
Abstract
The Himalayan orogeny played a pivotal role in shaping South Asia’s climatic and ecological evolution, leading to the deposition of approximately 6000 m of molasse sediments in the Himalayan Foreland Basin, known as the Siwalik Group. While fossil flora of the Siwalik sediments in western and eastern Himalaya has provided valuable insights into past vegetation shifts, the plant fossil record from eastern Nepal remains poorly documented. In this study, we describe fossil fern of Aspleniaceae and a leaf of Lagerstroemia (Lythraceae) from the Lower Siwalik sediments of the Kankai Mai River section, eastern Nepal. Aspleniaceae, a family of tropical and subtropical ferns and Lagerstroemia, a genus characteristic of tropical moist and dry deciduous forests, suggest a warm and humid climate with seasonal precipitation during the deposition period. The fossils and previously reported records from the region suggest that tropical evergreen to deciduous forest covered eastern Nepal during the Lower Siwalik period. The genus Lagerstroemia likely originated in India and diversified during the Paleogene, while the Aspleniaceae highlight potential phytogeographic connections between South Asia and Southeast Asia. These findings provide insights into possible migration routes of these taxa following the collision of the Indian and Eurasian plates.
INTRODUCTION
The deformation and uplift of the Himalayan belt led to the accumulation of approximately 6000 m of molasse sediments in the Himalayan Foreland Basin, extending ~2400 km from Pakistan in the west to Arunachal Pradesh in the east. The sediments in this basin, known as the Siwalik Group, lie between the Main Boundary Thrust (MBT) to the north and the Himalayan Frontal Thrust (HFT) to the south (Gansser, 1964; Valdiya, 2002). The Siwalik sediments, deposited from the Middle Miocene to the Lower Pleistocene, are classified into Lower, Middle and Upper Siwalik (Parkash et al., 1980; Pilgrim 1910, 1913). These units exhibit a coarsening-upward fluvial succession, while individual beds display a fining-upward pattern (Nakayama & Ulak, 1999; Rai & Yoshida, 2021).
The Siwalik flora is essential for understanding the impacts of Himalayan orogeny on climate, vegetation shifts, monsoon patterns and elevation changes in the Himalayan‒Tibetan region (Ding et al., 2017; Srivastava et al., 2018). A transition from evergreen to deciduous forests has been documented in the Lower and Middle Siwalik sediments of both eastern and western Himalaya (Adhikari et al., 2024c; Bhatia et al., 2021, 2022; Srivastava et al., 2018), likely driven by rising temperatures and declining precipitation (Bhatia et al., 2021, 2022; Hoorn et al., 2000; Srivastava et al., 2018; Wu et al., 2014). However, the floral diversity of eastern Nepal remains poorly documented, with existing fossil records being limited (Adhikari et al., 2018, 2022, 2023, 2024a, b; Srivastava et al., 2017). This makes the region crucial for understanding the evolutionary history of its plant communities.
In this study, we describe two new fossil impressions from the Lower Siwalik sediments of the Kankai Mai River section in eastern Nepal and discuss their phytogeographical and climatic significance.
GEOLOGY OF THE STUDY AREA
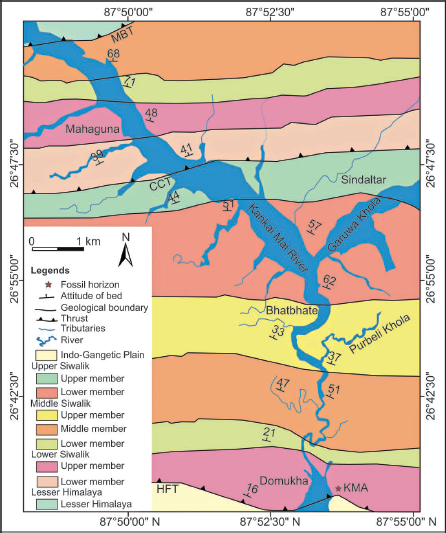
The study area is located within the Siwalik Group of the Kankai Mai River section in eastern Nepal (26.68° N, 87.9° E) (Figures 1 and 2). The Siwalik Group in this region is divided into southern and northern belts, separated by the Central Churia Thrust (CCT) (Ulak, 2009, 2016). Lithostratigraphically, it comprises three litho units—Lower Siwalik, Middle Siwalik and Upper Siwalik—differentiated based on lithological variations and the thickness of sandstone and mudstone (Ulak, 2009, 2016).
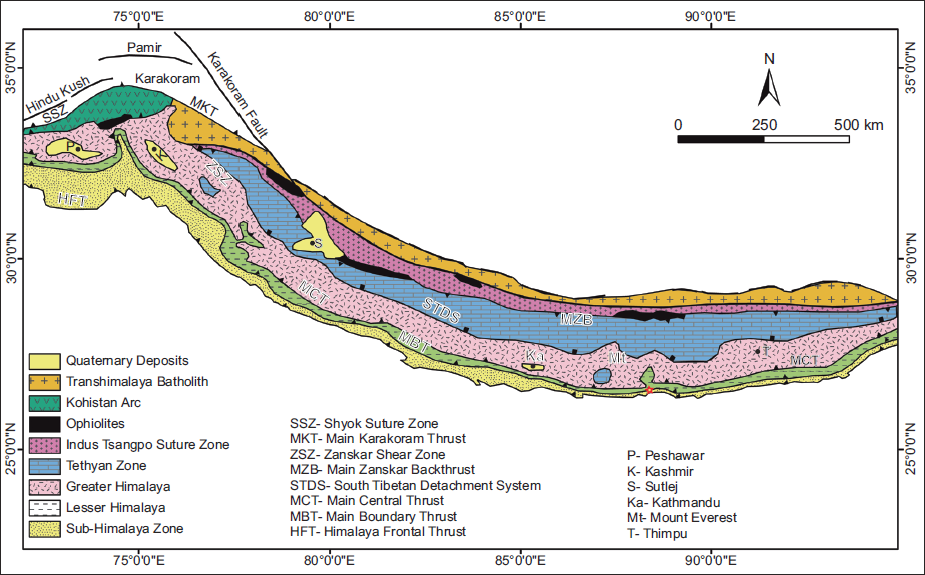
The Lower and Upper Siwalik are further divided into lower and upper members, whereas the Middle Siwalik comprises lower, middle and upper members. The Lower Siwalik is characterised by fine- to coarse-grained sandstone interbedded with variegated to dark grey mudstone and dark to light grey siltstone. The Middle Siwalik comprises medium- to very coarse-grained ‘salt and pepper’ sandstone to pebbly sandstone, dark grey to black mudstone and light grey siltstone. Less indurated sandstone beds are predominantly found in the middle part of this unit. The Upper Siwalik consists of poorly to well-sorted cobble-pebble-boulder conglomerates, reddish-brown sandstone, dark grey siltstone and mudstone. These Siwalik successions were deposited in a fluvial system (Ulak, 2009, 2016).
The Lower Siwalik and the lower member of the Middle Siwalik were formed under a meandering river system. In contrast, a braided river system deposited the middle to upper members of the Middle Siwalik and the Upper Siwalik (Ulak, 2009, 2016). The fossil leaf impressions were excavated from grey siltstone beds within the upper member of the Lower Siwalik (Figure 3).
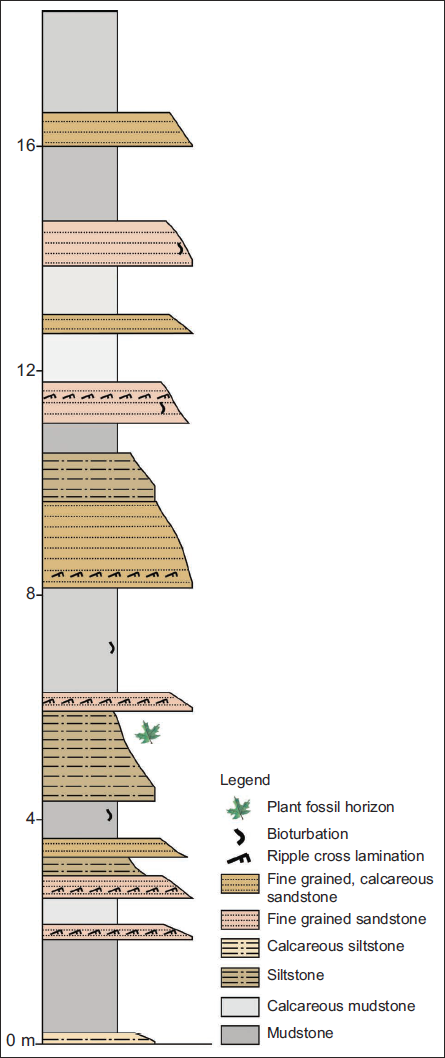
MATERIALS AND METHODS
A soft brush, hammer and fine chisel were used to carefully expose the fossil leaf impressions, which were then photographed under low-angle natural sunlight using a digital camera (Canon G7X Mark II). Minute features of the leaf impressions were examined using a stereoscopic microscope (Leica S8APO). The descriptive terminology follows the leaf architecture classification of Dilcher (1974) and Ellis et al. (2009). Fossil identification was carried out at the herbarium of the Forest Research Institute (FRI), Dehradun and the Central National Herbarium (CNH), Howrah, India. All fossil impressions are housed in the Department of Geology, Birendra Multiple Campus, Bharatpur, Chitwan, Nepal.
SYSTEMATICS
Thelypteridaceophyllum tertiarum Joshi and Mehrotra (2003) (Plate 1, Figures 1–3)
Description: Frond sterile, nanophyll to microphyll in size, chartaceous, margin entire, narrow to wide oblong in shape with medial symmetry, preserved length and width varying 1.4–4.3 cm and 0.8–2.2 cm respectively; apex and base broken; venation goniopteroid, mid vein moderate to stout in thickness; 6–11 pairs of fused pinnules visible, maximum preserved length and width of pinnules 8.2 mm and 3.1 mm respectively, size decreasing upwards, oblong to narrow oblong in shape with a thin midrib; relation to frond mid vein (costa) narrow acute to obtuse (35°–103°); 7−11 pairs of lateral veins in pinnules visible, angle of origin of lateral veins narrow to moderate acute (29–52°), simple, opposite, straight and fine.
Number of specimens: Three.
Figured specimen numbers: KMA46a and KMA46b (part and counterpart), KMA53.
Locality: Left bank of Kankai Mai River, Domukha.
Horizon: Lower Siwalik.
Age: Middle to Late Miocene.
Affinities: The diagnostic characteristics of the fossil frond, such as the costa, fused pinnules and the lateral venation pattern, suggest an affinity with the family Aspleniaceae (synonym Thelypteridaceae) (Beddome, 1970; Central National Herbarium (CNH) sheet ID: CNH 52291, Plate 1, Figure 1).
Holttum et al. (1970) analysed morphologically similar genera, including Ampelopteris Kunze, Cyclosorus Link, Goniopteris Presl and Thelypteris Schmidel. These genera are distinguished by traits such as unicellular glandular or non-glandular hairs, spores, rhizome anatomy and chromosome numbers. However, as the present fossil lacks these diagnostic characteristics, it cannot be identified at the generic level.
Joshi and Mehrotra (2003) established the organ genus Thelypteridaceophyllum for similar fossil fronds. Based on its morphological resemblance to the previously described Thelypteridaceophyllum tertiarum Joshi and Mehrotra (2003), the present fossil is assigned to the same species.
Lagerstroemia himalayaensis Srivastava, Gaur et Mehrotra (2015) (Plate 2, Figure 1)
Description: Leaf simple, complete, microphyll in size, chartaceous; lamina symmetrical, shape wide elliptic with medial symmetry; preserved lamina length and width 4.4 cm and 2.2 cm respectively, estimated length to width ratio 1.9; lamina unlobed, entire margin; apex seemingly convex, angle acute (71⁰); base convex, obtuse angle (104⁰); primary vein moderate to stout in thickness, straight; secondary veins 9 pairs visible, predominantly alternate with excurrent attachment to the primary vein, eucamptodromous venation, arching upwards to join super-adjacent secondary veins, distance between two secondary veins variable, ranging 1.5–8.7 cm, angle of divergence moderate−wide acute angle (48−70⁰); intersecondary veins present, covering <50% length from the margin, distal course of intersecondary veins parallel to secondary veins, terminating into percurrent tertiary veins; intercoastal tertiary veins percurrent, wavy, convex, alternate to sometimes opposite with RR, AO, OR angle of origin; further fine details poorly preserved.
Number of specimens: One.
Figured specimen numbers: KMA17.
Locality: Left bank of Kankai Mai River, Domukha.
Horizon: Lower Siwalik.
Age: Middle to Late Miocene.
Affinities: The fossil leaf exhibits characteristic features such as a symmetrical lamina with a wide elliptic shape, eucamptodromous venation with a moderate to wide acute angle of divergence and the presence of intersecondary veins, placing it taxonomically close to the genus Lagerstroemia L. of the family Lythraceae. Numerous herbarium sheets from families such as Anacardiaceae, Annonaceae, Combretaceae, Moraceae and Myrtaceae were consulted and compared with the nearest living relatives of the fossil, housed at the Central National Herbarium (CNH) in Howrah and the Forest Research Institute (FRI) in Dehradun, India, along with data from online databases like GBIF and POWO. Detailed comparisons with taxa such as Anacardium occidentale (L.) Gaertn, Parishia maingayi Hook f. and Toxicodendron acuminatum (DC.) Ming of the Anacardiaceae, Alphonsea lutea (Roxb.) Hook. f. & Thomson, Oxymitra fornicata (Roxb.) Hook. f. & Thomson of the Annonaceae, Combretum decandrum Jacq., Terminalia chebula Retz. of the Combretaceae, and Artocarpus integrifolia L. f. and Ficus pumila L. of the Moraceae revealed differences in venation patterns, apex and base morphology, intersecondary vein occurrence, and tertiary vein patterns compared to the fossil. Similarly, Pimenta dioica (L.) Merr. of the Myrtaceae, which exhibits brochidodromous venation, and Psidium guineense Sw. (Myrtaceae), with moderately acute secondary veins, also differed from the fossil.
Additionally, comparisons were made with various taxa within the Lythraceae family, such as Ammannia baccifera L., Lythrum salicaria L., Woodfordia fruticosa (L.) Kurz, Lagerstroemia calyculata Kurz, L. floribunda Jack, L. hypoleuca Kurz, L. indica Linn., L. parviflora Roxb., L. speciosa (L.) Pers., L. tomentosa Presl, L. turbinata Koehne and L. villosa Wall. ex Kurz, Lawsonia alba Lam. and Rotala indica (Willd.) Koehne. Characteristics such as brochidodromous venation observed in Ammannia baccifera, Lythrum salicaria and Woodfordia fruticosa leaves, nanophyll size lamina in Lawsonia alba and Rotala indica, attenuate to mucronate apex and rounded base in Lagerstroemia calyculata, L. indica Linn., L. tomentosa and L. villosa, simple to forked percurrent tertiary veins in L. hypoleuca, reticulate tertiary veins in L. turbinata and mesophyll-sized leaves in L. floribunda, were significant points of differentiation from the present fossil. The fossil leaf exhibited the closest similarity to Lagerstroemia parviflora Roxb. (FRI Herbarium sheet no. 21817; Plate 2, Figure 2).
Fossil leaves of Lagerstroemia have been reported extensively across different parts of the Indian subcontinent. The genus Lagerstroemia has been documented in Siwalik sediments in Nepal (Dwivedi et al., 2006; Prasad, 1994, 2013; Prasad et al., 2019), India (Alok et al., 2023; Antal & Awasthi, 1993; Khan et al., 2019; Lakhanpal & Dayal, 1966; Prasad et al., 2004, 2017 Srivastava et al., 2015), Oligocene sediments in Japan (Tanai & Uemura, 1991), Eocene sediments in India (Lakhanpal & Guleria, 1981; Singh et al., 2011, 2015, 2019), Tertiary (Singh & Prasad, 2009, 2010) and Miocene sediments in India (Agarwal, 2002). A comprehensive comparison of the present fossil with previously reported Lagerstroemia species indicates the closest resemblance to Lagerstroemia himalayaensis Srivastava et al. (2015), thus assigning the present fossil to the same species.
DISCUSSION
The Aspleniaceae, the largest family of ferns, comprises 24 genera and approximately 950 species, predominantly distributed across tropical and subtropical regions worldwide (POWO, 2024; Smith et al., 2006). Notably, species diversity is highest in low-latitude Malesia (~440 species), followed by the Neotropics (~300 species) and Africa and its neighboring islands (55 species) (Smith, 1990). Most family members thrive in damp, primary undisturbed forests, typically at low to mid-elevations within or near rainforest edges (Smith, 1990).
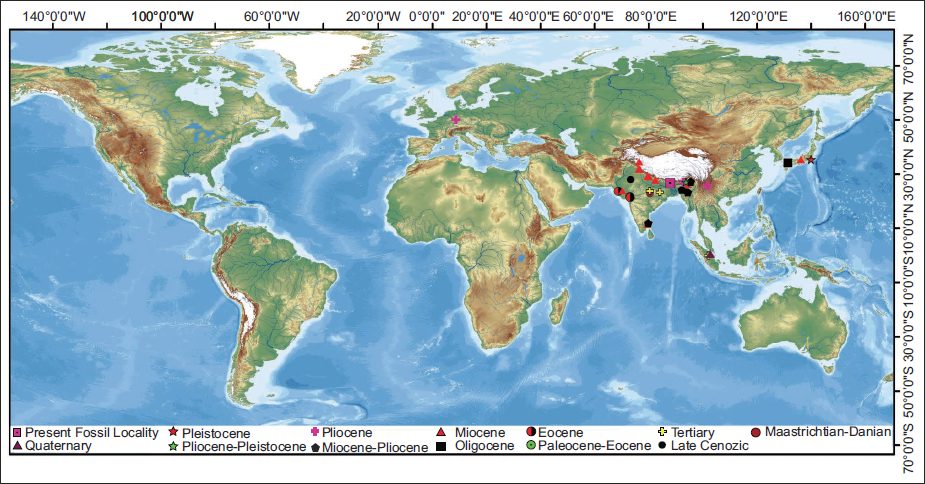
Megafossils of the Aspleniaceae family are well-documented from the Cretaceous and continue throughout the Cenozoic (Figure 4; Supplementary Table 1). The earliest known fossil of this family has been reported from the late Maastrichtian–early Paleocene sediments of central Colorado, USA (Berry, 2019). During the Paleogene, Aspleniaceae across Africa, China, Europe, North America, South America and Southeast Asia (Figure 4). In the Neogene, their fossil records indicate occurrences in Europe, New Zealand, South America, East Asia and South Asia (Figure 4), with evidence of their presence in South Asia during the Miocene. Fossilised remains of an Aspleniaceae genus from the Paleocene sediments of Indonesia (Kräusel, 1929) suggest that the family might have migrated to South Asia from Southeast Asia, possibly after the collision of the Eurasian and Indian plates (Chatterjee et al., 2013); however, other possible routes cannot be ruled out.
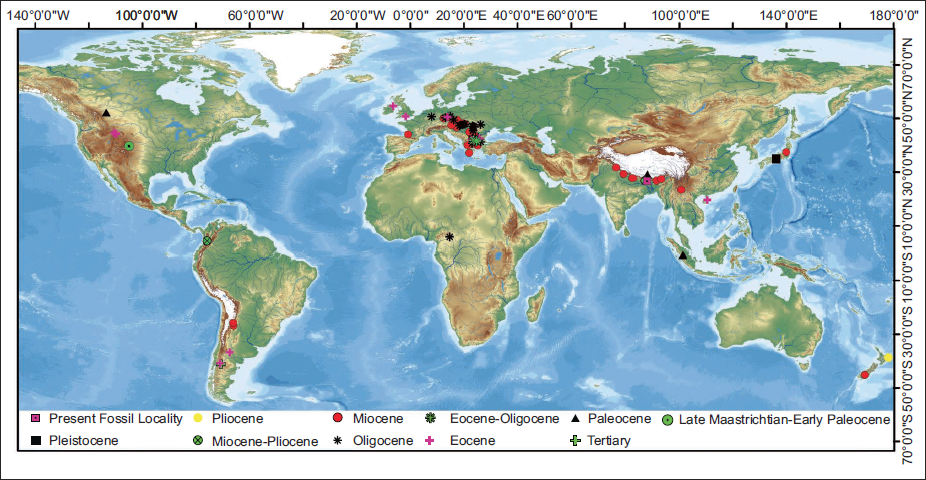
The genus Lagerstroemia, comprising approximately 53 species, includes deciduous and evergreen shrubs and trees distributed across tropical regions of South Asia, Southeast Asia, northern Australia and parts of Oceania (Mabberley, 1997). Lagerstroemia parviflora is predominantly found in tropical moist and dry deciduous forests of India, Myanmar and Nepal (Champion & Seth, 1968; Press et al., 2000). Megafossil records of Lagerstroemia, including leaves, woods and fruits, are well documented from the Indian subcontinent, dating back to the Cretaceous and continuing throughout the Cenozoic (Figure 5; Supplementary Table 2). Beyond the Indian subcontinent, fossil representatives of the genus have also been reported from Europe, East Asia, Southeast Asia and Oceania during the Cenozoic (Figure 5). The earliest known fossil record of Lagerstroemia, from the late Maastrichtian–early Danian sediments of the Deccan Intertrappean beds of India (Shukla et al., 2021), suggests a Gondwanan origin. Furthermore, it also suggests that the genus likely originated in India (Shukla et al., 2021; Srivastava et al., 2015) and subsequently diversified into other regions probably during the Paleogene.
Sterile frond of the fossil and the modern fern. (1) Fossil fronds of Thelypteridaceophyllum tertiarum Joshi & Mehrotra showing shape, size, and venation pattern such as mid vein of the pinna (costa) (red arrows), fused pinnule margin (dark green arrows), midrib of pinnules (yellow arrows), and lateral veins of pinnules (pink arrows). (2) and (3) Fossil pinna showing shape, size, and venation pattern such as mid vein of pinna (red arrow), fused pinnule margin (dark green arrow), midrib of pinnules (yellow arrows), and lateral veins of pinnules (pink arrows). (4) Modern pinna of Cyclosorus proliferous Tard. and Chr. showing shape, size, and venation pattern such as mid vein of pinna (red arrows), fused pinnule margin (dark green arrows), midrib of pinnules (yellow arrows), and lateral veins of pinnules (pink arrows) (Scale bar = 1 cm).

Fossil leaf of Lagerstroemia himalayaensis Srivastava, Gaur et Mehrotra and modern leaf of Lagerstroemia parviflora Roxb. (1) Fossil leaf of Lagerstroemia himalayaensis showing shape, size and venation pattern such as primary vein (white arrows), secondary vein (yellow arrows), base (light blue arrow) and apex (red arrow). (2) Modern leaf of Lagerstroemia parviflora showing shape, size, and venation pattern such as primary vein (white arrows), secondary vein (yellow arrows), base (light blue arrow), and apex (red arrow) (Scale bar = 1 cm).

The current megafossil record of Lagerstroemia (Lythraceae) and an Aspleniaceae frond from the Lower Siwalik sediments of eastern Nepal, along with a previously described Arecaceae leaf fossil (Adhikari et al., 2022), suggests the presence of a tropical evergreen to deciduous forest during the deposition period. Quantitative climate reconstruction of the Lower Siwalik sediments in the Darjeeling area, which is geographically close to the fossil locality, using the Co-existence Analysis (CA) indicates a mean annual temperature of 27.25 ± 0.35°C, with warm and cold month mean temperatures of 28.2 ± 0.1°C and 25.6 ± 0.3°C, respectively (Bhatia et al., 2022). The reconstructed mean annual precipitation was 2269.5 ± 58.5 mm, while precipitation levels during the wettest, driest and warmest months were 267 ± 4 mm, 31 ± 12 mm and 174 ± 47 mm, respectively (Bhatia et al., 2022). Overall, the quantified climatic data (Bhatia et al., 2022) and the plant megafossil records suggest that a warm, humid and seasonal climate prevailed at the fossil locality during the Lower Siwalik period.
CONCLUSIONS
The discovery of Aspleniaceae and Lagerstroemia fossils from the Lower Siwalik sediments of eastern Nepal enhances our understanding of the region’s palaeoflora and climate during the Middle Miocene. These fossil records and previously reported fossils suggest a warm and humid environment with seasonal precipitation, aligning with quantitative climate reconstructions from nearby Siwalik deposits, supporting the presence of tropical evergreen to deciduous forests. Furthermore, the documented fossil taxa indicate possible floristic exchanges between South Asia and Southeast Asia, providing insights into the migration and diversification of these lineages post-Indian-Eurasian collision. The genus Lagerstroemia possibly has an Indian origin, with possible diversification during the Paleogene. Future investigations into additional floral elements from the eastern Siwalik region will be crucial in refining our understanding of the palaeoecology and vegetation dynamics in response to Himalayan tectonics and climate change during the Middle to Late Miocene.
Footnotes
Acknowledgements
PA is grateful to the Head of Department, Central Department of Geology, Tribhuvan University, Kathmandu and Campus Chief, Birendra Multiple Campus, Tribhuvan University, Bharatpur, Chitwan. HB, S and GS are thankful to the Director of the Birbal Sahni Institute of Palaeosciences, Lucknow, for providing necessary facilities and encouragement during the research work. The authors are grateful to the Director of the Central National Herbarium (CNH), Howrah, and the Forest Research Institute (FRI), Dehradun, India, for providing access to herbarium sheets for consultation. We thank Lalit Kumar Rai and Ashok Poudel for their assistance during our fieldwork.
Data Availability Statement
All data used are available in the manuscript and the supplementary files.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest regarding the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: The present research work was supported by the University Grants Commission (UGC), Sanothimi, Bhaktapur, Nepal [Grant Number PhD-76/77-S&T-8] to PA.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.