
Abstract
Pteropod aragonitic shells contribute to 12% of biogenic inorganic carbon flux to the world ocean but are also prone to dissolution. Though their state of preservation and generic ratios, namely Creseis/Limacina have been used as a proxy for palaeoclimatic reconstruction, their taxonomic attributes have remained unexplored. The present study reports 20 pteropod species from a single location in the northeastern Arabian Sea offshore of Saurashtra, all of which show the same degree of preservation throughout the core. The occurrence of mesopelagic fauna in the study area located on the shelf region however points to palaeoenvironmental changes and underscores the importance of employing pteropod assemblages for palaeoclimatic reconstructions in the study area.
Introduction
As pteropods are made up of aragonite (a more soluble form of CaCO3), their shells are susceptible to changes in the water column chemistry as compared to calcitic foraminiferal shells. Thus, the state of preservation of pteropod shells has been used to understand the effect of ocean acidification, as even a slight variation in the seawater pH affects the aragonitic shell of pteropods (Bednaršek et al., 2017). They have mostly been used to understand the variation in the aragonite compensation depth (ACD) over the past centuries (Rottman, 1979; Singh, 2007; Singh et al., 2017). Based on their global distribution, it is estimated that pteropods contribute a minimum of 12% to the biogenic flux in the world oceans (Bednaršek et al., 2012). Despite being one of the main contributors to pelagic aragonite, they have been neglected during the carbonate budget estimation. If the calcification of their inorganic shells serves as a carbon sink through the biological pump, exporting particulate inorganic carbon to the ocean floor, it also releases carbon dioxide as part of the carbonate counter pump (Salter et al., 2014; Manno et al., 2018; Anglada-Ortiz et al., 2021).
Very few quantitative estimates of pteropod flux to the world oceans have been made (Anglada-Ortiz et al., 2021; Bednaršek et al., 2012), which are in turn dependent on the size and weight of taxa, which varies from ocean to ocean. Ambokar et al. (2022) for the first time demonstrated the potential of using pteropod taxonomy in paleoclimatic reconstructions underscoring the importance of the species-based, biological sensitivity of pteropods. Whilst shells of different species had the same state of preservation, temporal variation in climatic conditions was reflected by change in the assemblage. This approach looked beyond the aragonitic shells of pteropods and their preservation state, whilst using it as a palaeoclimatic indicator.
These results from the Saurashtra coast form the basis of the present work, which attempts to document all pteropod taxa from the same region. The offshore of Saurashtra is a very important sedimentary basin holding clues to changes in the evolution of the Indian monsoon and hydrodynamics therein, over the past glacial–interglacial cycles.
Study Area
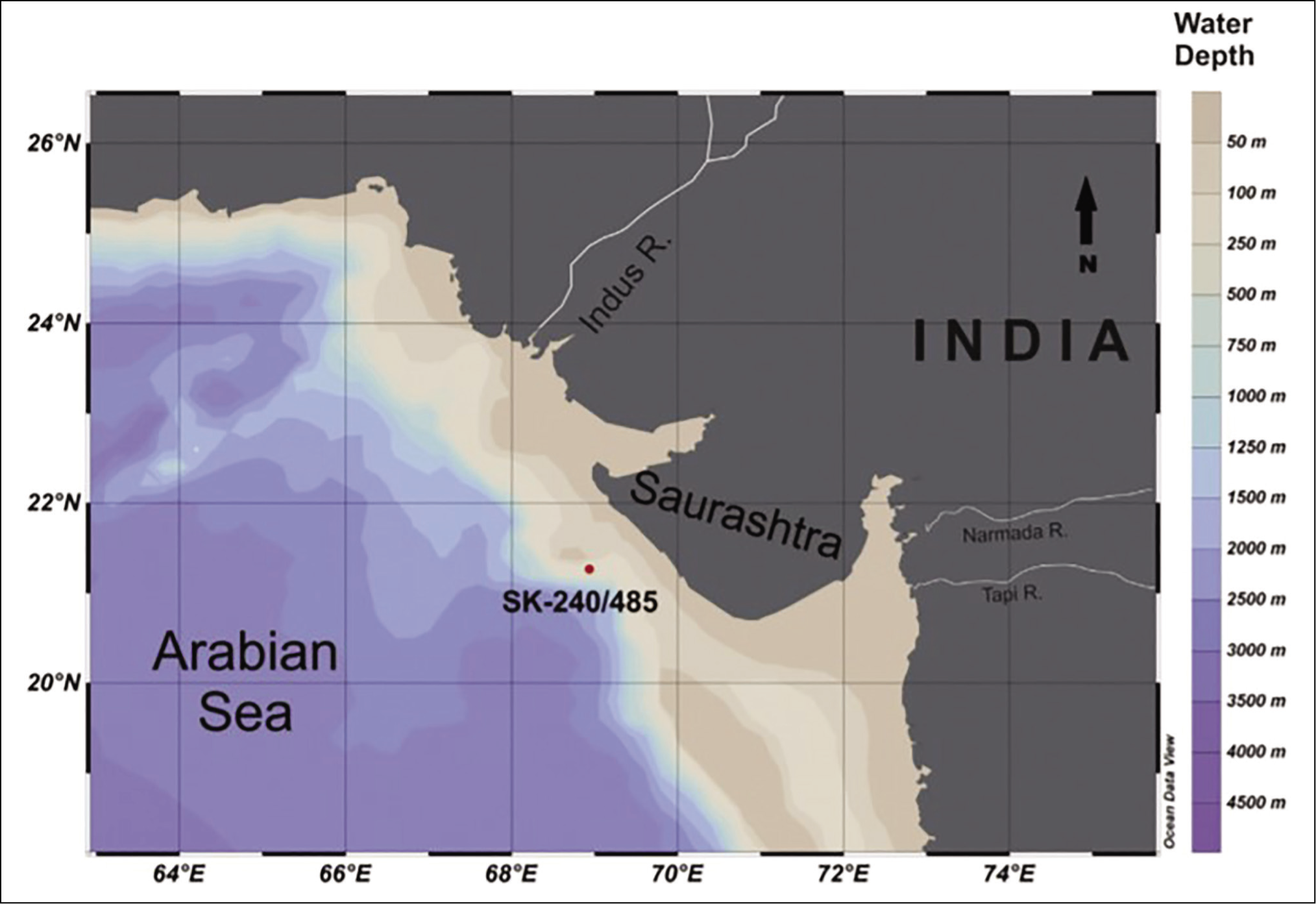
The present study addresses a location on the outer shelf of the western continental shelf of India (Figure 1), which is highly influenced by the freshwater influx received by the Indus, Tapti and Narmada Rivers (Rao & Rao, 1995). Ambokar et al. (2022) suggest the continuous influence of fresh water at the core site through 12,000 to 15,000 years BP. Azharuddin et al. (2017) suggested that the productivity in this region is influenced by the nutrients sourced from the Indus River. This region is also influenced by monsoonal upwelling caused by strong SW monsoon (Wyrtki, 1973).
Map of core locations studied (adopted from Azharuddin et al., 2017).
Methodology
For the present study, pteropods were picked out from the sand fraction of a sediment core SK- 240/485 collected at a water depth of 88 m off the shores of Saurashtra, India. The sand fraction was separated using standard and established procedures described in Panchang et al. (2007), Panchang and Nigam (2014), Ambokar et al. (2022) and several others. The sand fraction obtained was poured on a glass slab and cone-quartered until a representative fraction was obtained for picking pteropods. In total, 300 specimens from 85 samples have been picked for reliable taxonomic observation (Pielou, 1966). The picked pteropods were mounted on faunal slides. The pteropods were sorted based on species and counted. The established taxonomic classification for euthecosomatous pteropods identification by Bé and Gilmer (1977) has been adopted.
Results
In the present study, 20 species of pteropods have been recorded. Of these Limacina inflata (d’Orbigny, 1836) (95%–16%), Limacina trochiformis (d’Orbigny, 1836) (0%–12%), Limacina lesueuri (d’Orbigny, 1836) (0%–16%), Limacina bulimoides (d’Orbigny, 1836) (0%–9%), Creseis acicula (Rang, 1828) (0%–72%), Creseis virgula (Rang, 1828) (0%–21%), C. virgula (Rang, 1828) conica (Eschscholtz, 1829) (0%–12%) occur as major species and C. virgula (Rang, 1828) constricta (Chen & Be, 1964) (0%–4.9%), Peraclis reticulata (d’Orbigny, 1836–1846) (0%–6%), Peraclis depressa (Meisenheimer, 1906) (0%–3.55%), Peraclis rissoides (Tesch, 1903) (0%–1%), Styliola subula (Quoy & Gaimard, 1827) (0%–1.7%), Cuvierina columnella (Rang, 1827) (0%–1.4%), Diacria quadridenta (de Blainville, 1821) (0%–0.34%), Diacria trispinosa (de Blainville, 1821) (0%–0.72%), Clio pyramidata (Linnaeus, 1767) (0%–0.71%), Clio cuspidate (Bosc, 1801), Clio polita (Pelseneer, 1888) (0%–0.86%), Cavolina longirostris (de Blainville, 1821) (0%–0.71%) and Cavolina uncinata (Rang, 1829) (0%–0.24%) occur as minor species (Plate 1 and Plate 2). All the major species represent epipelagic assemblage; the minor species represent both, the epipelagic and mesopelagic assemblage, whilst the bathypelagic realm is represented by a single species C. polita.
1(a, b) Peraclis depressa (Meisenheimer, 1906); 2 Peraclis reticulata (d’Orbigny, 1836–1846); 3 Peraclis rissoides (Tesch, 1903); 4 Creseis virgula (Rang, 1828) conica (Eschscholtz, 1829); 5 Creseis virgula (Rang, 1828) constricta (Chen and Be, 1964); 6 Creseis acicula (Rang, 1828); 7 Creseis virgula (Rang, 1828); 8 Styliola subula (Quoy and Gaimard); 9 Limacina lesueuri (d’Orbigny, 1836); 10 (a, b) Limacina inflata (d’Orbigny, 1836); 11 Limacina bulimoides (d’Orbigny, 1836); 12 Limacina trochiformis (d’Orbigny, 1836); 13 Clio pyramidata (Linnaeus, 1767); 14 Cuvierina columnella (Rang, 1827); Scale Bar = 100 µm.
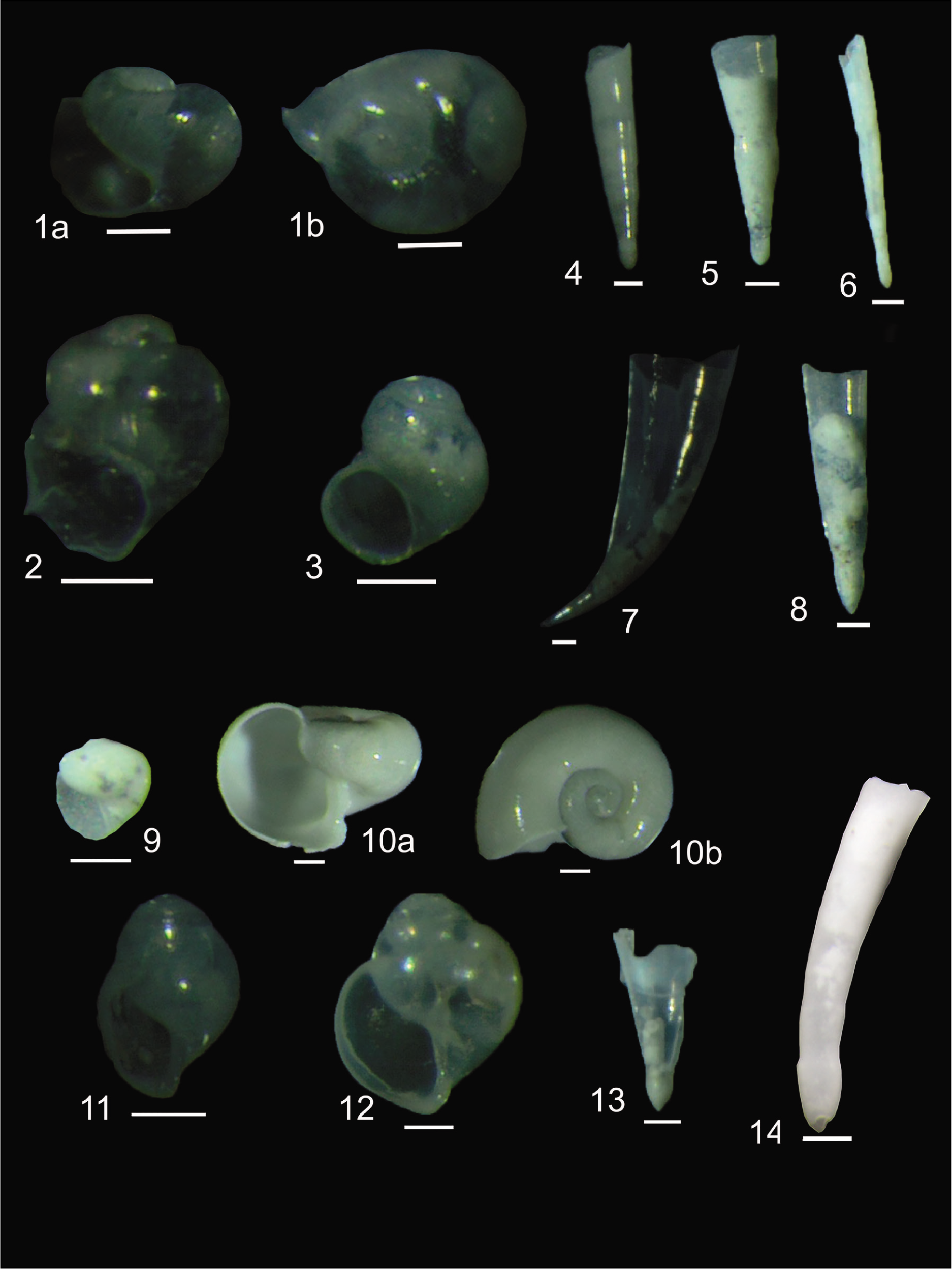
1 (a, b) Cavolina longirostris (de Blainville 1821); 2(a, b) Diacria quadridenta (de Blainville, 1821); Scale Bar = 500 µm.

Discussions
The present study for the first time reports 20 species of pteropods from a single location from off Saurashtra in the northeastern Arabian Sea. Bhattacharjee and Mallik (2000) reported only 4 to 5 pteropod species from 8 locations near the Carlsberg Ridge. Singh et al. (2005) studied the distribution of pteropods along the entire continental shelf of the eastern Arabian sea and reported only 8 pteropod species. Singh (1998) reported 6 pteropod species from the 3 locations off the Kerala coast. Almogi-Labin et al. (1991) also reported only 6 pteropod species from the 2-sediment cores located in the Red Sea. Bhattacharjee (2005) also reported 20 pteropod species from a vast area in the middle Andaman Sea, distributed over 8 different locations. Panchang et al. (2007) reported 10 pteropod species from the one sediment core location in the northern Andaman Sea and 18 species from the entire Ayeyarwady Delta Shelf, an area spread across 2 lakh sq km (Panchang, 2008; Panchang & Nigam, 2023). Hence, this is the highest diversity of pteropods being reported from any one location in the Northern Indian Ocean.
Out of all the species reported in the present study, C. columnella has never been reported before from the Arabian Sea. Sakthivel (1977) reported it from the southern Indian Ocean, whilst Bhattacharjee and Mallik (2000), Bhattacharjee (2005) and Bhattacharjee and Bandyopadhyay (2002) reported C. columnella from the Andaman Sea. It is worth exploring the environmental implications of the same.
Clift et al. (2002) and Azharuddin et al. (2017) have reported the study area to have been under the influence of freshwater from the Indus River. The change in pH of the water column due to freshwater influx could be the cause (Panchang & Ambokar, 2021) of the translucent, white colour of the pteropod shells recovered. However, the state of preservation of shells of all species is the same throughout the core, and thus is not indicative of paleoenvironmental variations in the study area, if any. Ambokar et al. (2022) demonstrated that temporal variation in even minor pteropod taxa effectively recorded past climatic changes. The downcore abundances of the pteropod taxa recovered in the present study should thus hold clues to such palaeoclimatic information given their biological sensitivity. Additionally, this taxonomic database supplemented with shell morphometrics should also help in quantifying the carbonate flux to the ocean in the study area.
Conclusions
This present study reports 20 species of pteropods from offshore Saurashtra, as baseline data for employability in palaeoclimatic studies. C. columnella is being reported for the first time from the Arabian Sea. All the species show the same state of preservation, thus undermining the importance of this attribute as a versatile palaeoclimatic indicator. The presence of mesopelagic species on the outer shelf off Saurashtra is suggestive of palaeo-temperature variations rather than palaeo-bathymetric variations in the study area.
Footnotes
Acknowledgements
The authors would like to thank the Ministry of Earth Sciences (MoES), Government of India for the research funding (Grant No. MoES/CCR/ Paleo – 1/ 2019). M. A. is thankful to MoES for the Senior Research Fellowship. The authors are thankful to the Vice Chancellor and Head, Department of Environmental Science, Savitribai Phule Pune University and Director, Birbal Sahni Institute of Palaeosciences for all infrastructure and logistic support to the ongoing project. We would also like to thank NCPOR, Goa and the Shipboard team of the Sagar Kanya-143 cruise for sampling core location SK-240/485.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors M.A. and R.P. received financial support for research from Ministry of Earth Sciences (MoES), Government of India (Grant No. MoES/CCR/ Paleo – 1/ 2019). P.G. and S.A. received financial support for research from Fastrack grant of SERB-DST under project No. SR/FTP/ES-53/2013.