Such imprints cannot be used as evidence that Quaestio was an animal, especially considering that it has no other animal-like characteristics. Quaestio has an internal curve that lacks bilateral or radial symmetry, sometimes taken as hallmarks of animals (Seilacher, 1992). Evans et al. (2024) are convincing in their argument that similarities between Quaestio and living animals such as Placozoa or Acoela are superficial at best, not the least in the greater relief of Quaestio compared with deeply buried, soft-bodied creatures (Retallack, 1994). Many specimens of Quaestio are irregularly lobed, more like a thallus than a body. Such lobes enable rolling in tumbleweed lichens, which can be dispersed by wind (Plate 3.3). The unfolding spiral growth and irregular crossing ridges of Quaestio are superficially similar with growth and thallus lobes of modern jelly (Collema) and rock tripe (Umbilicaria) lichens (Plate 3.4–5). Other Ediacaran fossils like modern jelly lichens are Obamus coronatus (Dzaugis et al., 2020), similar to the living jelly lichen Anema nummularium (Boissiere & Montavont, 2006), and Attenborites janeae (Droser et al., 2020), similar to Collema nigrescens (Otálora et al., 2010).
The occurrence of Quaestio on surfaces with ‘old-elephant skin’ (Plate 2.1) is evidence for subaerial exposure, because of hackly appearance, microtuffets, and both open and healed cracks characteristic of paleobotanical form taxon Rivularites repertus, unlike undulose subaqueous microbial textures (Retallack, 2024b). Quaestio is also associated with interflag sandstone laminae of eolian origin (Retallack, 2019).
On land, lateral displacement may have been caused by wind gusts of frozen individuals on melting ice (Retallack, 2021). Such ‘snow mice’ of displaced polsters of mosses or vagrant lichens are common in periglacial regions today (Hotaling et al., 2020; Pérez, 1997). Other evidence of periglacial conditions in the same beds as Quaestio include needle ice, periglacial convolutions, low chemical index of alteration, and geochemical low paleotemperature proxies (Retallack 2016b, 2021). Furthermore, the laterally correlative Billy Springs Formation of South Australia includes tillites and dropped pebbles (Jenkins, 2011), as part of the widespread Hankalchough glaciation (Wang et al., 2023).
MOBILITY OF KIMBERELLA?
Kimberella is a distinctive horseshoe-shaped impression of segments, enclosing an ovoid hollow, often with unusually high relief (Plate 4.1–2), though less concave examples also are known (Plate 4.4). Its segments have been compared with muscle attachments of limpets and monoplacophorans, but are not subtle, rounded, shallow shell indentations like muscle scars of molluscs (Fedonkin & Waggoner, 1997). The deep, quadrangular, hollow channels of Kimberella are most like the marginal segments of Ernietta (Retallack, 2016a), and deflated Kimberella with a terminal flange (Plate 4.4) is very similar to Ernietta eroded from a normal life position that was erect within sediment (Ivantsov et al., 2016). Not all Ernietta have strong lateral pleating; some are now known to have been smooth on the outside (Smith et al., 2017), like Kimberella. Kimberella is also an order of magnitude larger than the oldest known molluscs, and separated in age from them by 20 million years (Runnegar, 1981). Some Kimberella have been taken as evidence of trails (Ivantsov et al., 2019), but have high relief and walled, lateral segments along their length, suggesting that they are unusually elongate individuals, like elongate Ernietta (Smith et al., 2017).
Kimberella commonly has elongate structures radiating from the open end (Plate 4.1–2, which have been interpreted as feeding scratches made by a long proboscis (Gehling et al., 2014; Ivantsov et al., 2019). The feeding scratches are also found without ‘Kimberichnus’ (Plate 4.3), which has been considered to have fed and moved on, though not leaving a trace of their motion on freshly ploughed ground. The animal did not retreat backwards, because there are no trails exiting radial arrangements of the scratches (Plate 4.1–2, 4.3). These so-called ‘feeding scratches’ are much longer and more parallel than known molluscan feeding traces (Radulichnus: Retallack, 2021). Furthermore, they do not have the form of deep scratches with laterally overturned sediment, and some interpenetrate into the ground below and the slab above (Plate 4.3–4). Kimberichnus has yet to be found with an extended proboscis, and the elongate ones have the open end for ‘feeding scratches’ at one end (Ivantsov et al., 2019), here interpreted as needle ice. Thus, they are pseudofossils rather than trace fossils and should not be italicised (‘Kimberichnus teruzzi’: Retallack, 2021)
These structures represent a variety of ground ice. Bundles of these ice crystals are like frost flowers (Plate 4.5) with sharp, faceted terminations like monoclinic ice crystals with helical twinning (Plate 4.3–4). They are unusually sharp and parallel for radular scratches, and commonly displace small (1–2 mm) clods of sediment (Plate 4.1–2). The needles radiate from a segment of a terminal flange (Plate 4.4), like that of Ernietta (Ivantsov et al., 2016). Modern frost flowers form from supercooled reservoirs of water in hollows of plants (Mason et al., 1963). Frost flowers on high-latitude sea ice are distinct in having lower width-to-length ratios (Roscoe et al., 2011). Also found on these same surfaces are vertical palisades that raise large divots of substrate (Plate 4.1–2), like common vertical ground ice, known as pipkrake (Plate 4.6; Lawler, 1988, 1993). Thus, neither Kimberella nor its hypothetical proboscis were mobile.
MOBILITY OF FUNISIA?
Funisia dorothea is an enigmatic structure of serial spherical structures in monospecific assemblages covering large areas of many Ediacaran slabs (Plates 5.1–2), and interpreted as benthic marine sponges or cnidarians (Droser & Gehling, 2008). Like these invertebrates, it was envisaged as attached to the seafloor by a spherical, high-relief holdfast, but with the chains of spheres vertical and moving freely in water like seaweeds. The spherical elements were hollow and are preserved as both internal and external moulds (Surprenant et al., 2020).
Chief among the problems for this interpretation is the way in which Funisia forms pavements of mainly parallel branches, with limited overlap, and no strong divergence of branches, very different from fossil algae in the same beds (Xiao et al., 2020), or from algae on modern beaches (McIlroy et al., 2009). No Funisia strands have been found bending up into the overlying slab, which preserves them, including individual Funisia strands which curve or bend laterally (Surprenant et al., 2020). Furthermore, supposed ‘holdfasts’ of Funisia are small, and long chains are not folded as if tangled when fallen to the substrate. Similar observations of Ediacaran fronds such as Charnia (Plate 1.13) and Trepassia with small holdfasts and unfolded fronds have been taken as evidence that they were instead sediment recliners like Dickinsonia (Plate 1.1, 12) and Fractofusus (McIlroy et al., 2021; Narbonne & Gehling, 2003). Furthermore, Funisia resembles Dickinsonia (Retallack, 2007) in showing a gradation from high relief, well-preserved fossils to decayed and effaced surfaces with impersistent traces of laterally shared walls (Surprenant et al., 2020). Another problem is the occurrence of Funisia with the microbially induced fabric Rivularites repertus (Plate 5.1–2), widely known as ‘old elephant skin’. Rivularites have the cracked, buckled, and radial growth textures of what is also called biological soil crust or microbial earth, rather than undulose, domed, or folded fabrics of marine or lacustrine subaqueous microbial mats (Retallack, 2024b). True aquatic algae in these same beds are never found on surfaces with Rivularites (Xiao et al., 2020). Funisia is also found in interflag sandstone laminae with adhesion warts and granulometry indicating eolian deposition (McMahon et al., 2020; Retallack, 2019). A final problem is the high relief of Funisia, more like vendiobionts than probable algae in the same beds (Xiao et al., 2020).
An alternative view of Funisia dorothea is as a crustose to foliose lichen cover of the ground, like the living Namibian desert lichen, Stellarangia (Plate 5.3–5). This interpretation solves a problem for the misinterpretation of some nodes of Funisia as high relief, narrow holdfasts (Surprenant et al., 2020), because they are along rather than at the base of Funisia chains. Holdfasts of other Ediacaran fronds are much wider, sometimes with anchors of radiating rhizines (Tarhan et al., 2010). These less compacted parts of Funisia may instead be dense reproductive features such as sclerotia, soredia, or apothecia, similar to living Stellarangia elegantissima (Plate 5.5). Fresh versus decayed, and bent versus overlapping strands of Funisia (Surprenant et al., 2020) are also matched by this comparison (Plate 5.3). This is a novel terrestrial view of Funisia, very different from the vision of them as underwater meadows (Droser & Gehling, 2008).
CONCLUSIONS
Putative evidence that the iconic quilted Ediacaran fossils known as vendobionts could move is reexamined and found wanting. Supposed ‘footprints’ of Dickinsonia and Quaestio are not indications that they could move of their own accord, but rather tool marks from temporary stations of the specimens moved by wind or water. One mechanism suggested by paleoclimatic data from associated palaeosols and palaeoenvironmental implications of eolian interbeds is that Dickinsonia and Quaestio were like ‘snow mice’, polsters of lichen driven by wind on melting ice over periglacial soils (Retallack, 2021). Supposed feeding traces of Kimberella, show no resemblance to molluscan radular scratches, but instead are casts of needle ice like those of temperate and periglacial soils. Finally, Funisia is not preserved in tangled wracks or with diverging branches like seaweeds which were attached to the bottom but waved about in a water column. Funisia is found with microbial earth textures and eolian bed forms, and formed prostrate ground cover. Dickinsonia, Quaestio, Kimberella, and Funisia are unlike any known animals, and may have been terrestrial lichens.
Terrestrial red beds, unlike intertidal to marine grey beds, are now widely recognized in Ediacaran rocks using a variety of petrographic (Retallack, 2012, 2013a), sedimentological (McMahon et al., 2020), paleopedological (Driese et al., 2018), and geochemical evidence (Retallack, 2020, 2024a). These studies highlight a difference between marine grey beds with stromatolites and tubular fossils such as Cloudina and Corumbella of ‘Ediacaran Wormworld’ (Schiffbauer et al., 2016), compared with terrestrial red beds with vendobionts of Ediacaran ‘Mattressland’ (Retallack, 2022f). Classical quilted Ediacaran fossils (Vendobionta) were not motile animals but sessile, like vegetation, and dwarfed tiny trail-making animals (Figure 2). Preservation styles of Ediacaran fossils are inexplicable if they were animals (Seilacher, 1992), but can be matched by Phanerozoic preservation styles of fossil plants (Retallack, 2022c). Also, more like plants and lichens are the complex fractal growth patterns of Ediacaran ‘fronds’ (Brasier et al., 2012). Nearest neighbour analysis of vendobionts in red beds confirms the conclusions of this article, that they were sessile and had limited interaction (Mitchell et al., 2019, 2020). Also, more like terrestrial vegetation than marine communities, is the complex rank abundance distribution of Ediacaran assemblages (Darroch et al., 2015). Red bed vendobiont communities also had much higher beta diversity than modern and Phanerozoic benthic marine communities (Finnegan et al., 2019). Trace fossils and animals in non-marine red beds are small (Buatois & Mángano, 2016; Evans et al., 2020), as predicted from theoretical considerations of animals capable of terrestrial invasion (Boyce, 2023), but there is a greater variety and size of trace fossils in latest Ediacaran marine to intertidal deposits (Mángano & Buatois, 2017). Increased biomass and complexity of Ediacaran life on land by an order of magnitude compared to earlier in Earth history is reflected in a variety of geochemical proxies of weathering through geological time (Retallack, 2022b; Retallack & Bindeman, 2024).
A variety of Ediacaran megascopic life: algae (4), slime mould (3), worm tube (2), worm burrow (1), lichens (6–10), and enigmatic vendobionts (11–15): (1) Helminthoidichnites tenuis burrow cutting across Dickinsonia costata from Nilpena Member of Rawnsley Quartzite, Nilpena, South Australia; (2) Corumbella werneri from Tamengo Formation in Itaú quarry, Ladário, Brazil; (3) Lamonte trevallis from Shibantan Member of Denying Formation near Wuhe, China; (4), Longifuniculum sp. cf. dissolutum from Nilpena Member of Rawnsley Quartzite, Nilpena, South Australia; (5) Nilpenia rossi from Nilpena Member of Rawnelsy Quartzite Nilpena, South Australia; (6–8) Ganarake scalaris, hand specimens dark in matrix and details of thallus structure, spherical photobionts, tubular hyphae, and skeletal bipyramidal crystals of pruina whewhellite, from the Noonday Formation of Galena Canyon, Death Valley, California; (9), Ceratosphaeridium mirabile, Wilari Dolomite Member, Tanana Formation, Observatory Hill no., 1 well, northern South Australia; (10) Schizofusa zangwenlongii, Dey Dey Mudstone, Observatory Hill bore, northern South Australia; (11) Rangea scheiderhoehni from Kliphoek Member of Dabis Formation on farm Aar, near Aus, Namibia; (12) Dickinsonia costata from Ediacara Member of Rawnsley Quartzite Ediacara Hills, South Australia; (13) Charnia masoni from Bradgate Formation in Pocketgate quarry, England; (14) Pteridinium simplex from Aarhauser Sandstone on Aar farm, southern Namibia; (15) Ernietta plateauensis from Kliphoek Member of the Dabis Formation in Farm Aar, Aus, Namibia. Specimens are (1) South Australian Museum P35740 from J.G Gehling, (2) Ohio State University Geology Museum 46402 from Gabriel de Matos, (3) Virginia Polytechnic Institute Geoscience Museum 4670 from M. Meyer, (4) South Australian Museum P48767 from S. Xiao, (5) Nilpena RO8-155 from M.Droser, (6–8) Museum of Natural and Cultural History, University of Oregon F116110A ; (9–10) Geoscience Australia Canberra Paleontological Collection slides 36417 and 36355 from K. Grey, (11) Geological Survey of Namibia Museum, Windhoek F182, (12) South Australian Museum F137460, Retallack photo, (13), Leicester Museum LEIUG 2328, Retallack photo; (14) Retallack field photo; (15) Geological Survey of Namibia Museum Windhoek F1244, Retallack photo.

Dickinsonia costata, ‘Epibaion costatum’, Aulozoon soliorum, and comparable modern vagrant mosses and lichens: (1–2) Body fossil Dickinsonia costata, with presumed resting traces ‘Epibaion costatum’ from Ediacara Member of Rawnsley Quartzite in Ediacara Hils, South Australia; (3) detail of Aulozoon soliorum tubes (arrow) connected to Dickinsonia costata deep within slab, and below other specimens of Phyllozoon hanseni from Nilpena Member of Rawnsley Quartzite, Bathtub Gorge, South Australia; (4), thin section of Dickinsonia ornata below ripple-marked sandston from Ediacara Member of Rawnsley Quartzite in Brachina Gorge, South Australia; (5), Thin section of Aulozoon soliorum below basal impression of Dickinsonia costata from Nilpena member of Rawnsley Quartzite near Hookapunna Well, South Australia; (6) Grimmia pulvinata on alpine soil near Salzburg, Austria; (7) Xanthoparmelia reptans on red soil between red mallee (Eucalyptus socialis) near Damara station, inland New South Wales; (8) Dimelaena oreina on Steens Basalt at Paisley Caves, Oregon. Fossil specimen sources are (1–2) South Australian Museum P14359, Retallack photo; (3) South Australian Museum P35665–86; (4–5) Museum of Natural and Cultural History University of Oregon specimens F115735 and F117937; (6) Wikimedia Darkone; (7–8) Retallack photos.

Quaestio simpsonorum and comparable modern vagrant and jelly lichens: (1) body fossil (right arrow) and trace impression (left arrow) of Quaestio simpsonorum from the Nilpena Member of the Rawnsley Quartzite, Nilpena, South Australia; (2) reconstruction of Quaestio simpsonorum and its trace on ‘old elephant skin’ (Rivularites repertus) microbial earth on a Wadni pedotype; (3) tumbleweed shield lichen Xanthoparmelia vagans from Cortez, Colorado; (4) jelly lichen Collema tenax near Worland, Wyoming; (5) rock tripe lichen Umbilicaria torrefacta, Wells Gray Park, British Columbia Canada. Fossil specimen (1–2) NENP LV-FUN 591s 1287e (Evans et al., 2024); (3) photo Dale Ward; (4) photo Tyrell Perry; (5) photo Jason Hollinger.

Kimberella quadrata, ‘Kimberichnus teruzzi’, and comparable modern ice needles and ground ice: (1–2) specimen and annotated sketch of Kimberella quadrata, and supposed radular scratches given invalid name ‘Kimberichnus teruzzi’, here interpreted as ice needles, from Zimnie Gory Formation at Zimnie Gory, White Sea, Russia; (3) frost flower (‘Kimberichnus teruzzi’ from Zimnie Gory Formation at Solza River near Archangelsk Russia; (4) Kimberella quadrata and ‘Kimberichnus teruzzi’ from Ediacara Member of Rawnsley Quartzite in Brachina Gorge, South Australia; (5) frost flower growing from water-filled hollow in plant stalk from roadside pond near Frisco, Colorado (2 May, 2011); (6) Frost palisades uplifting soil clods southwest of Granges–sur–Vologne, France (28 December, 2008). Fossil specimen (1–2) in Paleontological Institute Moscow 3493/5137, photo Dmitry Grazhdhankin; (3) Paleontological Institute Moscow 3993/5619, photo Dmitry Grazhdhankin; (4) South Australian Museum P35657, photo Retallack; (5) photo Robert Berwyn; (6) photo Thomas Bresson.

Funisia dorothea, and comparable living lichens Stellarangia namibiensis and S. elegantissima: (1–2) Funisia dorothea from the Nilpena Member of the Rawnsley Formation at Nilpena, South Australia; (3–4) Stellarangia namibiensis from 15 km east of Swakopmuind, Namibia; (5) Stellarangia elegantissima with prominent apothecia, from 20 km northeast of Swakopmund, Namibia. Fossils (1–2) South Australian Museum P40275 and P40309; lichen photos (3–5) by Ingeborg van Leeuwen.

Footnotes
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author disclosed receipt of the following financial support for the research, authorship and/or publication of this article: Sandal Society of the Museum of Natural and Cultural History of the University of Oregon.